
大野荞根和茎的解剖结构及其黄酮组织化学定位研究
2017-03-30 05:38:15陈庆富
广东农业科学 2017年12期
唐 链,陈庆富
(贵州师范大学荞麦产业技术研究中心,贵州 贵阳 550001)
荞麦为蓼科(Polygonaceae)荞麦属(Fagopyrum Mill)植物,别名为乌麦、三角麦、花荞、荞子等。全世界已报道并正式命名的荞麦有24个种,主要分布于亚洲、欧洲和美洲,为一年生、多年生草本,少数种为半灌木。其中,多年生荞麦的主要类型为金荞复合物(F.cymosum complex),包括二倍体毛野荞(F. pilus QF Chen)、二倍体大野荞(F. megaspartanium QF Chen)和异源四倍体金荞(F. cymosum Meissn)3个种[1]。金荞复合物具有营养和保健的双重功效,块根可入药,清热解毒、排脓祛瘀[2],且在生产上易于人工栽培管理、生长快、产量高、病虫害少,是一种分布广泛的植物[3-6]。大野荞是其中最主要和常见的种类[1]。目前关于大野荞的研究主要集中在同工酶[7]、黄酮[8]、蛋白质[9]、可溶性糖[10]等方面,而有关大野荞营养器官的解剖学研究尚未见报道。
金荞复合物经药理和临床证实有保肝、抗菌、镇痛、抗炎、抗肿瘤、抗氧化等作用,能有效地控制和治疗糖尿病,预防和控制高血压、心血管硬化疾病、健胃消食,增加机体对疾病的免疫能力,同时对儿童的生长和智力的开发也有帮助[3,11]。其中黄酮类化合物是金荞复合物中最有效、含量最多的药用成分。关于荞麦黄酮类物质在荞麦植株组织器官中的分布尚未见报道。本研究采用植物解剖学和组织化学方法,对大野荞根、茎、根茎部的显微结构特征及黄酮类化合物在各器官中的分布情况进行分析,旨在揭示大野荞营养器官的结构特征与黄酮的积累规律,为进一步研究大野荞中有效成分的累积、变化,以及荞麦属植物的起源与进化研究提供理论依据。
1 材料与方法
1.1 试验材料
2014年8月5日,按照株间距5 cm、行间距30~40 cm的规格,将大野荞播种于贵州师范大学荞麦产业技术研究中心柏杨基地,进行常规栽培管理,于2014年10月18日采集其健康植株进行试验。
1.2 试验方法
1.2.1 解剖学方法 (1)石蜡切片法:选取生长健康的新鲜大野荞为材料,取成熟叶片、根茎部及一年生和多年生根与茎,将根、茎和根茎部切成长约0.4~0.7 cm的小段,迅速放入FAA固定液中固定[12],经系列浓度乙醇脱水、透蜡、包埋、HHQ-2508B轮转式切片机切片,切片厚度为8~12 μm,以番红固绿双重染色,中性树胶封片,在OLYMPUS BX51显微镜下观察,并用与镜体相连的OLYMPUS DP70显微镜数码相机拍照。
