
一株能拮抗3种土传病害病原真菌的长枝木霉
2017-03-23 03:46杨兴堂姜传英姚志红任龙辉张荣沭
草业科学 2017年2期
常 媛,杨兴堂,姜传英,姚志红,贾 让,任龙辉,张荣沭
(东北林业大学园林学院,黑龙江 哈尔滨 150040)
一株能拮抗3种土传病害病原真菌的长枝木霉
常 媛,杨兴堂,姜传英,姚志红,贾 让,任龙辉,张荣沭
(东北林业大学园林学院,黑龙江 哈尔滨 150040)
为了丰富生防木霉菌(Trichodermaspp.)种资源,在园林植物八宝景天(Sedumspectabile)根际土壤中分离得到一株优势木霉。根据该菌株菌落的形态特征、形态学显微观察、菌株rDNA-ITS序列分析以及邻接法同源性比对等结果判定木霉菌株种类。采用平板对峙法将该菌株与3种土传病害病原菌[(Sclerotiniasclerotiorum)、立枯丝核菌(Rhizoctoniasolani)和细链格孢菌(Alternariaalternata)]进行对峙培养。并用该木霉诱导山新杨(Populusdavidiana×P.albavar.Pyramidalis)组培苗,观察离体叶片抵抗细链格孢菌侵染的能力。结果表明,此木霉为长枝木霉(Trichodermalongibrachiatum),命名为Tl-70。该菌株对3种土传病害病原真菌均有较明显的拮抗作用。对核盘菌的抑菌率最高为77.71%,显著高于对细链格孢菌和立枯丝核菌的抑制率(P<0.05);对立枯丝核菌的抑菌率为58.56%,显著高于对细链格孢菌的抑制率(P<0.05);对细链格孢菌的抑菌率最低,为53.32%。并且,该木霉能提高山新杨组培苗叶片拮抗细链格孢菌侵染的能力。说明此菌株是具有潜力的生防菌株。
木霉菌;形态特征;ITS序列分析;系统发育树;拮抗作用;山新杨
木霉菌(Trichodermaspp.)是世界上公知的、重要的植物真菌病害防治菌,它的生防优势在于不仅具有杀真菌与促生长的作用,还具有改良土壤等多种功效[1-5]。既能防治农作物及林木生长期真菌引起的病害,还能防治采摘后的水果蔬菜及园艺花卉材料的存储期病害[6-8];同时还具有诱导植物免疫反应、降解土壤中的盐类化合物、农药残留物及重金属等功能[4,9-12]。因而,对木霉菌的研究越来越受到重视。
木霉菌广泛存在于土壤及其它基质中, 其生存能力强, 适应性广[13]。赵晓燕等[14]在中国13个省(自治区)的采集总土样647份,分离的木霉菌株总数为827株,并发现在土壤中比水样中更容易分离出木霉菌[14]。景天属植物以其观赏价值高、抗旱、耐寒、耐贫瘠性强、少病虫害、维护成本低、繁殖容易等优点,成为节水、节能、节材、节力型生态园林建设不可或缺的优良地被植物[15-16]。八宝景天(Sedumspectabile),是景天科景天属多年生宿根肉质草本植物,主要分布于东北、华北、华东及华中各地[14]。对八宝景天的研究主要集中在生药学、植物源农药和观赏绿化等方面[17]。鉴于八宝景天具有多种抗逆特性,期望在八宝景天根际土壤中可以分离获得能拮抗常见土传植物病害的木霉菌株,为生物防治植物土传病害提供物质资源。核盘菌(Sclerotiniasclerotiorum)、立枯丝核菌(Rhizoctoniasolani)和细链格孢菌(Alternariaalternata)是常见的土传植物病害病原菌,能引发多种林木、农作物和观赏植物的枯萎病、根腐病、立枯病和猝倒病等病害[18-21],本研究鉴定了一株从八宝景天根际土壤中分离获得的木霉菌,采用平板对峙法观察该菌株对3种土传病原菌的抑制效果;并用该木霉诱导山新杨(Populusdavidiana×P.albavar.Pyramidalis),观察木霉诱导对山新杨拮抗细链格孢菌侵染能力的影响,以期为确定该菌株的生防作用提供依据。
1 材料与方法
1.1 材料
八宝景天根际土壤: 采集于哈尔滨市道里区兆麟公园内步行道路边,光照充足。将八宝景天全株拔出,剪下根部,置于无菌PE袋中,置于冰盒带回实验室,备用。
供试病原菌:核盘菌、立枯丝核菌和细链格孢菌均由东北林业大学林学院刘志华博士提供。
1.2 方法
