
左山一葡萄与SO4及101-1杂交F1代部分植株抗旱性鉴定
2017-04-07 03:29:13郭淑华刘笑宏牛彦杰陈天行翟衡杜远鹏
中外葡萄与葡萄酒 2017年2期
郭淑华,刘笑宏,牛彦杰,陈天行,翟衡,杜远鹏
(山东农业大学园艺科学与工程学院,山东泰安 271018)
左山一葡萄与SO4及101-1杂交F1代部分植株抗旱性鉴定
郭淑华,刘笑宏,牛彦杰,陈天行,翟衡,杜远鹏*
(山东农业大学园艺科学与工程学院,山东泰安 271018)
本文对左山一葡萄为母本,SO4为父本杂交的5个子代株系(A15、A17、A34、A35、A48)和左山一葡萄为母本,101-1为父本杂交的2个子代株系(B24、B26)的一年生扦插苗的抗旱性进行评价。以砧木1103P以及克瑞森葡萄为对照,杂交株系的盆栽土壤保持相对含水量为45%的中度干旱水平,对照盆栽保持土壤相对含水量为80%(各试材均设置未经水分胁迫的自身对照),以检测电导率、相对含水量、根系活力、叶片厚度、丙二醛、叶绿素、过氧化氢酶、过氧化物酶等相关抗旱指标,并对其抗旱能力初步评价及分级。结果表明:绝大部分杂交后代具有较强的抗旱能力,其中抗旱性极强的为砧木1103P;抗旱性强的为A17、A15;抗旱性一般的为B24、A48、B26、A34;抗旱性较弱的类型为A35、克瑞森。
葡萄;杂交砧木;抗旱性
我国干旱、半干旱地区约占国土总面积的二分之一[1],三分之一以上的葡萄种植区位于干旱和半干旱地区[2]。一直以来干旱就是世界性的话题,干旱胁迫对果树产生的影响是国内外许多学者探讨的热点问题。干旱影响植物生长发育的各个阶段,如营养生长、生殖生长及生理代谢过程,是制约葡萄产业发展的重要环境因子之一[3]。2015年胶东半岛长达半年的干旱,导致酿酒葡萄新梢生长量明显不足,葡萄减产三分之一甚至更多。因此选育抗旱砧木及品种是保证我国葡萄产业可持续发展的重要措施。
欧美国家在葡萄野生资源研究利用领域的工作开展的比较早,无论是砧木还是栽培品种均可用种间杂交的方法培育新品种,且成果显著。葡萄野生种以东亚种群和北美种群为主[4],目前各国常用的抗根瘤蚜砧木基本以河岸葡萄、沙地葡萄及冬葡萄为亲本,抗性较强的种间杂交种栽培品种如威代尔、白维拉、摩尔多瓦、诺顿等采用美洲种与欧亚种杂交育成。我国在葡萄野生资源方面的研究主要用于栽培育种,如山欧杂种,而用于砧木育种的非常少见。山葡萄(Vitis amurensis Rupr)是我国珍贵的抗寒葡萄资源,其抗寒性可达-39.8 ℃[5],但不抗根瘤蚜[6]。而国外引进的砧木抗根瘤蚜能力较强,但抗寒性不足,因此本试验选择山葡萄中栽培性状较好、抗寒性强的优良品种左山一,与国外引进的砧木SO4及101-1进行杂交,从杂交子代中选择抗寒性较好的砧木进行抗旱能力评价,以期丰富杂交砧木的生态抗逆性数据资料。