(2)徒手切片法:以大野荞新鲜根、茎、根茎部、叶为材料,分别用双面刀片切成薄片,放置于载玻片上,滴加少许清水,盖上盖玻片制成临时装片,在OLYMPUS BX51-DP70显微镜下观察并拍照。
1.2.2 组织化学方法 (1)氢氧化钠法:对大野荞新鲜根、茎、根茎部和叶进行徒手切片,放置于载玻片上,在切片上滴加5% NaOH溶液[13],染色10 min,使切片中的总黄酮显色,制成临时装片,立即置于OLYMPUS BX51-DP70显微镜下观察并拍照。
(2)荧光法:对大野荞新鲜根、茎、根茎部和叶进行徒手切片,放置于载玻片上,滴加1%醋酸镁甲醇溶液[14],染色30 s,使切片中的总黄酮显色,制成临时装片,立即置于BXURA2荧光显微镜下(激发波长405~480 nm)观察并拍照。
2 结果与分析
2.1 根的解剖结构
大野荞植株根的横切面近圆形,根的初生结构均由表皮、皮层以及中柱共同构成(图1A、B,封二)。表皮位于根的最外方,由一些体积较小、长方形的细胞组成,排列规则、紧密,无细胞间隙。部分表皮细胞的外壁向外延伸,形成根毛。表皮之内为皮层,皮层由多层体积较大、近椭圆形的薄壁细胞组成(图1C,封二),薄壁细胞内含有少量晶体。中柱位于内皮层之内,由中柱鞘、初生韧皮部、形成层、初生木质部以及髓组成。中柱鞘是中柱外的一圈细胞,由体积较小、近圆形的薄壁细胞构成,排列紧密。初生木质束和韧皮部束相间排列。初生木质部为四原型,初生结构中导管为环纹增厚(图1D,封二)。初生木质部和韧皮部发育方式均外始式。木质部的内侧为髓,由排列疏松的薄壁细胞组成,有晶体分布。木质部脊对应的中柱鞘细胞为侧根原基发生处(图1A,封二)。
当根的初生生长结束后,进入次生生长。根次生结构从外到内都是由周皮和次生维管组织组成。周皮由木栓层、木栓形成层和栓内层组成,细胞壁栓质化,染色较深,排列紧密(图1E,封二)。次生维管组织发达,次生木质部由导管、木射线、木薄壁细胞、木纤维等组成(图1F、G,封二)。其中导管为网纹和孔纹增厚(图1H,封二),单个或数个聚生存在,呈放射状分布。木纤维细胞壁木质化加厚,常聚集成束分布于导管周围(图1I,封二)。木薄壁细胞、韧皮薄壁细胞和髓薄壁细胞中通常含有内含物。次生韧皮部由韧皮纤维、筛管、伴胞、韧皮薄壁细胞、韧皮射线等组成(图2A,封二)。韧皮纤维呈多边形,常成群分布(图2B,封二)。韧皮射线较宽大,经形成层与木射线连接,形成维管射线。