1.2.1 菌株的分离与纯化 将八宝景天带土的根部放入无菌蒸馏水中,按照刘志华等[22]的植物根际木霉分离方法进行分离,将获得的菌落在无菌条件下分别单独接种于PDA(potato dextrose agar)平板培养基(直径9 cm)上,25 ℃恒温全光照培养。待刚长出菌丝时,挑取菌丝接种在新的PDA培养基上。如此操作,直至获得无杂菌污染的单菌落。最后,将纯化菌株转接到PDA斜面培养基中,4 ℃储存留做鉴定。
1.2.2 菌株的形态学鉴定 将待鉴定菌株接种到PDA平板上,置于25 ℃恒温培养,观察菌落形态和颜色。另紧贴平板内缘在培养基上平铺一张盖玻片,待菌丝生长覆盖盖玻片后,取盖玻片使用光学显微镜观察菌丝隔膜、分生孢子梗、瓶梗和分生孢子等显微结构的形态,并在TrichodermaandHypocreaTaxonomy网站(http://nt.ars-grin.gov/taxadescriptions/keys/TrichodermaIndex.cfm)与木霉的形态特性进行比较,从形态上对菌株进行初步鉴定。
1.2.3 菌株的分子生物学鉴定
1)收集菌株的菌丝体:将待鉴定菌株的分生孢子(1×104cfu·mL-1200 μL)在无菌条件下接种至PDB培养基内,25 ℃,180 r·min-1振荡培养4 d后,用安子旋等[23]的方法收集菌丝体。将获得的菌丝置于液氮中速冻,置于-80 ℃冰箱中备用。
2)菌株的基因组rDNA提取、ITS序列扩增及测序:采用CTAB法[24]提取木霉菌的基因组rDNA,利用通用引物对ITS1/ITS4扩增ITS片段。引物序列为,ITS1:5’-TCCGTAGGTGAACCTGCGG-3’;IST4:5’-TCCTCCGCTTATTGATATGC-3’。采用TaKaRa Ex Taq®(宝生物工程(大连)有限公司)试剂盒,按照说明书步骤进行PCR扩增反应。PCR反应体系为50 μL。PCR扩增条件:95 ℃预变性5 min;95 ℃变性30 s,55 ℃退火30 s,72 ℃延伸1 min,35个循环;72 ℃延伸5 min,4 ℃保存。通过琼脂糖凝胶电泳分析PCR扩增产物,使用Trans2k Plus DNA Marker(Beijing Transgen Biotech Co., Ltd.)指示扩增产物分子量的大小。采用Promega Wizard SV Gel and PCR Clean-up System (Promega)试剂盒,按照说明书步骤回收PCR产物。序列测定由英潍捷基(上海)贸易有限公司完成。
3)基因比对及进化树分析:将测得的序列提交到ISTH网站(http://www.isth.info/)进行TrichOKEY分类鉴定,来确定待鉴定菌株的菌种。并将测序结果在TrichoBLAST的ITS Database数据库中进行比对。用MEGA6.06软件与已知菌种ITS序列作同源性比较,并对检索获得的同源性较近的菌株采用邻接法(Neighbor-Joining, NJ)构建系统发育树(Bootstrap=1 000),分析亲缘关系和系统发育。
1.2.4 平板对峙培养 先将已鉴定的木霉菌株与3种病原真菌同步活化4 d,至菌落分布均匀,用直径6 mm的打孔器打取菌碟,试验组分别将木霉菌和每种病原菌菌碟同时接种在内径9 cm的PDA平板上,菌碟圆心相聚5 cm,连线经过平板圆心;对每种病原菌设置对照组,接种方法同试验组,不接种木霉菌。接种后的平板静置于培养箱内,25 ℃培养144 h。每个对峙试验做3个重复。在96和144 h观察木霉菌与病原菌的生长状况,在144 h测量病原菌对照菌落半径和病原菌落指向木霉菌的半径,按下式计算待鉴定菌株对3种病原真菌的抑菌率[25]。
抑菌率=[(病原菌对照菌落半径-病原菌落指向木霉菌的半径)×病原菌对照菌落半径-1]×100% 。
1.2.5 木霉菌与山新杨组培苗互作后拮抗细链格孢菌的能力 山新杨组培苗按照吕曼曼等[26]的方法繁殖。配制浓度为1×105cfu·mL-1的Tl-70的分生孢子溶液。取200 μL该溶液加入到装有20 mL WPM液体培养基的试管中,将山新杨组培苗根部在无菌条件下植入,进行Tl-70与杨树苗根组织互作,设为处理组;同时,以只加200 μL无菌水培养基培养的山新杨组培苗为对照,处理和对照组均在温度为26 ℃的生物培养箱中培养24 h,此时可见山新杨根部已缠绕Tl-70的菌丝体。
