
干旱胁迫对草莓苗期叶片光合特性的影响
2017-03-17 00:07张庆华曾祥国向发云韩永超陈丰滢
湖北农业科学 2016年23期
张庆华+曾祥国+向发云+韩永超+陈丰滢+过聪+韩玉萍+顾玉成

摘要:采用盆栽控水方法研究了干旱胁迫对5个草莓品种苗期叶绿素含量和光合特性的影响。结果表明,在干旱胁迫下,草莓叶片的叶绿素含量(SPAD)、净光合速率、蒸腾速率、气孔导度、胞间CO2浓度均下降,且甜查理的下降幅度最小,艳丽的下降幅度最大。在轻度干旱胁迫下,5个草莓品种的净光合速率和胞间CO2浓度均与对照存在显著差异(P<0.05);叶绿素含量、蒸腾速率和气孔导度与对照无显著差异(P>0.05)。在重度干旱胁迫下,5个草莓品种的净光合速率、蒸腾速率和气孔导度均与对照存在显著差异(P<0.05);胞间CO2浓度与对照无显著差异(P>0.05);艳丽和宁丰的叶绿素含量与对照相比显著下降(P<0.05)。以上结果表明,在轻度干旱胁迫下,草莓主要是通过降低净光合速率和胞间CO2浓度响应干旱胁迫,气孔因素影响光合作用;在重度干旱胁迫下,草莓主要是通过降低净光合速率、蒸腾速率和气孔导度响应干旱胁迫,非气孔因素影响光合作用。不同的草莓品种光合作用对干旱的响应能力不同。其中,甜查理具有较强的抗旱性,艳丽的抗旱性较差。
关键词:草莓;干旱胁迫;叶绿素含量;光合特性
中图分类号:S668.4 文献标识码:A 文章编号:0439-8114(2016)23-6147-04
DOI:10.14088/j.cnki.issn0439-8114.2016.23.036
Abstract: Chlorophyll content and photosynthetic characteristics of five strawberry varieties in seedling stage were studied under different levels of drought treatments. The results showed that the chlorophyll content(SPAD),net photosynthetic rate (Pn),transpiration rate (Tr),stomatal conductance (Gs) and intercellular CO2 concentration (Ci) were decline under drought stress,and the decline rate of these indexes in Sweet Charlie were minimum,but in Yanli were maximum. Under mild drought stress, compared with the control, the differences of Pn and Ci in different varieties were statistically significantly(P<0.05),but the differences of chlorophyll content,Tr,and Gs were no significantly(P>0.05). Under severe drought stress,compared with the control, the differences of Pn,Tr and Gs in different varieties were significantly(P<0.05),but the difference of Ci was no significantly(P>0.05);the chlorophyll content of Ningfeng and Yanli were significantly decline(P<0.05). In concluded,Pn and Ci were decline in response to mild drought stress,and stomatal factors affected photosynthesis. Nevertheless,Pn,Tr and Gs were decline in response to severe drought stress,and non-stomatal factors affected photosynthesis. Meanwhile,different varieties of strawberry had different capabilities to repose to drought stress,for example,Sweet Charlie had stronger drought resistance and Yanli had weaker drought stress resistance in five strawberry varieties.