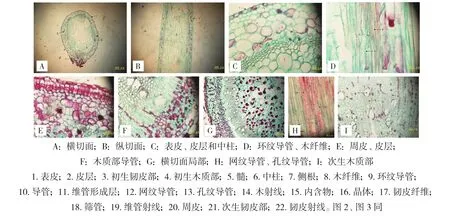
图1 大野荞1年生根解剖结构
随着根年限的增长,多年生主根逐渐增粗,其内部结构与一年生根的类似,也是由周皮和次生维管组织构成,在各类细胞的组成和排列上基本相同,但在细胞的数量上有所增加(图2C,封二)。随着根不断加粗,维管形成层和周皮的层数也增多(图2D,封二)。多年生与一年生根相比,最显著的是产生次生木质部的细胞数量远大于次生韧皮部(图2E,封二),因此根的增粗主要是通过次生木质部细胞数目的增多。多年生根的木薄壁细胞、木射线及髓薄壁细胞里含有丰富的内含物(图2F,封二),而一年生根里所含较少,木纤维增加,维管射线也较一年生根明显。
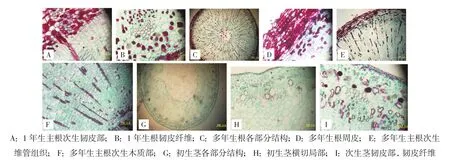
图2 大野荞一年生和多年生根茎解剖结构
2.2 茎的解剖结构
茎的解剖结构从外到内依次为表皮、皮层、维管束和髓(图2G,封二)。表皮由一层体积较小、长方形的细胞组成,排列紧密,有少量的气孔和表皮毛,具角质层。皮层最外面紧贴表皮的为几层厚角组织,内方为近排列疏松的椭圆形薄壁细胞,薄壁细胞里有大量的内含物。维管束的大小不一,大维管束与小维管束交替排列。维管束由初生韧皮部、束中形成层和木质部构成,为外韧型维管束(图2H,封二)。导管口径较小,无规则排列。韧皮纤维的细胞壁增厚,常聚集成束分布在韧皮部的最外侧(图2I,封二)。维管束之间有薄壁细胞构成的初生髓射线,连接皮层和髓部。幼茎的中央为髓,由近圆形的薄壁细胞组成,含有晶体,有明显的胞间隙。
次生茎的木质部发达,其所占比例较韧皮部大,由导管、木纤维、木薄壁组织组成(图3A,封二)。导管口径较大,为环纹增厚(图3B,封二)。茎的髓部成熟较早,在茎的生长发育过程中,髓薄壁细胞随着茎的生长而不断被破损,因此在成熟茎的中央通常会形成髓腔。
2.3 黄酮的组织化学定位
黄酮类化合物能与氢氧化钠水溶液发生显色反应,在光学显微镜下观察,切片中的黄酮类化合物会呈现出黄色至橙色的颜色变化。在根中,周皮、韧皮部和初生木质部被染成橙黄色,维管形成层几乎全部呈浅黄色,含量从外到内逐渐降低(图3C,封二)。在茎中,表皮、棱角处的厚角组织细胞、皮层薄壁细胞、韧皮薄壁细胞、束中形成层、束间形成层、木薄壁细胞均被染成黄色或浅黄色(图3D,封二)。
黄酮类化合物能与醋酸镁甲醇溶液发生反应,在荧光显微镜下可见蓝白色荧光。在根中,主要在周皮、韧皮纤维、木质部处显现荧光(图3E,封二);在茎中,主要在表皮、韧皮纤维、束中形成层、束间形成层、木质部处显现荧光(图3F,封二);其他部位的荧光很微弱。从根和茎的着色面积来看,茎的着色面积比根大。