无菌条件下,分别取处理和对照组中山新杨大小相近的成熟叶片,置于9 cm的无菌平皿中完全湿润的滤纸上。在叶片相同位置上接种直径为5 mm的细链格孢菌菌饼,封口后置于温度为26 ℃的生物培养箱中培养,分别以没接种病原菌的木霉处理过的和未处理的叶片为对照,每处理3次重复,每个培养皿中3个叶片。观察接种病原菌5 d后叶片染病情况。
1.3 数据分析
测得的数据采用Microsoft Office Excel 2007软件包及Minitab 16统计软件包进行分析处理。对不同菌株抑菌率的差异显著性(P<0.05)进行分析。
2 结果与分析
2.1 菌株形态鉴定
在八宝景天根际土壤中共分离得到27个木霉菌落,对生长在平板PDA培养基上的这些单菌落按形态特征进行比较鉴别,共有5种木霉,分别有11、6、5、3、2个菌落。本研究选取得到11个菌落的一种木霉(优势木霉)进行菌种鉴定和拮抗作用研究。为了后续研究及保存,将此菌株命名为Tl-70。
25 ℃全光照条件下,木霉Tl-70的菌落生长初期菌丝白色,继续生长产生分生孢子,分生孢子绿色,以接种点为圆心呈同心纹状分布,分生孢子堆呈黑绿色(图1 A)。平板背面可见以接种点为中心产生黄色的可扩散色素(图1 B)。菌落生长迅速,有明显的轮纹,周围有白色菌丝的生长带,最后整个菌落全都变成黑绿色。显微(100×)观察可见(图1 C):分生孢子椭圆形或细长椭圆形,光滑,大小为(2.0~3.2) μm×(1.5~2.3) μm。典型的分生孢子梗包括发育强壮的中轴,在中轴上向顶端产生单生的瓶梗,随着离顶端距离的延伸,产生成对的长度逐渐增加的二次分枝;瓶梗直接着生在二次分枝上。瓶梗主要为单生,典型形状为圆柱形,呈涡状排列时中间部分膨大,直或弯曲成钩状。参照TrichodermaandHypocreaTaxonomy网站对长枝木霉菌落形态分类鉴定的描述,初步确定为长枝木霉(T.longibrachiatum)。
2.2 菌株分子鉴定
木霉Tl-70的ITS基因片段PCR产物经凝胶电泳检测,条带清晰单一,大小约600bp。扩增产物经测序,结果长度为637 bp,与木霉ITS序列长度理论值一致。

图1 观察的Tl-70菌落形态和显微形态特性
注:A为平板培养72-144 h的菌落形态;B为平板背面,示黄色素;C为显微(100×)结构:a,分生孢子;b,瓶梗;c,分生孢子梗;d,菌丝隔膜。
Note: A, Colonial morphology of Tl-70 cultured for 72 to 144 h (obverse side); B, reverse sides of dishes of Tl-70 (shows yellow pigment, cultured from 72 to 144 h); C, micro-morphology of Tl-70 observed using optical microscopy (100× magnification). a, Conidium; b, phialides; c, conidiophore; d, Hyphae septum.
将木霉Tl-70的ITS基因片段测序结果提交到ISTH网站(http://isth.info/)的TrichOKEY中进行比对鉴定。共发现5个特异种源标记(anchors),位置分别在Tl-70序列的第74、98、280、438和532碱基处。种属鉴别为长枝木霉(T.longibrachiatum)或东方肉座菌(Hypocreaorientalis)。种属鉴定的可靠性均为高。
将木霉Tl-70的ITS基因序列提交到ITS Database中进行TrichoBLAST同源性搜索(Expect=0.0),获得数据库中一致性高的ITS1和ITS2序列共11条,分值范围在882~735。这些序列与Tl-70的ITS基因序列在覆盖序列长度为445~437 bp范围内的一致性在96~100%,Tl-70的ITS基因序列与长枝木霉(T.longibrachiatum, ATCC18648)的一致性最高,达到100%(表1)。而与东方肉座菌(H.orientalis, GJS88-81)的一致性为99%。因此,综合在TrichOKEY和TrichoBLAST中比对的结果,判定Tl-70为长枝木霉。