Key words: strawberry; drought stress; chlorophyll content; photosynthetic characteristics
草莓為多年生温带草本果树,是中国广泛栽培的重要经济作物之一[1]。但由于其根系分布浅,叶面积较大,更易受到干旱胁迫的影响[2]。干旱胁迫会对植物的生长、代谢等多方面产生影响, 其中对光合作用的影响尤为突出和重要。因此,干旱胁迫下,植物光合作用的强弱可作为鉴定其抗旱能力强弱的指标之一[3]。植物叶片气体交换参数和光合色素含量是光合作用的基础,通过对叶片光合气体交换参数和光合色素含量的分析,可以了解植物叶片对光能的吸收和利用[4,5]。目前关于草莓干旱胁迫下光合特性的研究较少[2]。本研究以5个草莓品种为材料,研究了干旱胁迫对不同草莓品种光合特性和光合色素的影响,探讨了不同品种对干旱胁迫的适应性反应,旨在为改进草莓抗旱栽培技术措施和选育优良抗旱品种提供理论依据。
1 材料与方法
1.1 试验材料
供试草莓品种为艳丽、宁丰、甜查理、晶瑶、晶玉。
1.2 试验方法
1.2.1 试验处理 试验在湖北省农业科学院经济作物研究所草莓实验基地进行,选用4~5叶1心的5种草莓品种的组培移栽苗,栽于盆钵,盆土为育苗用土,土壤类型为沙壤土。待所有材料经过充分的缓苗后,再进行干旱处理。土壤含水量分为3个等级:30%~40%土壤含水量为重度干旱胁迫,50%~60%土壤含水量为轻度干旱胁迫,70%~80%土壤含水量为正常灌水(CK),每个处理10盆。采用土壤称重法控制土壤含水量,每天上午8:30称取盆重,并补充水分,使各处理保持设定的土壤含水量。干旱处理10 d后,测定相关光合生理参数。
1.2.2 试验测定方法 干旱处理后第11天在8:30~10:30,选择叶位、叶龄、受光部位相对一致的成熟草莓叶片作为测定对象,净光合速率、蒸腾速率、胞间CO2浓度和气孔导度等光合生理参数用美国Li-COR公司生产的LI-6400型光合作用仪测定[6],测定时光照强度为800 μmol/(m2·s),叶室温度为25 ℃,大气CO2浓度约为400 μL/L,每个处理重复测定3次。叶绿素含量采用日本SPAD-502 Plus叶绿素仪进行测定[7-9],测定时选取无叶脉的部位,每片叶子至少选择8个点进行测量,并求其平均值,每个处理重复测定3次。
1.3 试验数据分析
数据采用Excel 2016整理和分析,用分析软件SPSS19.0进行单因素方差分析和多重比较。
2 结果与分析
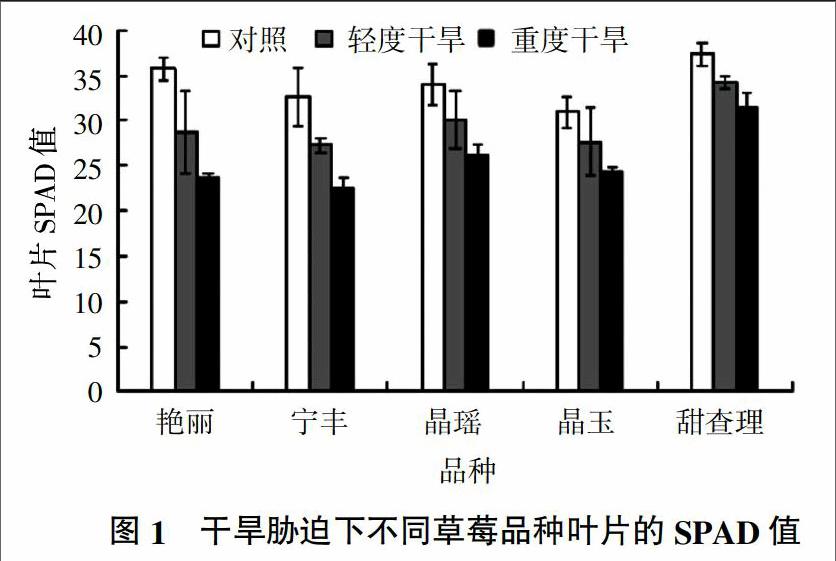
