
棉花S—腺苷甲硫氨酸合成酶基因的克隆及表达分析
2017-03-17 11:33宋成攀孟娟夏松波王孝刚张教海张
湖北农业科学 2016年23期
宋成攀+孟娟+夏松波+王孝刚+张教海+张友昌+别墅

摘要:利用iTRAQ技术从陆地棉(Gossypium hirsutum L.)HM-40中筛选并克隆到S-腺苷甲硫氨酸合成酶GhSAMS基因。GhSAMS基因的开放阅读框全长为1 182 bp,编码393个氨基酸,预测GhSAMS蛋白质含有15个磷酸化位点,推测其功能的行使可能与激酶磷酸化相关,进化分析表明S-腺苷甲硫氨酸合成酶与茶树(Camellia sinensis)的SAMS蛋白质相似性最高。qRT-PCR分析表明,在接种黄萎病菌VD07菌系后,GhSAMS表达量逐渐增加,随着接种时间的延长,其相对表达量随之增加,推测其在陆地棉抗黄萎病防卫反应中可能起重要作用。在嫁接棉株中,利用VIGS技术成功地沉默GhSAMS基因,然后对沉默棉株接种黄萎病菌VD07,鉴定其病情指数为62.5,抗性级别为感病;而转化空载体和未接种载体处理的嫁接棉株病情指数分别为30.0和31.2,抗性级别为耐病,表明GhSAMS基因沉默后嫁接棉对黄萎病的抗性丧失。推测GhSAMS基因在陆地棉抗黄萎病的过程中可能起重要作用。
关键词:棉花(Gossypium hirsutum L.)嫁接;S-腺苷甲硫氨酸合成酶基因;黄萎病;VIGS
中图分类号:S562 文献标识码:A 文章编号:0439-8114(2016)23-6261-06
DOI:10.14088/j.cnki.issn0439-8114.2016.23.065
Abstract:The GhSAMS gene was cloned from cotton(Gossypium hirsutum L.) cultivar HM-40, which was selected by iTRAQ of sea-land grafting cotton. The GhSAMS open reading frame(ORF) was 1 182 bp and encoded a protein of 393 amino acids. Many Phosphorylation sites were found in GhSAMS protein, which suggested that the function of GhSAMS is related to phosphorylation of these sites. The cloned gene was most closely related to Camellia sinensis CHS protein by phylogenetic analysis. Real-time quantitative reverse transcription polymerase chain reaction showed that GhSAMS gene was rapidly activated after inoculation with Verticillium dahliae. When time prolonged,the expression of related genes always inc reased. It was assumed that the GhSAMS gene might play an important role in the defense reaction against Verticillium wilt. Virus induced gene silencing was used to silence endogenous genes in resistant upland cotton by sea-land grafting cotton targeting a fragment of GhSAMS. VD07 was used to inoculate silenced cotton plants to identify disease resistance. The results showed that the disease indices of wild type vector control and sea-land grafting cotton plants were 30.0 and 31.2,respectively. The disease index of silenced GhSAMS plants was 62.5. These results confirmed that the GhSAMS gene might relate to Verticillium wilt resistance in cotton.