1 材料与方法
1.1 试验材料
以左山一葡萄为母本,SO4为父本杂交的5个子代株系(A15、A17、A34、A35、A48)和以左山一葡萄为母本,101-1为父本杂交的2个子代株系(B24、B26)为试材,采用双对照,即以砧木中抗旱性很强的1103P和栽培品种中抗旱性较强的品种克瑞森葡萄为对照。所有试材均是一年生扦插苗。
1.2 试验设计与方法
于2015年4~8月在山东农业大学葡萄园防雨棚内采用盆栽进行试验。当年扦插苗定植于直径14 cm、高18 cm的塑料盆,每盆装基质-沙-土1∶1∶1(基质为珍珠岩、蛭石与草炭的混合物,土为壤土)混合物3.1 kg,保持土壤相对含水量为45%的中度干旱水平[7](对照盆栽保持土壤相对含水量为80%),采用称重法控水并记录,每天下午6点补充水分,每试材设3个重复,每个重复2株葡萄苗。干旱胁迫25 d之后,取组织进行各个指标的测定。
参照《植物生理学实验指导》的方法测定叶片相对含水量、相对电导度、丙二醛含量;用紫外分光光度法测叶绿素、根系活力、CAT;用愈创木酚法测定POD。
分别于干旱处理后的第7天、第15天、第21天、第25天测定荧光参数,利用连续激发式荧光仪(Handy PEA,Hansatech,英国)测定快速叶绿素荧光诱导曲线。从OJIP曲线上可直接获得如下参数:Fo为最小荧光(20 μs);Fk为K点(300 μs)的荧光;Fj为J点(2 ms)的荧光;Fi为I点(30 ms)的荧光;Fm为0.3~2 s之间的最大荧光;Ft为任意时刻的荧光数值。通过JIP-test分析得到的OJIP曲线,可获得如下参数:J点相对可变荧光Vj=(Fj-Fo)/(Fm-Fo);光合性能指数PIABS=Vj/(1-Vj);PSII最大光化学效率Fv/ Fm=φPo=TRo/ABS=[1-(Fo/Fm)]。
采用英国Hansatech公司的FMS-2型便携脉冲调制式荧光仪测定,测定程序如下:首先对胁迫前的叶片进行30 min暗适应,打饱和脉冲光(12 000 μmol/m2•s),测定暗适应下的最大荧光Fm。对光适应下的叶片先打60 s作用光(与各处理光强一致),打极弱的(<0.05 μmol/m2•s)光测得叶片最小荧光(Fo'),再打饱和脉冲光,测定光适应下的最大荧光值Fm',各荧光参数意义及计算公式如下:PSII光下实际光化学效率ФPSII=(Fm'-Fs)/Fm';PSII激发压1-qP=1-(Fm'-Fs)/(Fm'-Fo)。