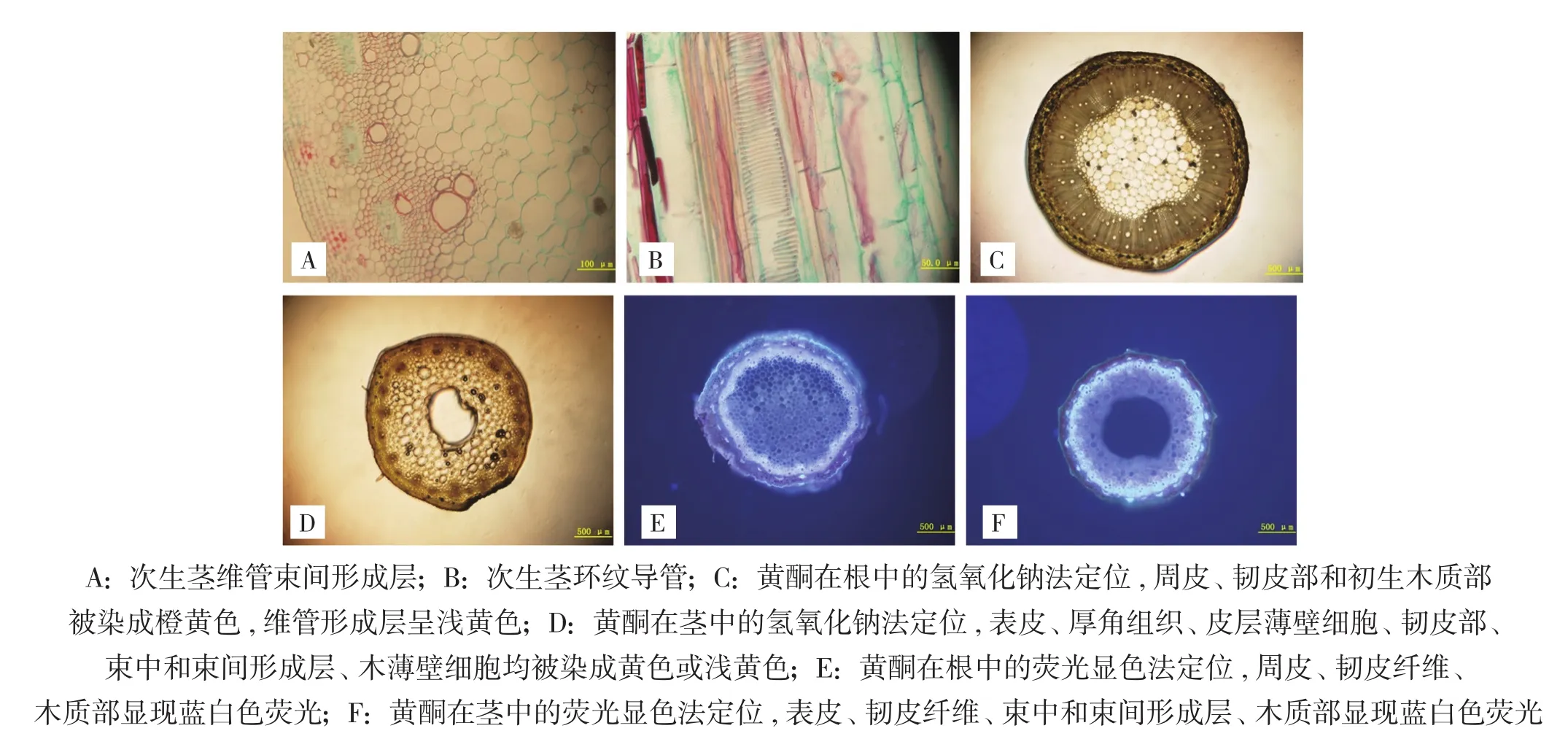
图3 大野荞茎解剖结构与根、茎黄酮定位
3 结论与讨论
李淑久等[15]、慕勤国[16]、游亚丽[17]、赵钢等[18]对荞麦的营养器官进行了形态解剖学研究。李淑久等[15]对甜荞、苦荞、金荞麦和齿翅野荞麦的营养器官形态与解剖学比较研究,结果表明4种荞麦主根结构基本相同,为四原型,无髓。而慕勤国[16]在苦荞和甜荞的根中观察到髓,后因细胞破损而变成空腔。游亚丽[17]、赵钢等[18]研究表明荞麦根属于典型双子叶植物根的构造,中柱中央是由薄壁细胞组成的髓,薄壁细胞内含晶体。本研究结果表明,大野荞的根为四原型,髓部发达,髓薄壁细胞含大量晶体,这与前人的研究结果相似。
李淑久等[15]认为甜荞、苦荞、金荞麦和齿翅野荞麦4种荞麦茎的解剖结构基本相似,表皮上有单细胞乳突毛,有花外蜜腺。皮层由厚角组织、基本组织构成。维管束为外韧型,髓射线较宽,导管类型大体相同。慕勤国[16]研究发现苦荞茎表皮外具角质层,维管束26~40个,大小不等。本研究表明,大野荞茎由表皮、皮层、维管束和髓构成,表皮外具角质层,为外韧型维管束,这与李淑久等[15]、慕勤国[16]的研究结果一致。
黄酮类化合物是一大类天然酚性化合物,具有重要药理作用,在植物界分布较为广泛。荞麦中的黄酮类化合物是荞麦的主要药用成分,许多药理作用都与该成分直接相关,它具有抗菌、降血糖、控制高血压和心血管硬化疾病、增强机体的免疫力等作用[19-21]。运用组织化学定位方法来研究有效药用成分在植物器官和组织中的分布,是一种有效的研究方法。廖云海等[22]用氢氧化钠和三氯化铝乙醇溶液对光果甘草营养器官中黄酮类物质进行组织化学研究,结果表明黄酮类物质主要分布在根的周皮,茎的周皮、韧皮部和髓部粘液细胞,叶的表皮细胞、厚角组织、腺毛和叶肉中的胶囊细胞中。葛楚源等[23]用氢氧化钠和醋酸镁甲醇溶液对草珊瑚叶、茎中黄酮类物质进行组织化学研究,发现黄酮类化合物主要分布在叶的表皮、厚角组织、类栅栏组织、维管束、分泌细胞,茎的表皮、厚角组织、分泌细胞、韧皮部中。
到目前为止,没有发现荞麦黄酮类物质在荞麦植株不同组织器官中分布规律方面的研究报道。本研究首次对荞麦黄酮物质在大野荞的根、茎中的分布进行了研究,发现在大野荞的根和茎均有黄酮的分布,在根中,主要分布在周皮、韧皮部和维管形成层附近的薄壁细胞中;在茎中,主要分布在表皮、厚角组织、皮层薄壁细胞、韧皮薄壁细胞、束中形成层、木薄壁细胞中;茎中的着色面积比根中大。上述结果与其他植物上的报道如廖云海等[22]、葛楚源等[23]的研究结果部分相似。从显色面积来看,茎中的着色面积比根中大,因此可以推断大野荞茎的黄酮含量比根高,这与胡鞒缤等[24]用紫外分光光度法对荞麦植株各部分的总黄酮含量的测定结果一致,都是茎的黄酮含量高于根的黄酮含量。