Key words:cotton(Gossypium hirsutum L.) grafting;GhSAMS gene;Verticillium wilt;VIGS
S-腺苷甲硫氨酸合成酶(S-adenosylmethionine synthetase,SAMS)是生物合成乙烯和多胺等物质的关键酶[1],主要参与了植物代谢中转甲基、转硫和转氨丙基等多种生化反应,并且能催化L-甲硫氨酸和ATP合成S-腺苷甲硫氨酸[2,3]。因此,对S-腺苷甲硫氨酸合成酶基因的研究在植物抗逆反应,尤其是抗病虫害等方面的研究具有重要的意义。陆地棉(Gossypium hirsutum L.)的生产长期遭受黄萎病(Verticillium wilt)的侵袭[4],目前为了进一步提高棉花对黄萎病的抗性,棉花黃萎病抗病性的分子机理研究倍受重视。
目前,陆地棉的抗黄萎病研究主要有四个方面,生理生化特性、克隆抗病相关基因、鉴定抗病相关蛋白和表观遗传学。通过对不同嫁接方式的棉花接种黄萎病后进行生理生化特性的研究,发现嫁接能够提升棉花的黄萎病抗病性[5]。利用基因步移等方法克隆出了GhSKIP35基因,推测其参与了棉花早期抗病信号的传递途径[6]。采用RNA-seq测序技术鉴定出Dirigent-like蛋白在不同抗性棉花品种中的表达变化,发现其与黄萎病菌侵入后棉花植株的抗性反应相关[7]。本研究利用iTraQ技术筛选在抗黄萎病过程中可能起作用的蛋白质,然后克隆得到其相应基因完整的ORF区序列,对其进行相关生物信息学分析和接种黄萎病菌后的表达分析,另外利用VIGS技术沉默这个基因,然后调查了沉默棉花的抗病指数,以期为更深入地研究其功能提供参考。
1 材料和方法
1.1 试验材料及处理
陆地棉感病品种HM-40,海岛棉高抗品种海7124,由武汉大学生命科学院实验室鉴定保存,于两片1元硬币大小的真叶时期嫁接处理,海岛棉为砧木,陆地棉为接穗。嫁接成活后7 d接强力致病菌VD07(中国农业科学院棉花研究所朱荷琴提供),每株接菌20 mL,浓度为107个/mL,分别取0、2、4、6、8、10 d的棉花真叶,液氮冷冻后保存于-80 ℃冰箱待用。
VIGS研究在嫁接成活后7 d 进行,方法参照Gao等[8]转化棉花的技术方法,分别注射接种农杆菌pYL-156-pYL-192和pYL-156-GhSAMS-pYL-192,在16 d后取嫁接棉真叶,液氮冷冻后保存于 -80 ℃冰箱待用。在农杆菌转化16 d后接大丽轮枝菌VD07,每株接菌20 mL,浓度为107个/mL,并在接种25 d后调查病情指数。
DP441植物RNA快速提取试剂盒购自北京天根生物科技有限公司;PrimeScriptTM RT reagent Kit with gDNA Eraser反转录试剂盒和pMD18-T载体购自TAKARA(大连)生物科技有限公司;OMEGA E.Z.N.A.?誖Cycle Pure Kit纯化试剂盒、EsTaq mix、大肠杆菌感受态top 10购自武汉欧瑞文生物科技有限公司;kod plus neo高保真聚合酶购自Toyobo(上海)生物科技有限公司;GV3101农杆菌感受态实验室自制,烟草脆裂病毒沉默载体pYL-192、pYL-156由清华大学刘玉乐教授馈赠。
1.2 总RNA提取及cDNA制备