将木霉Tl-70和在ISTHDatabase中搜索到的与其亲缘关系近的11个菌种的ITS基因序列作同源性比较,并构建系统进化树(图2)。上述12种菌株聚类为2组,Tl-70位于Ⅱ组,与长枝木霉ATCC18648(Z31019)和东方肉座菌H.orientalis为一组。Tl-70与长枝木霉ATCC18648(Z31019)的亲缘关系最近;有研究者[8]认为,东方肉座菌包含长枝木霉集合种的一部分,遗传上H.orientalis与T.longibrachiatum非常接近,本研究鉴定结果与此一致。然而,木霉Tl-70与红褐肉座菌(H.jecorina)/李氏木霉[T.reesei(ATCC13631)]、栗褐肉座菌(H.schweinitzii)、橘绿木霉[T.citrinoviride(CBS258.85)]、木霉MA3642和加纳木霉T.ghanense(GJS95-137)的亲缘关系最远。
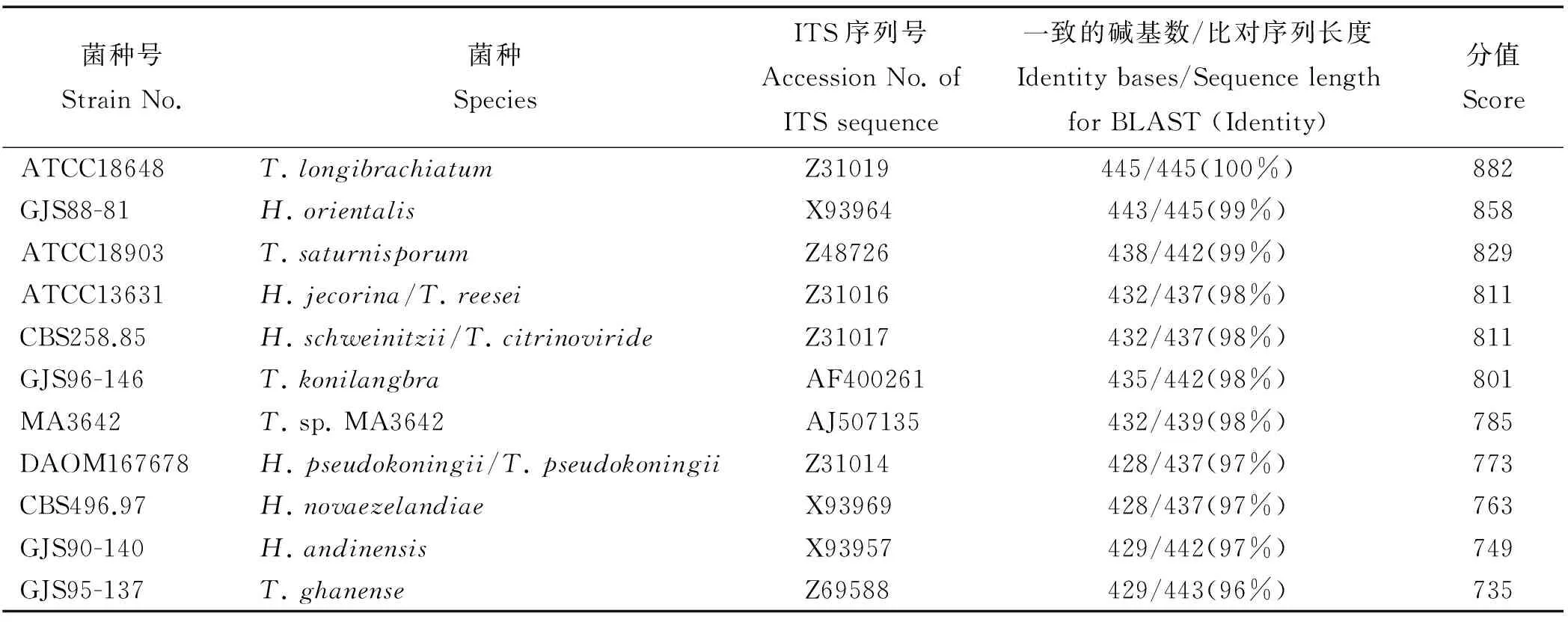
表1 用Tl-70的ITS基因序列进行同源性搜索获得的同源菌种及信息

图2 Tl-70菌株与高同源性菌株ITS基因序列的系统进化树
2.3 平板对峙培养及抑菌率分析
对峙培养结果发现:木霉Tl-70对3种病原菌均具有拮抗作用。在对峙培养96 h时,病原菌菌落均已与Tl-70的菌落边缘发生接触,但3种病原菌生长受到木霉菌抑制的程度不同(图3)。观察发现,3种病原菌菌丝生长速度为立枯丝核菌>核盘菌>细链格孢菌(图3a、b、c)。细链格孢菌生长速度最慢,对峙培养中细链格孢菌菌落周围尚有木霉菌丝没有蔓延过去的空间,这或许是细链格孢菌能分泌某种挥发性物质,影响了Tl-70的生长和产孢(图3d)。同时,由于立枯丝核菌生长速度最快,与Tl-70对峙培养时先占据了平板的大部分营养和空间,使Tl-70生长空间受到侵占,2个菌株的菌落边缘已完全接触,Tl-70的产孢量与图3d和f相比最少。但是,在Tl-70与立枯丝核菌交界处已见少量木霉菌分生孢子寄生在立枯丝核菌的菌丝体上,说明Tl-70菌株具有重寄生作用(图3e)。此外,尽管核盘菌的菌丝生长较快,但其与Tl-70对峙培养96 h时,已被限制在较小的营养和空间内,说明Tl-70与核盘菌对峙时呈优势生长(图3f)。
对峙培养144 h时,病原菌菌落所占据的空间与96 h时观察的结果相比均已缩小,并且Tl-70的产孢量在增加、分生孢子的颜色变成深绿色(图3j、k、l)。由Tl-70与细链格孢菌的对峙培养可见,Tl-70菌株占据了空余的营养和环境空间,细链格孢菌的生长受到限制。Tl-70对细链格孢菌抑菌作用最弱,抑菌率为53.32%(图3f)。同时发现,Tl-70已经控制立枯丝核菌生长,使其不再扩大生长,Tl-70对立枯丝核菌的抑菌率为58.56%,方差分析结果显示显著高于对细链格孢菌的抑菌率(P<0.05)(图3 k)。此外,优势生长的Tl-70菌株已将核盘菌限制在一个很小的空间区域内,对核盘菌的抑菌率最高,为77.71%,显著高于对立枯丝核菌和细链格孢菌的抑菌率(P<0.05)(图3l)。
2.4 木霉Tl-70与山新杨组培苗互作后拮抗细链格孢菌的能力
在Tl-70处理和未处理的山新杨组培苗叶片上接种细链格孢菌3 d时,可见细链格孢菌菌丝开始生长。但对照组叶片比处理组菌丝生长早,且病斑出现快。接种细链格孢菌5 d时,与Tl-70互作后杨树苗叶片上的病斑明显小于对照叶片上的病斑(图4 C和D)。对照组比处理组叶片上病斑的平均半径约大1.7 mm(n=9),存在显著差异(P<0.05)。说明与Tl-70互作能增强山新杨对细链格孢菌侵染的抵抗能力。
3 讨论与结论
本研究根据木霉Tl-70菌落的形态特征、形态学显微观察、菌株rDNA-ITS序列分析以及邻接法同源性比对等结果进行分类鉴定,确定分离获得的木霉为长枝木霉。观察发现,该菌株对3种土传病害病原真菌均有较明显的拮抗作用。对核盘菌的抑菌率最高为77.71%,显著高于对细链格孢菌和立枯丝核菌的抑制率(P<0.05);对立枯丝核菌的抑菌率为58.56%,显著高于对细链格孢菌的抑制率(P<0.05);对细链格孢菌的抑菌率最低,为53.32%。此外,Tl-70诱导山新杨组培苗后能增强其叶片对细链格孢菌侵染的抵抗能力。说明木霉Tl-70是具有潜力的生防菌株。