按以下标准对植株进行旱害分级,0级:植株生长发育与未处理对照无明显差异;1级:植株叶片有较轻微萎蔫;2级:植株叶片中度萎蔫;3级:植株叶片严重萎蔫;4级:植株叶片部分脱落;5级:植株叶片全部脱落。
1.3 各株系抗旱性评定
对试验结果进行隶属函数[8]分析,对9种株系抗旱性进行综合评价,从而比较各品种的抗旱性强弱。对已测指标用下式求出各株系各性状的具体函数值:
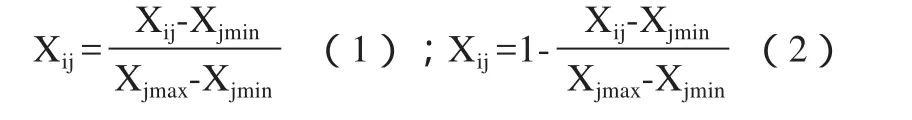
Xij为i品种j性状的隶属函数值,Xjmin为各品种j性状的最小值,Xjmax为各品种j性状的最大值。当j性状与植物的抗旱性成正相关时用(1)式,当j性状与植物抗旱性成负相关时用(2)式。
把每个品种各个性状的具体抗旱隶属值进行累加,并求出平均值Xi,Xi为i品种的抗旱隶属函数平均值,Xi越大,抗旱性越强;反之越弱。
1.4 数据处理
采用Microsoft Excel 2010软件处理数据和制图,采用DPS 7.05对数据进行方差分析和差异显著性检验。
2 结果分析
2.1 抗旱性表观症状及分级
对试材干旱胁迫至25 d之后,观察各试材的表观症状及旱害情况。
克瑞森与A35植株大部分叶片焦枯并脱落,旱害级数为4级;A34、B26植株叶片严重萎蔫,旱害级数为3级;A15、A17、B24、A48的旱害级数为2级;1103P则为1级。
2.2 干旱胁迫下各株系葡萄叶片Fv/Fm、ФPSII、1-qP、PIABS的变化
干旱胁迫下各株系的PSⅡ激发压(1-qP)均随时间延长呈增高趋势,说明干旱胁迫会增加反应中心关闭程度,导致受体接受电子的能力减弱[9]。如图1所示:随着胁迫时间的延长,各株系叶片的最大光化学效率(Fv/Fm)、实际光化学效率(ФPSII)和PIABS均逐渐降低,1103P的变化幅度较小,而克瑞森则出现较大的降低幅度。在杂交砧木中A15的Fv/Fm、ФPSII、PIABS和1-qP均呈现最小的变化幅度,说明A15的光合性能较强。A17的Fv/Fm,A35的ФPSII,A34的PIABS,A48的1-qP也表现较小的变化幅度。
2.3 干旱胁迫下各株系生理生化指标的测定
2.3.1 对各株系相对含水量、电导率的影响