采用天根DP441试剂盒提取总RNA,然后用TAKARA PrimeScriptTM RT reagent Kit with gDNA Eraser试剂盒将RNA反转录成第一链cDNA。
1.3 GhSAMS基因的克隆
采用Oligo 7软件设计引物。以GhSAMS-F(5′AAAACAGCAGAGAAATTAAGGA 3′)和GhSAMS-R(5′ GAAGAAGCTCCAATTAAGCA 3′)为引物,用kod plus neo进行基因扩增。PCR程序为94 ℃预变性,5 min;98 ℃变性15 s,55 ℃退火30 s,68 ℃延伸1 min,30个循环;继续延伸68 ℃,10 min,4 ℃结束反应。扩增产物于1%的琼脂糖水平凝胶中电泳检测。PCR产物纯化使用Omega PCR产物纯化试剂盒,严格按照说明进行操作。然后采用pMD18-T连接,转化大肠杆菌感受态细胞top 10,倒置培养过夜。挑取单克隆,进行菌液PCR检测,并且测序进行验证。测序由苏州金唯智科技有限公司完成。
1.4 GhSAMS基因的VIGS载体的构建
设计VIGS引物,以pYL-156-SAMS-F(5′ CCGGAATTCAAAACAGCAGAGAAATTAAGGA 3′)和pYL-156-SAMS-R(5′CCGCTCGAGGAAGAAG- CTCCAATTAAGCA 3′)为引物(下划线部分为保护碱基和酶切位点),用LA Taq进行基因扩增。PCR程序为94 ℃预变性5 min;94 ℃变性30 s,55 ℃退火30 s,72 ℃延伸1.5 min,30个循环;后延伸72 ℃,10 min,4 ℃结束反应。扩增产物于1%的琼脂糖水平凝胶中电泳检测。PCR产物纯化使用Omega PCR产物纯化试剂盒,严格按照说明进行操作。然后进行酶切反应,同样酶切pYL-156质粒,solutionⅠ连接4 h,转化大肠杆菌感受态细胞top 10,倒置培养过夜。挑取单克隆,进行菌液PCR检测,并且测序进行验证。测序由苏州金唯智科技有限公司完成。
提取阳性质粒,用农杆菌感受态细胞GV3101电转,涂平板后28 ℃倒置暗培养过夜,挑取单克隆,进行菌液PCR检测。挑取阳性的农杆菌克隆,分别为pYL-156-GhSAMS[8]。
1.5 克隆序列的生物信息学分析
使用在线软件Protparam和PSORT分析这两个蛋白质的理化性质并进行亚细胞定位的预测。采用SignalP 4.1分析蛋白质是否含有信号肽。采用TMHMM预测跨膜螺旋。然后在NCBI上的Protein blast进行氨基酸同源序列比对,用DANMAN进行氨基酸同源性分析,用MEGA 5软件对其他植物GhSAMS蛋白构建系统进化树。应用NetPhos 2.0程序预测磷酸化位点[9]。
1.6 GhSAMS基因表达模式分析
按照PrimeScriptTM RT reagent Kit with gDNA Eraser反转录试剂盒的说明书,采用qRT-PCR 分析GhSAMS基因的相对表达量。扩增目标基因引物为qSAMS-F(5′TGATACTTATGGAGGATGGG3′) 和qSAMS-R(5′ATGGCGTAGGAGACTTGC3′);内参基因引物为actin-F(5′TCACGGAAGCACCTCTCAAC 3′)和actin-R(5′ACAAAGAGAGAACGGCCTGG 3′),每个样品3次技术重复和3次生物学重复,使用Bio-Rad CFX96 Manager荧光定量仪进行分析,按照2^-ΔΔCt法计算相对表达量。qRT-PCR程序:95 ℃预变性30 s;95 ℃变性5 s,57 ℃退火延伸20 s,40个循环;65~95 ℃,每步增加0.5 ℃,5 s做溶解曲线以确定扩增产物的特异性,延伸5 s。