长枝木霉作为木霉属中重要的一种,能对小麦叶斑病菌(Bipolaristriticicola)、小麦赤霉病菌(Fusariumsporotrichioides)、南瓜叶枯病菌(A.cucumerina)、黄瓜枯萎病菌(F.oxysporum)、西瓜枯萎病菌(F.oxyspornum)、大豆根腐病菌(F.solani)、百合枯萎病菌(F.oxysporum)、棉花立枯病菌(R.solani)、苹果树腐烂病菌(Valsaceratosperma)、甘草叶斑病菌(A.azukiae)、独活叶斑病菌(Septoriadearnessii)和大黄叶斑病菌(Ascochytarhei)12种植物病原菌生长具有抑制作用[27],已经引起国内外很多学者的重视[28]。本研究分离得到长枝木霉菌Tl-70,将为开发木霉生防制剂提供物质基础。

图4 组培苗叶片拮抗细链格孢菌能力
注:A,对照组叶片;B,木霉处理组叶片;C,对照组叶片接种细链格孢菌;D,木霉处理组叶片接种细链格孢菌
Notes:A) Leaves of untreated tissue-cultured seedlings (control group), B) leaves of seedlings inoculated with Tl-70 (T group),C) leaves of control group infected withAlternariaalternata, and D) leaves of T group infected withAlternariaalternata
研究发现,木霉菌生物防治机制存在多种形式,包括定殖在植物根际的木霉菌通过对营养和空间的竞争来抑制病原真菌的生长和繁殖[27],木霉菌对病原真菌的重寄生[28-29],木霉菌发酵产生抗生素等活性物质来抑制或杀死病原真菌[29-30],及木霉菌促进植物生长诱导植物系统抗病性等[31-32]。其中竞争、重寄生、发酵产物杀菌等机制是生防菌与病原微生物单一的作用过程。平板对峙培养法便于观察拮抗菌占领营养和空间的能力、重寄生或溶菌现象。Tl-70在72 h时已占据整个培养皿的空间并产生大量的分生孢子(图1)。而这3种病原菌菌丝在96 h时还没有占据整个培养皿的生长空间(图3)。因此,相对于病原菌,Tl-70具有很好的营养和空间竞争优势。然而,本研究发现,木霉Tl-70和3种不同病原菌对峙培养144 h内,Tl-70菌丝生长、产孢速度和最终对病原菌产生的拮抗效果不同。与细链格孢菌对峙,菌丝生长、产孢速度慢于与核盘菌的对峙培养,表现为逐渐占据大部分生长空间。而与核盘菌的对峙培养中,在144 h内一直表现出很好的竞争作用(图3)。在与立枯丝核菌对峙培养时,最初虽然没占据空间优势,在144 h时,Tl-70大量产孢。总之,尽管Tl-70占据空间的大小不同,但一直表现为良好的生长状态。虽然对峙培养不能全面反映田间施用拮抗菌的实际情况,但揭示了Tl-70对病原菌可能的拮抗作用和途径、以及木霉菌与病原菌接触时的生长状态,这些结果将对木霉Tl-70的开发应用起到积极的指导作用。
现有的研究[29-31]认为,木霉菌诱导植物产生系统抗性的机制涉及植物多种信号途径和代谢途径,十分复杂,尚未阐明。已有较多文献报道木霉诱导产生的植物系统抗病性主要依赖于jasmonate-ethylene途径,同时伴随多个抗病基因的短期表达[32-33]。而采用抑制消减杂交方法验证长枝木霉MK1能通过活跃活性氧信号途径和加固细胞壁来提高番茄(Solanumlycopersicum)对灰霉病菌(Botrytiscinerea)的防御能力[34]。将分离得到的长枝木霉Tl-70与洋葱(Alliumcepa)共生互作,通过影响洋葱的抗氧化能力、以及碳水化合物、木质素、硫同化的代谢途径,提升了洋葱生长速率和对尖孢镰刀菌(F.oxysporum)的防御能力[35]。以上研究表明,在不同的木霉菌株诱导不同植物提高其系统抗病性的过程中,可能既有共通机制,又有特异机制。本研究发现经长枝木霉Tl-70诱导后,山新杨组培苗叶片抵抗细链格孢菌侵染能力明显增强。长枝木霉Tl-70与山新杨互作后影响了哪些代谢途径,从而使其提高对细链格孢菌的抗性有待于深入研究。
References:
[1] Mukherjee P K,Horwitz B A,Herrera-Estrella A,Schmoll M,Kenerley C M.Trichodermaresearch in the genomeera.Annual Review of Phytopathology,2013,51:105-129.