从表1的数据可以看出,供试材料中克瑞森叶片含水量最低,低于90%,其他砧木均在90%~95%之间;干旱处理后降低了各植株叶片相对含水量,以杂交砧木A35、克瑞森和A34降低量最高,分别达18.83%、17.90%、16.5%,其他砧木降低量均在11%以下;干旱胁迫后克瑞森叶片相对含水量低于70%,其次为A34和A35,含水量在70%~80%之间,其他砧木含水量均在80%~90%之间。
电导率的大小直接反应植物的抗性,伤害愈重,外渗愈多,电导度的增加也就越大。从表1可以看出,克瑞森在正常生长情况下叶片电导率明显低于其他几个株系,电导率变化幅度最小的依次为1103P、A15、B24、A35、A48,其余几个株系干旱处理和对照相比变化幅度均大于0.3。
2.3.2 对各株系叶片厚度、叶绿素含量的影响
由表2可看出,供试品种中克瑞森和A15叶片厚度较薄,干旱胁迫后,葡萄叶片厚度均呈下降趋势,其中A15叶片厚度下降幅度最小,其次为1103P、A34和克瑞森,下降幅度均在0.02以下;叶绿素是反映光合作用强弱的重要指标,干旱胁迫降低了叶片的叶绿素含量,克瑞森和B26叶片叶绿素含量降低量最大,为0.127,而1103P、A15、A34和A48的叶绿素降低量均在0.1以下。
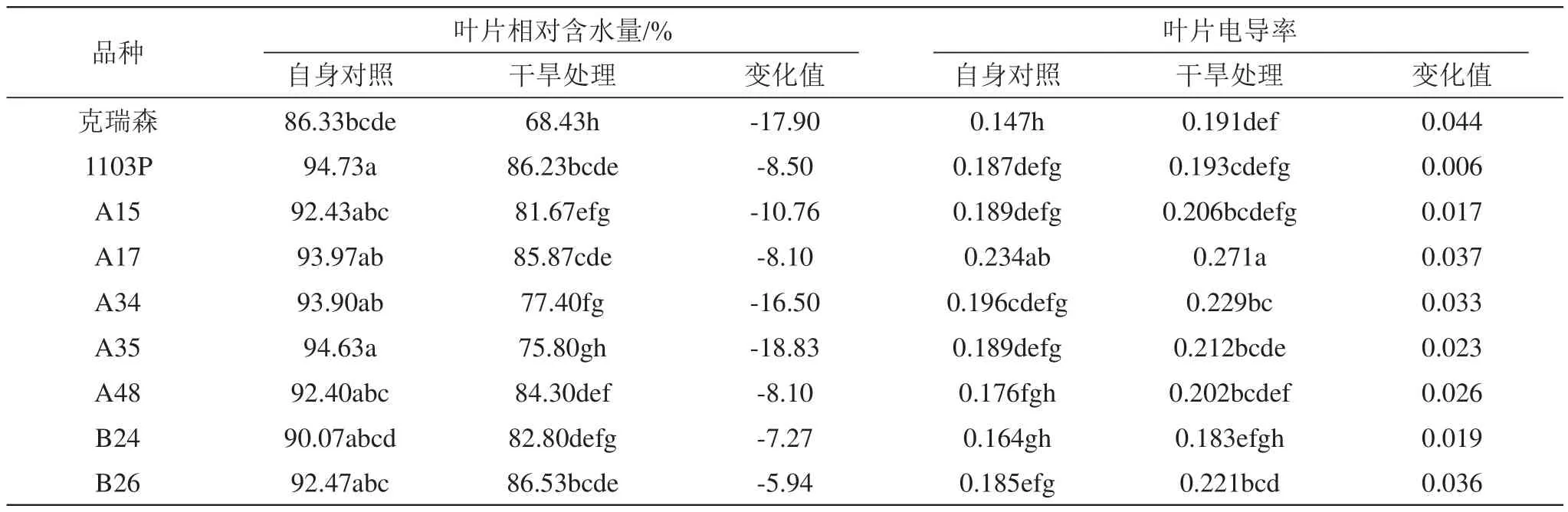
表1 干旱胁迫对各株系叶片相对含水量、电导率的影响
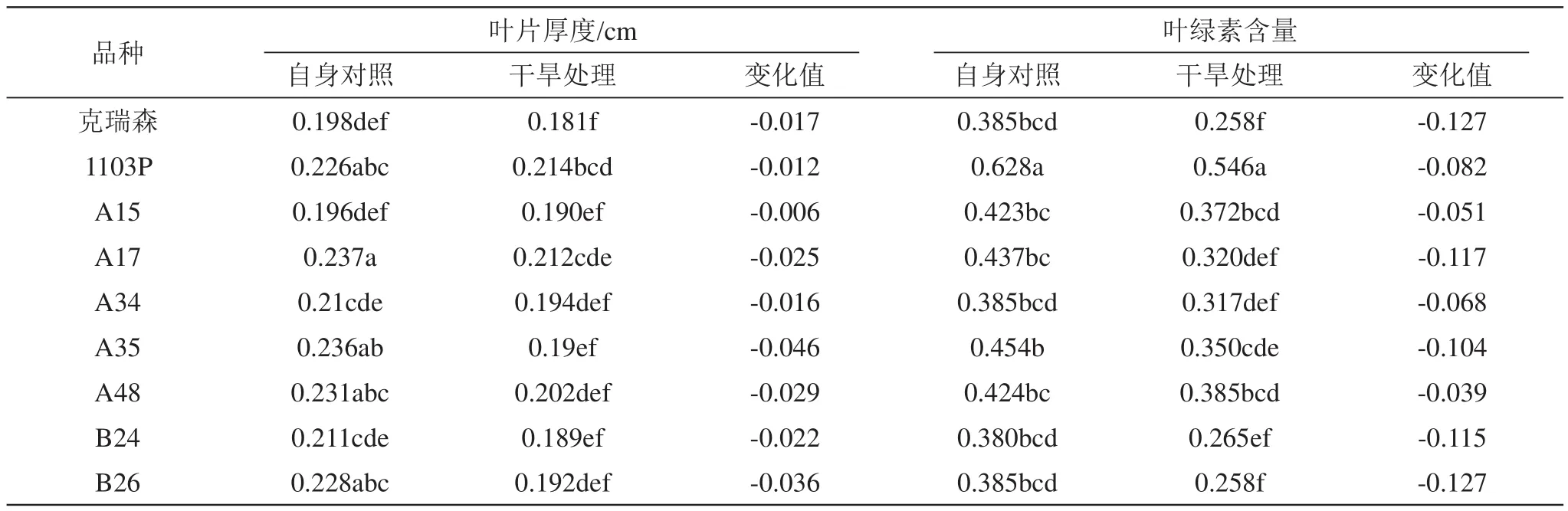
表2 干旱胁迫对各株系叶片厚度、叶绿素含量的影响
2.3.3 对各株系丙二醛含量、根系活力的影响
植物器官在逆境下受到伤害时,往往会产生丙二醛,伤害越重,产生的丙二醛越多,而丙二醛的积累可能对膜和细胞造成一定的伤害。由表3可见,各株系的丙二醛含量差别很大,正常水分供应下的株系几乎没有丙二醛的产生。干旱胁迫后丙二醛含量呈显著升高趋势,以1103P变化幅度最小,仅为2.68,其次为A15、A17、A48和B26,变化幅度均低于10。克瑞森变化幅度最大,为17.21。
当土壤干旱时,作物根系首先感受到胁迫并迅速发出信号,使整个植株对干旱做出反应,所以根系活力也是反映各株系抗旱性能的一个重要指标[10]。胁迫处理前,克瑞森葡萄和B26根系活力较低,均为21.53%;干旱胁迫后,葡萄根系活力下降,1103P和A15根系活力维持在20 μg/h•gFW以上,而克瑞森和B26根系活力较低,均为11.33 μg/h•gFW。
2.3.4 对各株系CAT、POD含量的影响