1.7 GhSAMS基因的功能半定量和qRT-PCR 分析
按照PrimeScriptTM RT reagent Kit with gDNA Eraser反轉录试剂盒的说明书,将反转录的cDNA原液稀释10倍,采用引物V-qSAMS-F(5′ AGCCATTGTCTGTATTTG 3′)和V-qSAMS-R(5′ CTCATTGCTATTTCATTC 3′),内参基因引物为actinB-F(5′ GTTATGGTTGGGATGGG 3′)和actinB-R(5′ CAAGGTCAAGACGGAGG 3′)进行半定量分析,PCR程序为94 ℃预变性,5 min;94 ℃变性30 s,55 ℃退火30 s,72 ℃延伸20 s,分别进行20、25、28、31、32、33、34、35、36、37、38个循环;延伸72 ℃,10 min,4 ℃结束反应。扩增产物于1%的琼脂糖水平凝胶中电泳检测。
采用qRT-PCR 分析GhSAMS基因的相对表达量。扩增目标基因引物为qvGhSAMS-F:(5′ GTTTATCTTATCTGCTGTC 3′)和qvGhSAMS-R(5′ CTC- ATTGCTATTTCATTC 3′);内参基因引物为actin-F(5′ TCACGGAAGCACCTCTCAAC 3′)和actin-R(5′ ACAAAGAGAGAACGGCCTGG 3′),qPCR方法同上。
2 结果与分析
2.1 陆地棉基因GhSAMS的克隆
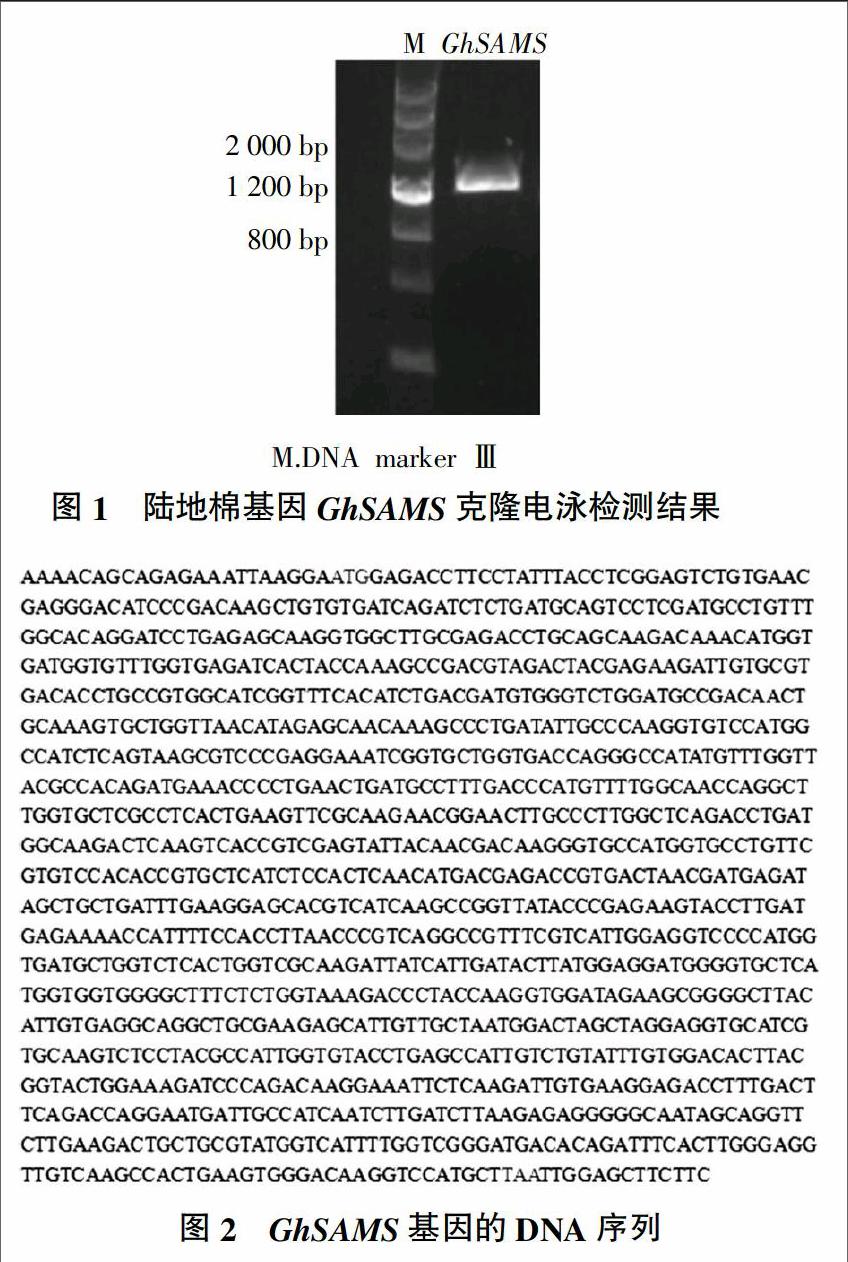
以反转录得到的cDNA为模板,以GhSAMS-F和GhSAMS-R为引物,获得了1 217 bp的阳性条带(图1),测序结果见图2。由图2可知,ORF全长为1 182 bp。
2.2 GhSAMS蛋白质的理化性质
将GhSAMS蛋白质的氨基酸序列输入分析软件Protparam,该蛋白质的分子式为C1 903H3 001N525O583 S15,其等电点约为5.65,分子量43.044 kD,氨基酸数目结果见表1,使用频率较高的氨基酸有Lys、Val、Ala、Gly、Thr、Asp,呈负电荷的酸性氨基酸(Glu和Asp)共56个,呈正电荷的碱性氨基酸(Lys和Arg)共45个,表明该蛋白质带负电且呈酸性。预测其不稳定指数为21.76,在体外的哺乳动物网织红细胞中表达时半衰期约30 h,在酵母中表达时半衰期超过20 h,在大肠杆菌中表达时半衰期超过10 h,蛋白质表现稳定。
2.3 GhSAMS蛋白质的亲疏水性和亚细胞定位预测
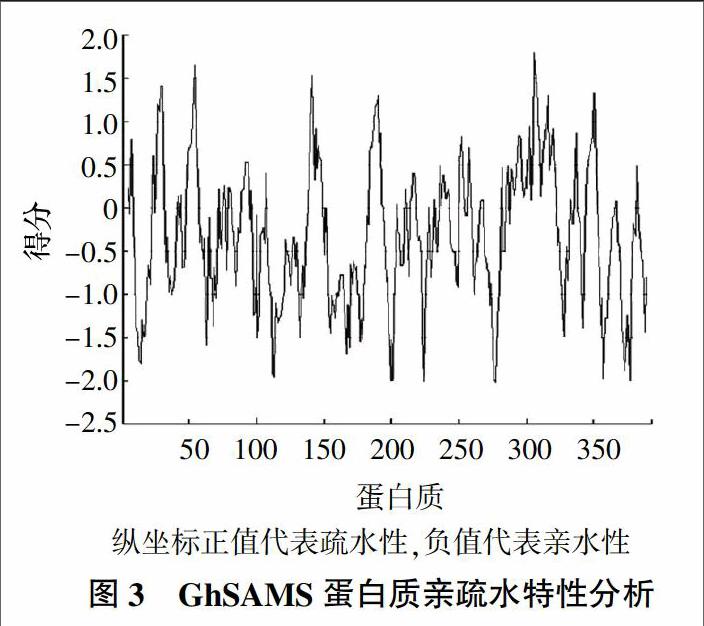