[2] Olson H A,Benson D M.Induced systemic resistance and the role of binucleateRhizoctoniaandTrichodermahamatum382 in biocontrol ofBotrytisblight in geranium.Biological Control,2007,42(2):233-241.
[3] Druzhinina I S,Seidl-Seiboth V,Herrera-Estrella A,Horwitz B A,Kenerley C M,Monte E,Mukherjee P K,Zeilinger S,Grigoriev I V,Kubicek C P.Trichoderma:The genomics of opportunistic success.Nature Reviews Microbiology,2011,9(10):749-759.
[4] 李江遐,蒋立科.木霉菌在坪用黑麦草上的应用效果初报.草业科学,2006,23(1):103-105. Li J X,Jiang L K.Primary report of application effects ofTrichodermaon perennial ryegrass.Pratacultural Science,2006,23(1):103-105.(in Chinese)
[5] Hanada R E,Pomella A W V,Soberanis W,Loguercio L L.Biocontrol potential ofTrichodermamartialeagainst the black-pod disease (Phytophthorapalmivora) of cacao.Biological Control,2009,50(2):143-149.
[6] Wijesinghe C J,Wijeratnam R S W,Samarasekara J K R R,Wijesundera R L C.Biological control ofThielaviopsisparadoxaon pineapple by an isolate ofTrichodermaasperellum.Biological Control,2010,53(3):285-290.
[7] Chen L,Yang X,Raza W,Luo J,Zhang F,Shen Q.Solid-state fermentation of agro-industrial wastes to produce bioorganic fertilizer for the biocontrol ofFusariumwilt of cucumber in continuously cropped soil.Bioresource Technology,2011,102(4):3900-3910.
[8] 杨合同.木霉分类与鉴定.北京:中国大地出版社,2009:160-306. Yang H T.TrichodermaClassification and Identification.Beijing:China Land Press,2009:160-306.(in Chinese)
[9] Sant D,Casanova E,Segarra G,Aviles M,Reis M,Isabel-Trillas M.Effect ofTrichodermaasperellumstrain T34 onFusariumwilt and water usage in carnation grown on compost-based growth medium.Biological Control,2010,53(3):291-296.
[10] 闫峰,徐凤花,顾金刚,姜瑞波.木霉属真菌的生物降解及生物转化作用研究进展.微生物学杂志,2009,29(3):77-80. Yan F,Xu F H,Gu J G,Jiang R B.Research progress of biodegradation and biotransformation withTrichoderma.Journal of microbiology,2009,29(3):77-80.(in Chinese)
[11] 金潇,颜冬云,秦文秀.有机磷农药的微生物降解技术.湖南农业科学,2011(5):93-97. Jin X,Yan D Y,Qin W X.Technology of microbial degradation for organophosphorus pesticides.Hunan Agricultural Sciences,2011(5):93-97.(in Chinese)
[12] 杨振兴,田从魁,党晨原,常方,倪晋仁.真菌对重金属Pb(Ⅱ),Cd(Ⅱ),As(Ⅲ)和Cr(Ⅵ)耐受性的比较研究.北京大学学报:自然科学版,2015,51(4):667-675. Yang Z X,Tian C K,Dang C Y,Chang F,Ni J R.Comparative study on Pb(Ⅱ),Cd(Ⅱ),As(Ⅲ),Cr(Ⅵ) resistance characteristics of fungus.Acta Scientiarum Naturalium Universitatis Pekinensis,2015,51(4):667-675.(in Chinese)
[13] 姚彦坡,王雅玲,吕国忠.木霉对草坪上2种重要土传病害生防效果的研究.草业科学,2007,24(8):96-99. Yao Y P,Wang Y L,Lyu G Z.Research on preventing the turfgrass plant diseases withTrichodermaharzianumpreparation.Pratacultural Science,2007,24(8):96-99.(in Chinese)
[14] 赵晓燕,吴晓青,李哲,赵忠娟,陈泉,陈凯,李纪顺,杨合同.13个省、自治区滩涂湿地木霉菌分布初探.山东科学,2015,28(3):34-37. Zhao X Y,Wu X Q,Li Z,Zhao Z J,Chen Q,Chen K,Li J S,Yang H T.Trichodermadistribution of coastal beach and wetland of thirteen provinces and autonomous regions.Shandong Science,2015,28(3):34-37.(in Chinese)
[15] 张寅媛,刘英,白龙.干旱胁迫对4种景天科植物生理生化指标的影响.草业科学,2014,31(4):724-731. Zhang Y Y,Liu Y,Bai L.Effect of drought stress on physiological indexes of 4 Crassulaceae species.Pratacultural Science,2014,31(4):724-731.(in Chinese)
[16] 汤聪,刘念,郭微,蔡鑫,苏建华,刘萍.广州地区8种草坪式屋顶绿化植物的抗旱性.草业科学,2014,31(10):1867-1876. Tang C,Liu N,Guo W,Cai X,Su J H,Liu P.The drought stress tolerance of several lawn-style roof green species in Guangzhou area.Pratacultural Science,2014,31(10):1867-1876.(in Chinese)
[17] 赵成爱,余梅燕,吴现芳,杨琳琳.八宝景天乙醇提取物对菜青虫的生物活性.农药,2012,51(1):62-63,70. Zhao C A,Yu M Y,Wu X F,Yang L L.Bioactivity ofSedumspectabileBoreau extracts againstPierisrapae.Agrochemicals,2012,51(1):62-63,70.(in Chinese)
[18] Taylor J W,Jacobson D J,Kroken S,Kasuga T,Geiser D M,Hibbett D S,Fisher M C.Phylogenetic species recognition and species concepts in fungi.Fungal Genetics and Biology,2000,31(1):21-32.