过氧化氢酶(CAT)是植物抗氧化保护系统中的另一重要酶,能清除逆境中产生的H2O2[11]。CAT活性与抗旱性呈正相关[12]。如表4所示,1103P的CAT活性变化幅度最小,为0.65 Δ240/min•gFW,其次为A48、A34,变化量分别为0.99 Δ240/min•gFW和1.04 Δ240/min•gFW,B24的CAT活性变化幅度最大,为4.49 Δ240/min•gFW。
过氧化物酶(POD)可以清除体内过多的H2O2等过氧化物,是植物细胞内防御酶系统中体内自由基的清除剂。由表4可看出,A35的POD活性变化幅度最小,为0.35 Δ470/min•gFW,克瑞森变化幅度最大,为3.68 Δ470/ min•gFW。
2.4 各株系抗旱性综合评价
为了综合评价各抗旱指标,本试验采用模糊隶属函数法对这9种株系所测得的8种指标进行分析(表5),并对这9种株系的抗旱性进行综合评价(表6),得出9种株系的抗旱性顺序为1103P>A17>A15>B24>A48>B26>A34>A35>克瑞森。
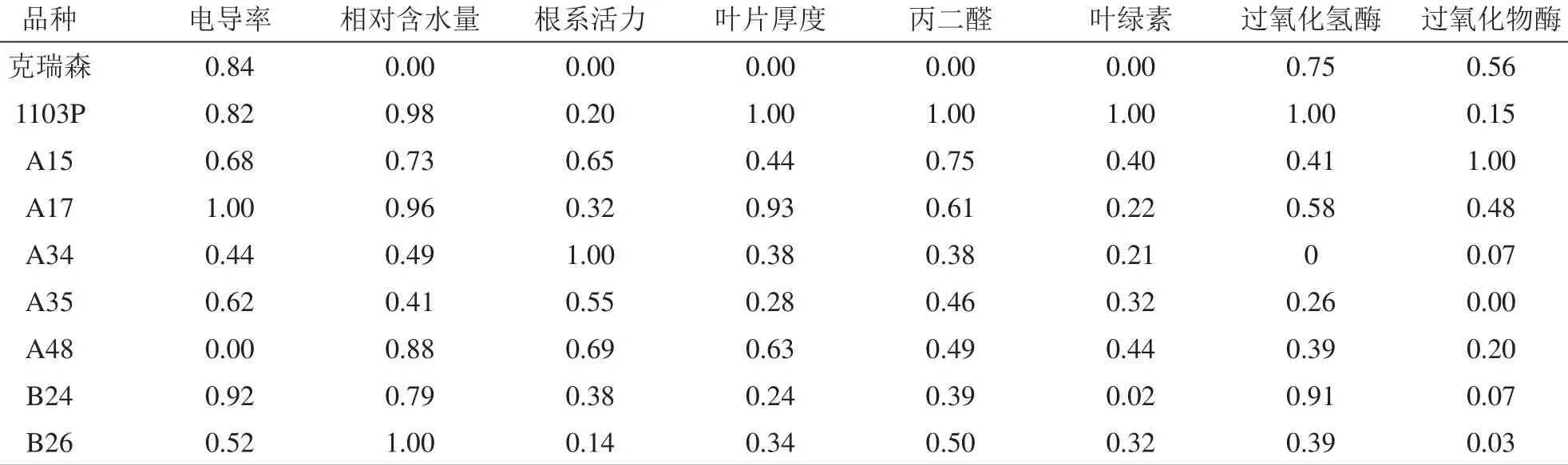
表5 不同株系抗旱指标隶属函数值

表6 不同株系平均隶属函数值及抗旱性排序
3 讨论
葡萄的抗旱机制是一个复杂的生理生化变化过程,涉及多种复杂的代谢过程,受多种因素影响,单一依靠一种或少数几个生理指标作为抗旱性强弱鉴定的依据具有片面性,得出的结论也是不太可靠的,并且很难反映其抗旱本质[13]。本试验通过检测叶绿素和电导率得出的结论就有差别,因此本试验采用模糊隶属函数法对各株系进行综合指标抗旱性鉴定,认为该方法消除了若干指标间的差异[14],能够较为准确地评价葡萄砧木的抗旱性。本研究利用隶属函数法最终将各品种/株系划分为4个抗旱类型:抗旱性极强为砧木1103P;抗旱性强为A17、A15;抗旱性一般的为B24、A48、B26、A34;抗旱性较弱的类型A35、克瑞森。
在现有的葡萄砧木中,1103P的极强抗旱能力是世界公认的[15-17],国内的研究也有大量证明[18-20],而克瑞森是美国为干旱地区培育的鲜食栽培品种,是欧亚种中比较抗旱的品种,但和砧木相比,抗旱性还是有一定差距[21-22]。本试验以抗旱性能好的砧木1103P和抗旱性较强的栽培品种克瑞森作为对照,结果显示所有杂交苗的抗旱性能均比克瑞森强,尤其是A15、A17、A48和B26,平均隶属值均在0.5以上,接近1103P(平均隶属值为0.83),只有B26的平均隶属值与克瑞森相当,但仍高于克瑞森,说明本试验所有砧木抗旱性均较强。