使用蛋白质的亲疏水性预测在线软件ProtScale(http://web.expasy.org/protscale/)将GhSAMS蛋白质的氨基酸序列输入,分析结果见图3。GhSAMS蛋白质的亲水性平均系数(GRAVY)是-0.300,表明整个蛋白质呈亲水性。再将GhSAMS蛋白质的氨基酸序列输入到在线分析软件PSORT中进行亚细胞定位预测,结果显示GhCHS蛋白质可能定位于细胞质(得分0.450)、过氧化物酶体(得分0.437)、溶酶体(得分0.100)和线粒体基质(得分0.100);使用SignalP 4.1分析GhSAMS蛋白质有无信号肽序列,结果显示GhSAMS蛋白质无信号肽序列,那么该蛋白质无法进行跨膜运输;使用TMHMM预测该蛋白质的跨膜结构域,结果显示该蛋白质没有跨膜螺旋区。根据以上的预测,推断GhSAMS蛋白质是一种亲水性细胞质蛋白质。
2.4 GhSAMS蛋白质的同源比对和系统进化树分析
用BlastP进行同源性分析,发现陆地棉 GhSAMS的氨基酸序列与其他物种的该蛋白质具有很高的同源性。选取10个代表性物种,如木豆(Cajanus cajan,AEY85025.1)、茶树(Camellia sinensis,AHB32111.1)、棕榈(Elaeis guineensis,XP_010921340.1)、野生大豆(Glycine soja,KHN28175.1)、番薯(Ipomoea batatas,ABP35525.1)、苜蓿(Medicago sativa,AAT40304.1)、桃树(Prunus persica,AGF95108.1)、白梨X(Pyrus X bretschneideri,AJF23500.1)、刺槐(Robinia pseudoacacia,AIT39705.1)和杯萼海桑(Sonneratia alba,AGJ71754.1),采用DNAMAN軟件进行同源序列比对,结果如图4所示,氨基酸序列具有很高的相似性。使用MEGA 5.0软件建构蛋白质系统进化树,结果如图5所示,陆地棉的GhSAMS蛋白质与茶树的AHB32111.1进化关系最近。
2.5 GhSAMS蛋白质的磷酸化位点预测
磷酸化是蛋白质修饰的一种方式,参与并调控着许多细胞活动,在真核生命活动中有着重要意义,蛋白质磷酸化修饰后,改变了蛋白质的构象、稳定性、活性以及在细胞中所处的位置[10]。GhSAMS蛋白质含有30个苏氨酸和16个丝氨酸,潜在的磷酸化位点较多。运用蛋白质磷酸化位点预测在线软件NetPhos 2.0进行分析,预测结果见图6。GhCHS蛋白质潜在苏氨酸(Thr)磷酸化位点有6个,潜在丝氨酸(Ser)磷酸化位点有6个,潜在酪氨酸(Tyr)磷酸化位点有3个。预测GhSAMS蛋白质磷酸化位点共15个,GhSAMS蛋白质的功能与激酶磷酸化相关。
2.6 GhSAMS基因在黄萎病菌胁迫下的相对表达量
利用qRT-PCR方法分析GhSAMS基因在嫁接棉(海7124为砧木,HM-40为接穗)真叶中的相对表达量。结果显示(图7),GhSAMS基因在接种大丽轮枝菌VDO7菌株初期,基因表达量增加,随着接菌处理时间的延长,相对表达量逐渐增加,且2、4、6、8、10 d与未接菌时相比差异显著(P<0.05),在接种10 d时,该基因的表达量上调了约28倍,推测其在嫁接棉抗黄萎病防卫反应中可能起重要作用。
2.7 利用VIGS技术研究GhSAMS基因的功能