关于木质部导管的增厚方式,初生木质部中最早形成的导管通常是环纹加厚,以后依次产生的是螺纹、梯纹、网纹、孔纹等加厚方式,一般认为孔纹加厚是次生木质部中导管的特征[25]。钟心尧[26]对3种越橘茎的比较解剖学研究结果表明,次生木质部导管为梯纹和孔纹增厚。本研究结果显示,大野荞根初生结构中导管为环纹增厚,次生木质部导管为网纹和孔纹增厚,在茎中导管为环纹增厚,这与前人的研究结果相似。
本研究结果表明,大野荞木质部发达,导管口径较大,有利于水分运输,并且在次生木质部薄壁细胞、射线细胞及髓薄壁细胞常积累丰富的内含物,这些特征说明其对营养物质的运输和贮存能力较强,这与金荞麦块根有较高的药用价值相关[17]。周皮的细胞层数较多,能保护根部抵御干旱环境[27]。在周皮及次生韧皮部含有少量晶体,晶体的存在可以增加机械性能,还可避免对细胞的毒害作用、改变细胞的渗透势、缓解逆境胁迫等作用[28-29]。这些结构可帮助大野荞适应干旱的环境,从而维持植物正常的生理生态机能。
[1]陈庆富. 荞麦属植物科学[M]. 北京:科学出版社,2012:3-20.
[2]中国植物志编辑委员会. 中国植物志(第25卷第1分册)[M]. 北京:科学出版社,1998:111-112.
[3]张以忠,陈庆富. 荞麦研究的现状与展望[J].种子,2004:23(3):39-42.
[4]周小理,李宗杰,周一鸣. 荞麦治疗糖尿病化学成分的研究进展[J]. 中国粮油学报,2011(5):119-121.
[5]赵钢,唐宇,王安虎. 金荞麦的营养成分分析及药用价值研究[J]. 中国野生植物资源,2002(5):39-41.
[6]凌永霞,黄凯丰,陈庆富. 2种提取方法测定大野荞植物叶中的黄酮含量[J]. 安徽农业科学,2011,39(13):7767-7770.
[7]张以忠,陈庆富. 荞麦属植物三叶期幼叶酯酶同工酶研究[J]. 武汉植物学研究,2008(4):428-432.
[8]冯晓英,陈庆富. 大野荞不同器官中总黄酮含量的测定[J]. 贵州农业科学,2007(4):15-16.
[9]王燕,陈庆富. 甜荞·苦荞和大野荞的高分子量种子蛋白亚基研究[J]. 安徽农业科学,2010(33):18678-18680.