[1] 汤章城. 植物对水分胁迫的反应和适应性-Ⅱ植物对干旱的反应和适应性[J]. 植物生理学通讯, 1983, 4(7): 1-7.
[2] 苏来曼, 艾则孜, 木合塔尔•扎热, 等. 两个无核葡萄品种叶绿素荧光特征参数对不同程度土壤干旱的响应[J]. 新疆农业科学, 2015, 52(8): 1447-1453.
[3] 陈文荣, 曾玮玮, 李云霞, 等. 高丛蓝莓干旱胁迫的生理响应及其抗旱性综合评价[J]. 园艺学报, 2012, 39(4): 637-646.
[4] MULLINS M G, BOUQUET A, WILLIAMS L E. Biology of the grapevine[M]. Cambridge University Press, 1992.
[5] 宋润刚, 路文鹏, 王军, 等. 山葡萄种间杂交选育酿造葡萄品种的途径及其效果[J]. 中国农业科学, 1998, 31(5): 48-55.
[6] 杜远鹏, 翟衡, 王忠跃, 等. 葡萄根瘤蚜抗性砧木研究进展[J].中外葡萄与葡萄酒, 2007(3): 25-29.
[7] 井大炜, 邢尚军, 杜振宇, 等. 干旱胁迫对杨树幼苗生长、光合特性及活性氧代谢的影响[J]. 应用生态学报, 2013, 24(7): 1809-1816.
[8] 张文娥, 王飞, 潘学军. 应用隶属函数法综合评价葡萄种间抗寒性[J]. 果树学报, 2007, 24(6): 849-853.
[9] 孙永江, 付艳东, 杜远鹏, 等. 不同温度/光照组合对‘赤霞珠’葡萄叶片光系统II功能的影响[J]. 中国农业科学, 2013, 46(6): 1191-1200.
[10] 景蕊莲, 胡荣海. 作物抗旱性的根系研究[J]. 国外农学: 麦类作物, 1995(3): 37-39.
[11] 赵薇, 惠竹梅, 林刚, 等. 硒对水分胁迫下赤霞珠葡萄幼苗叶片生理生化指标的影响[J]. 果树学报, 2011, 28(6): 984-990.
[12] 许宏, 綦伟, 翟衡. 葡萄砧木及品种抗旱鉴定初报[J]. 中国果树, 2004(2): 30.
[13] 白志英, 李存东, 刘渊. 干旱胁迫下小麦叶片脯氨酸和蛋白质含量变化与染色体的关系[J]. 植物遗传资源学报, 2007, 8(3): 325-330.
[14] 范晓丹, 刘飞, 王衍安, 等. 不同苹果砧木对缺锌胁迫的耐性评价[J]. 应用生态学报, 2015, 26(10): 3045-3052.