在接种农杆菌16 d后分别取接种空载体pYL-156-pYL-192和pYL-156-GhSAMS-pYL-192的棉花叶样品提取总RNA,进行RT-PCR分析,然后进行半定量试验(图8)和qRT-PCR试验,发现接种带目的基因载体的比接种带空载体的基因表达量分别下降91.85%(图9)。对未接农杆菌处理嫁接棉、转化空载体嫁接棉和基因沉默嫁接棉接种大丽轮枝菌后25 d进行调查,结果表明,未接农杆菌嫁接棉病情指数为31.2,表现为耐病;转化空载体嫁接棉病情指数为30.0,表现为耐病;基因沉默嫁接棉病情指数分别为62.5,表现为感病(图10)。
3 讨论
20世纪生命科学领域持续飞速发展,越来越多新的技术被开发出来,并且与计算机学科交叉结合起来产生了生物信息学这门全新的学科,科研工作者不断有新的发现和重大突破。在植物基因工程研究方面,大量的基因被克隆出来,进而深入的研究其功能,尤其是VIGS(Virus induced gene silencing)技术的开发与应用,在研究植物基因功能时,不再必须获得RNAi的转基因纯合植株,通过VIGS技术即可快速沉默相关基因,无疑加速了研究基因的功能[11]。S-腺苷甲硫氨酸合成酶是植物合成乙烯和多胺等物质过程中的关键酶,还能合成S-腺苷甲硫氨酸,植物代谢过程中许多生长发育、抗逆反应相关的次生代谢物质的生成均与这些途径密不可分,大量研究证实他与乙烯和多胺的合成相关,这些物质都与植物的抗逆防御反应有直接关系[1,12,13]。虽然对棉花抗黄萎病相关基因的研究较多,但是目前关于S-腺苷甲硫氨酸合成酶在棉花抗黄萎病的研究上还较少,值得深入研究,也能进一步了解这类基因在植物细胞中参与的过程与作用。
本研究克隆到陆地棉的GhSAMS基因的ORF区后,对其进行了生物信息学分析预测,预测GhSAMS蛋白质是一种亲水性的细胞蛋白质,另外预测了GhSAMS蛋白质含有一些磷酸化位点,因而推测其功能的行使与激酶磷酸化可能相关,并且发现这个基因受黄萎病菌胁迫诱导表达,推测其可能与植物抗病防御反应有关。成功构建了GhSAMS基因的病毒沉默载体,也转化到嫁接的棉花植株,发现沉默该基因使其丧失功能后,会导致棉花抗黄萎病抗性的丧失,进一步验证了这个基因的某些生理功能,即可能参与了棉花的抗病防御反应,为更深入研究其基因功能以及棉花抗黄萎病的相关途径提供了理论支持。
参考文献:
[1] NAIR R B,BASTRESS K L,RUEGGER M O,et al. The Arabidopsis thaliana REDUCED EPIDERMAL FLUORESCENCEI gene encodes an aldehyde dehydrogenase involved in ferulic acid and sinapic acid biosynthesis[J].The Plant Cell,2004, 16(2):544-554.
[2] MATO J,ALVAREZ L,ORTIZ P,et al. S-adenosylmethionine synthesis:Molecular mechanisms and clinical implications[J].Pharmacol Therapeutics,1997,73:265-280.