[19] 李兴龙,李彦忠.土传病害生物防治研究进展.草业学报,2015,24(3):204-212. Li X L,Li Y Z.Research advances on biological control of soil-borne disease.Acta Prataculturae Sinica,2015,24(3):204-212.(in Chinese)
[20] 蒋家珍,赵美琦,薄怀霞,肖杰,李学锋.生物农药在草坪病害防治中的应用研究.草业科学,2004,21(11):93-95. Jiang J Z,Zhao M Q,Bao H X,Xiao J,Li X F.Study on application of biological fungicide to several turf diseases.Pratacultural Science,2004,21(11):93-95.(in Chinese)
[21] 张茹,李金华,柴兆祥,王蒂.甘肃河西马铃薯根际生防木霉菌对接骨木镰刀菌的拮抗筛选及鉴定.草业学报,2009,18(2):138-145. Zhang R,Li J H,Chai Z X,Wang D.Screening of biocontrolTrichodermafrom potato rhizosphere of Western Gansu againstFusariumsambucinumand their identification.Acta Prataculturae Sinica,2009,18(2):138-145.(in Chinese)
[22] 刘志华,范海娟,张荣沭,季世达.一种植物根际木霉菌分离方法:中国专利201510354447.3.2015-09-30 Liu Z H,Fan H J,Zhang R S,Ji S D.A strategy for seperating plant rhizosphericTrichoderma.China,Patent application receipted by State Intellectual Property Office of the People's Republic of China,2015,Accepted:201510354447.3.(in Chinese)
[23] 安子旋,吕曼曼,朱国栋,张佩佩,常媛,刘志华,张荣沭.液体培养真菌菌丝体及发酵液的高效分离方法.贵州农业科学,2015,43(6):98-102. An Z X,Lyu M M,Zhu G D,Zhang P P,Chang Y,Liu Z H,Zhang R S.The collection method of fungal mycelium of liquid culture and its fermentation broth.Guizhou Agricultural Scicnces,2015,43(6):98-102.(in Chinese)
[24] Pan H,Yang C P,Wei Z G,Jiang J.DNA extraction of birch leaves by improved CTAB method and optimization of its ISSR system.Journal of Forestry Research,2006,17(4):298-300.
[25] 张丽荣,康萍芝,沈瑞清.木霉菌对土传病害病原真菌的拮抗作用.内蒙古农业科技,2007(5):48-50. Zhang L R,Kang P Z,Shen R Q.Antagonistic effects ofTrichodermaagainst soil-borne pathogenic fungi.Inner Mongolia Agricultural Science and Technology,2007(5):48-50.(in Chinese)
[26] 姜传英,朱国栋,姚志红,杨兴堂,刘志华,张荣沭.3个棘孢木霉菌株对杨树组培移栽苗生长和光合特性的影响.植物研究,2016,33(6):1189-1199. Jiang C Y,Zhu G D,Yao Z H,Yang X T,Liu Z H,Zhang R S.Effects of threeTrichodermaasperellumstrains on the growth and photosynthetic charateristics of tissue-culturedPopulusdavidiana×P.albavar.pyramidalisseedlings.Pratacultural Science,2016,33(6):1189-1199.(in Chinese)
[27] Morán-Diez E,Hermosa R,Ambrosino P,Cardoza R E,Gutierrez S,Lorito M,Monte E.The ThPGI endolygalacturonase is required for theTrichodermaharzianum plant beneficial interaction,Nolecalar plant-Microbe Interactions,2009,22(8):1021-1031.
[28] 李纪顺,陈凯,杨合同,黄玉杰,张广志.木霉抗生性代谢产物研究进展.农药,2010,49(10): 713-716. Li J S,Chen K,Yang H T,Huang Y J,Zhang G Z.Bioactive secondary metabolites fromTrichodermaspecies.Agrochemicals,2010,49(10):713-716.(in Chinese)
[29] Liu Z H,Yang X X,Sun D M,Song J Z,Chen G,Juba O,Yang Q.Expressed sequence tags-based identification of genes in a biocontrol strainTrichodermaasperellum.Molecular Biology Reports,2010,37(8):3673-3681.
[30] Seidl V,Song L,Lindquist E,Gruber S,Koptchinskiy A,Zeilinger S,Schmoll M,Martinez P,Sun J,Grigoriev I,Herrera-Estrella A,Baker S E,Kubicek C P.Transcriptomic response of the mycoparasitic fungusTrichodermaatrovirideto the presence of a fungal prey.BMC Genomics,2009,10(12):1-13.
[31] Sun P X,Zheng C J,Li W C,Jin G L,Huang F,Qin L P.Trichodermanin A,a novel diterpenoid from endophytic fungus culture.Journal of Natural Medicines,2011,65(2):381-384.