[10]邓琳琼,张以忠. 大野荞不同器官可溶性糖的抗氧化能力[J]. 食品研究与开发,2016(7):23-27.
[11]张美莉,胡小松. 荞麦生物活性物质及其功能研究进展[J]. 杂粮作物,2004(1):26-29.
[12]李正理. 植物制片技术[M]. 北京:科学出版社,1978:129-137.
[13]谭玲玲,胡正海,蔡霞,等. 北柴胡营养器官中主要化学成分的组织化学定位及其含量比较[J]. 分子细胞生物学报,2007,40(4) :214-221.
[14]Ni X L,Peng L,Liu W Z. Structures,components and functionsof secretory tissues in Houttuynia cordata[J]. J Integrative PlantBiol,2007,49 (12):1734.
[15]李淑久,张惠珍,袁庆军,等. 四种荞麦营养器官的形态学与解剖学比较研究[J]. 贵州农业科学,1992(5): 9-14.
[16]慕勤国. 苦荞营养器官的解剖学研究[J]. 西北植物学报,1994,14(6):138-140.
[17]游亚丽. 金荞麦和荞麦的生物学特征比较研究[D]. 合肥:安徽师范大学,2012.
[18]赵钢,彭镰心,向达兵,等. 荞麦栽培学[M].北京:科学出版社,2015:31-32.
[19]冯黎莎,陈放,白洁. 金荞麦的抑菌活性研究[J]. 植物科学学报,2006,24(3):240-244.
[20]王元福,陈晓玉,韩淑英,等. 甜荞麦叶总黄酮镇痛抗炎作用的实验研究[J]. 上海中医药杂志,2004,38(11):54-55.
[21]熊建新,王志路,韩淑英,等. 甜荞麦种子提取物对糖尿病大鼠血浆及肾组织糖基化终产物的影响:剂量依赖性效应[J]. 中国组织工程研究,2005,9(23):138-139.
[22]廖云海,陆嘉惠,李娜,等. 光果甘草营养器官结构及其总黄酮的组织化学定位和含量研究[J]. 西北植物学报,2010,30(12):2406-2411.
[23]葛楚源,陈文列,李钻芳,等. 草珊瑚植物叶、茎显微结构与黄酮组织化学定位研究[J]. 中国中药杂志,2012,37(4):438-441.
[24]胡鞒缤,姚瑛瑛,李艳琴,边俊生,陕方. 荞麦植株各部位总黄酮含量的测定与比较[J]. 食品与药品,2013(6):394-396.
[25]K伊稍. 种子植物解剖学[M]. 李正理译. 上海:上海科学技术出版社,1982:161-174.
[26]钟心尧. 3种越橘茎的比较解剖学研究[D]. 长春:吉林农业大学,2014.
[27]郑丽,蔡霞,胡正海. 狭叶柴胡根的发育解剖学研究[J]. 植物研究,2009,29(6):659-644.
[28]李芳兰,包维楷. 植物叶片形态解剖结构对环境变化的响应与适应[J]. 植物学通报,2005,22(S):118-127.
[29]韦存虚,张军,王建军,等. 星星草营养器官适应盐胁迫的结构特征[J]. 植物资源与环境学报,2006,15(1):51-56.
猜你喜欢
作品(2022年10期)2022-10-18 10:24:18
作品(2022年6期)2022-06-30 09:45:38
孙子研究(2021年1期)2021-05-12 01:01:36
植物研究(2021年2期)2021-02-26 08:40:10
东坡赤壁诗词(2020年4期)2020-09-02 13:53:46
东坡赤壁诗词(2018年2期)2018-05-10 11:08:24
六盘山(2016年3期)2016-06-07 19:24:56
天津农业科学(2015年11期)2015-12-03 00:03:49
江苏农业科学(2015年4期)2015-06-15 21:11:35
安徽农学通报(2014年7期)2014-04-29 03:00:24