[15] MALSINA M, SMART DR BAUERLE T, HERRALDE F D, et al. Seasonal changes of whole root conductance by a drought-tolerant grape root system[J]. Journal of Experimental Botany, 2011, 62(1): 99-109.
[16] KOUNNDOURAS S, TSIALTAS I T, ZIOZIOU E, et al. Rootstock effects on the adaptive strategies of grapevine (Vitis vinifera L. cv. Cabernet-Sauvignon) under contrasting water status: Leaf physiological and structural responses[J]. Agriculture Ecosystems & Environment. 2008, 128(1/2): 86-96.
[17] PARANYCHIANAKIS N V, NIKOLANTONAKIS M, SPANAKIS Y, et al. The effect of recycled water on the nutrient status of Soultanina grapevines grafted on different rootstocks[J]. Agricultural Water Management. 2006, 81(1/2):185-198.
[18] 綦伟, 谭浩, 翟衡. 干旱胁迫对不同葡萄砧木光合特性和荧光参数的影响[J]. 应用生态学报. 2006, 17(5): 835-838.
[19] 孙茜, 任磊, 马文婷, 等. 四种葡萄砧木抗旱性的鉴定[J]. 湖北农业科学, 2015, 54(3): 623-626.
[20] 陈继峰. 国外抗旱葡萄砧木品种简介[J]. 中国果树, 2001(6): 55.
[21] CONESA M R, ROSA J M D L, DOMINGO R, et al. Changes induced by water stress on water relations, stomatal behavior and morphology of table grapes (cv. Crimson Seedless) grown in pots[J]. Scientia Horticulture, 2016, 202(1): 9-16.
[22] 严大义, 罗树祥, 赵常青. 克瑞森无核葡萄引种研究初报[J].中国果树, 2005(6): 41-43.
Drought tolerance of F1hybrid progenies of V. amurensis Zuoshanyi × SO4 and Zuoshanyi × 101-1
GUO Shuhua, LIU Xiaohong, NIU Yanjie, CHEN Tianxing, ZHAI Heng, DU Yuanpeng*
(College of Horticultural Science and Engineering, Shandong Agricultural University, Tai'an 271018, Shandong China)
One year old F1hybrid progeny (A15, A17, A34, A35, A48) of ♀V. amurensis Zuoshanyi×♂SO4 and two clones (B24, B26) of ♀V. amurensis Zuoshanyi×♂101-1 were used as materials to test their drought stress tolerance. Strong drought stress tolerance of potted rootstock 1103P and V. vinifera Crimson Seedless were used as double controls. The materials were treated with moderate water stress, the soil relative water content (SRWC) was maintained at about 45%. SRWC of control was maintained at about 80%. With three replicates for each treatment the leaf relative water content, electrical conductivity, leaf thickness, chlorophyll content, MDA content, root activity, CAT and POD activity were tested after 25 days of drought stress treatment. Results indicated that the clones A17 and A15 had strong drought resistance, but they were still weaker than 1103P. The drought resistance level of B24, A48, B26 and A34 were medium, and they were all better than Crimson Seedless. A35 showed similar drought resistance with Crimson Seedless.
grape; hybrid stock; drought tolerance
S663.1
A
10.13414/j.cnki.zwpp.2017.02.001
2016-10-24
国家葡萄产业技术体系(CARS-30);教育部长江学者和创新团队发展计划创新团队(IRT15R42)
郭淑华,硕士,研究方向为葡萄抗逆生理。E-mail: 641358683@qq.com
*通讯作者:杜远鹏,副教授,E-mail: duyuanpeng001@163.com
猜你喜欢
水利水电快报(2023年2期)2023-02-21 02:34:54
亚热带植物科学(2022年1期)2022-05-17 12:39:32
今日农业(2021年14期)2021-11-25 23:57:29
今日农业(2020年13期)2020-12-15 09:08:51
四川农业科技(2019年5期)2019-07-01 09:46:46
河南水利年鉴(2017年0期)2017-05-19 02:29:16
浙江农业学报(2017年1期)2017-05-17 06:13:36
西南农业学报(2016年4期)2016-05-17 05:42:13
浙江柑橘(2016年1期)2016-03-11 20:12:31
山西农经(2015年7期)2015-07-10 15:00:57