[32] Shoresh M,Yedidia I,Chet I.Involvement of jasmonic acid/ethylenesignaling pathway in the systemic resistance induced in cucumber byTrichodermaasperellumT203.Phytopathology,2005,95:76-84.
[33] Korolev N,David D R,Elad Y.The role of phytohormones in basal resistance andTrichoderma-induced systemic resistance toBotrytiscinereainArabidopsisthaliana.BioControl,2008,53:667-683.
[34] De Palma M,D’Agostino N,Proietti S,Bertini L,Lorito M,Ruocco M,Caruso C,Chiusano M L,Tucci M.Suppression Subtractive hybridization analysis provides new insights into the tomato (SolanumlycopersicumL.) response to the plant probiotic microorganismTrichodermalongibrachiatumMK1.Journal of Plant Physiology,2016,190:79-94.
[35] Abdelrahman M,Abdel-Motaal F,El-Sayed M,Jogaiah S,Shigyo M,Ito S,Tran L S.Dissection ofTrichodermalongibrachiatum-induced defense in onion (AlliumcepaL.) againstFusariumoxysporumf.sp. cepa by target metabolite profiling.Plant Science,2016,246:128-138.
(责任编辑 苟燕妮)
ATrichodermalongibrachiatumstrain with antagonistic effects against three soil-borne pathogenic fungi
Chang Yuan, Yang Xing-tang, Jiang Chuan-ying, Yao Zhi-hong, Jia Rang, Ren Long-hui, Zhang Rong-shu
(College of Landscape Architecture, Northeast Forestry University, Harbin 150040, China)
To enrich the microbial resources ofTrichodermaspp. for biocontrol, a dominantTrichodermastrain was isolated from the rhizosphere soil of a landscape plant, Sedum spectabile Boreau. The isolatedTrichodermastrain was identified using colonial-and micro-morphological observations, ribosomal DNA (rDNA)-internal transcribed spacer (ITS) sequence alignment, and homology analysis using a Neighbour-Joining method. An agar disk diffusion method was used to investigate the antagonism between the isolatedTrichodermastrain and three other soil-borne pathogenic fungi,Sclerotiniasclerotiorum,Rhizoctoniasolani, andAlternariaalternata. The tissue-cultured plantlets ofPopulusdavidiana×P.albavar.Pyramidaliswere further inoculated with the isolatedTrichodermastrainand the resistance of the inoculated seedling leaves againstA.alternatawas evaluated in vitro. The results demonstrated that the isolatedTrichodermastrain wasTrichodermalongibrachiatum and it exhibited evident and differential antagonistic effects against each of the three tested soil-borne pathogenic fungi. We designated the isolated Trichoderma strain as Tl-70. The antifungal activity of Tl-70 againstS.sclerotiorumwas a 77.71% inhibition, which was the highest and significantly higher than that againstR.solaniorA.alternata(P<0.05). The antifungal activity againstR.solaniwas a 58.56% inhibition, and significantly higher than that againstA.alternata(P<0.05, 53.32%), which was the lowest of the three tested strains. Moreover, inoculation with Tl-70 enhanced the disease resistance of the leaves of tissue-culturedPopulusdavidiana×P.albavar.Pyramidalisseedlings against the pathogenA.alternata. Our results suggest that Tl-70 is a potential biocontrol strain.
Trichoderma; morphology; ITS sequence analysis; Phylogenetic tree; antagonistic effect;Populusdavidiana×P.albavar.Pyramidalis
Zhang Rong-shu E-mail:zrs6504@sina.com
10.11829/j.issn.1001-0629.2016-0234常媛,杨兴堂,姜传英,姚志红,贾让,任龙辉,张荣沭.一株能拮抗3种土传病害病原真菌的长枝木霉.草业科学,2017,34(2):246-254.
2016-05-06接受日期:2016-11-17
2015年国家级本科生创新训练项目(201510225062);中央高校基本科研业务专项E类预研项目(2572015EY01);国家自然科学基金(31370642)
常媛(1993-),女,河南济源人,在读本科生,主要从事园林植物种质资源研究及植物病害生物防治。E-mail:630603391@qq.com
张荣沭(1965-),女,黑龙江哈尔滨人,副教授,博士,主要从事园林植物种质资源研究及植物病害生物防治。E-mail:zrs6504@sina.com
S432.4+4
A
1001-0629(2017)2-0246-09
Chang Y,Yang X T,Jiang C Y,Yao Z H,Jia R,Ren L H,Zhang R S.ATrichodermalongibrachiatumstrain with antagonistic effects against three soil-borne pathogenic fungi.Pratacultural Science,2017,34(2):246-254.
猜你喜欢
浙江农林大学学报(2022年5期)2022-10-12
甘肃教育(2021年10期)2021-11-02
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
食品与机械(2019年1期)2019-03-30
食品与机械(2019年1期)2019-03-30
天然产物研究与开发(2018年7期)2018-08-21
现代食品(2018年5期)2018-06-06
中国酿造(2017年11期)2017-12-06
化学与生物工程(2017年11期)2017-11-29
西藏大学学报(自然科学版)(2015年1期)2015-12-29