[3] BELBAHRI L,CHEVALIER L,BENSADDEK L,et al.Differentexpression of an S-adenosylmethionine synthetase gene in transgenic tobacco callus modifies alkaloid biosynthesis[J].Biotechnol Bioeng,2000,69:11-20.
[4] 简桂良,邹亚飞,马 存.棉花黄萎病连年流行的原因及对策[J].中国棉花,2003,30(3):13-14.
[5] 陈营营,别 墅.海陆嫁接棉苗对黄萎病抗性及相关生理指标研究[J].分子植物育种,2014,12(6):1148-1158.
[6] 刘 凯,简桂良.棉花GhSKIP35基因的克隆与表达分析[J].棉花学报,2015,27(2):111-117.
[7] 郭宝生,师恭曜,华金平,等.黄萎病菌侵染下陆地棉dirigent-like蛋白基因表达差异分析[J].中国农业科学,2014(22):4349-4359.
[8] GAO X Q,ROBERT C,BRITT,et al.Agrobacterium-mediated virus-induced gene silencing assay in cotton[J].Journal of Visualized Experiments,2011,54:e2938.
[9] 張天豹,阴祖军,叶武威,等.陆地棉线粒体耐盐基因ccm C的克隆与表达分析[J].分子植物育种,2015(7):1502-1508.
[10] 方智振,周丹蓉,姜翠翠,等.李Ran基因的克隆与生物信息学分析[J].分子植物育种,2014,12(4):780-787.
[11] ROBERTSON D. VIGS vectors for gene silencing: Many targets,many tools[J].Annu Rev Plant Biol,2004,55:495-519.
[12] CHEN X J,LIU G S.A review of the pathway of ethylene biosynthesis and the relevant genetic engineering[J].Journal of Tropical and Subtropical Botany,2002,10(1):83-98.
[13] HEBY O,PERSSON L.Molecular genetics of polyamine synthesis in eukaryotic cells[J].Trends Biochem Sci,1990,15:153-158.
猜你喜欢
食品科学(2023年13期)2023-08-12
中国生物化学与分子生物学报(2022年7期)2022-09-07
中国药科大学学报(2021年6期)2021-12-31
天津医科大学学报(2019年6期)2019-08-13
分析化学(2017年12期)2017-12-25
中国食用菌(2016年4期)2016-01-25
分析测试学报(2015年9期)2015-12-17
安徽医科大学学报(2015年9期)2015-12-16
遗传(2014年3期)2014-02-28
新疆农垦科技(2014年7期)2014-02-28