2.1 干旱胁迫对不同草莓品种叶绿素含量的影响
叶绿素是反映光合作用强弱的重要指标,其含量与植物的生长状况和叶片的光合能力密切相关[10,11]。叶绿素的相对含量可用SPAD-502 Plus叶绿素仪测得SPAD值(叶色值,Leaf color values)来衡量[8]。由图1可以看出,5个草莓品种在干旱胁迫下叶绿素含量有明显的变化。随着干旱胁迫的加重,叶绿素含量不断下降,其中,甜查理的叶绿素含量下降幅度最小,艳丽的下降幅度最大。与对照相比,在轻度干旱胁迫下,5个草莓品种叶绿素含量的下降幅度分别为艳丽18.83%、宁玉16.63%、晶瑶11.57%、晶玉10.56%、甜查理8.56%;在重度干旱胁迫下,5个草莓品种叶绿素含量下降幅度分别为艳丽32.58%,宁玉30.41%,晶瑶22.94%,晶玉20.58%,甜查理15.78%。方差分析结果显示,轻度干旱胁迫下5个草莓品种的叶绿素含量均与对照无显著差异(P>0.05);重度干旱胁迫下艳丽和宁丰与对照有显著差异(P<0.05),其他品种无显著差异(P>0.05)。说明不同草莓品种叶绿素对干旱胁迫的响应有显著差异。
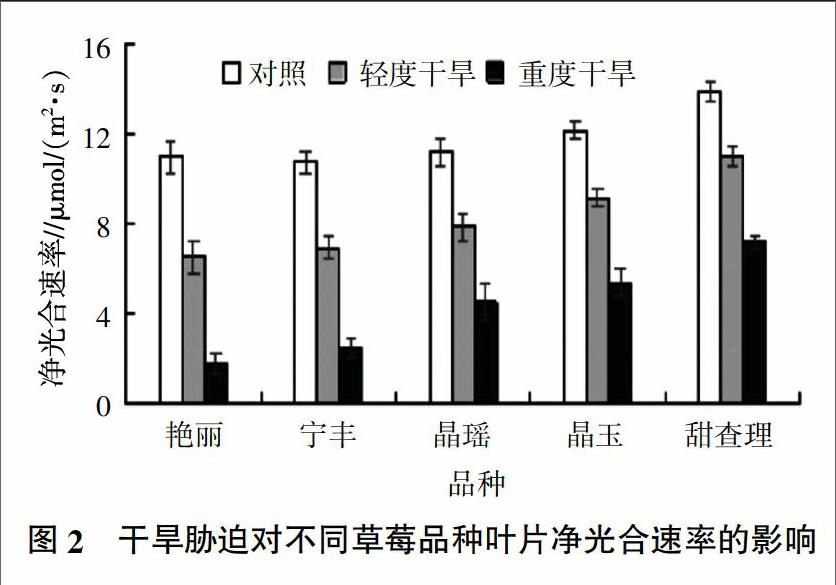
2.2 干旱胁迫对不同草莓品种净光合速率的影响
从图2可以看出,草莓在受到干旱胁迫后净光合速率下降,且随着胁迫的加重,净光合速率下降幅度越大,其中,甜查理的净光合速率下降幅度最小,艳丽的最大。与对照相比,在轻度干旱胁迫下,草莓品种净光合速率快速下降,下降幅度达到20%~40%,说明在轻度干旱胁迫下,草莓净光合速率已受到较大影响;在重度干旱胁迫下,草莓净光合速率急剧下降,5个草莓品种净光合速率下降幅度分别为艳丽83.63%、宁玉77.10%、晶瑶59.71%、晶玉55.78%、甜查理48.24%。方差分析結果显示,轻度和重度干旱胁迫下5个草莓品种的净光合速率均与对照存在显著差异(P<0.05)。说明草莓品种净光合速率对干旱胁迫的响应很敏感,且不同草莓品种对干旱的响应能力不同。
2.3 干旱胁迫对不同草莓品种蒸腾速率的影响
从图3可以看出,草莓在受到干旱胁迫后蒸腾速率下降,且随着干旱胁迫的加重,蒸腾速率逐渐降低。在轻度干旱胁迫下,草莓品种的蒸腾速率缓慢下降,方差分析结果显示,5个草莓品种的蒸腾速率均与对照无显著差异(P>0.05)。在重度干旱胁迫下,草莓蒸腾速率下降幅度达到50%左右,与对照相比,5个草莓品种蒸腾速率下降幅度分别为艳丽58.32%、宁玉56.09%、晶瑶50.26%、晶玉46.12%、甜查理43.05%,方差分析结果显示,5个草莓品种的蒸腾速率均与对照存在显著差异(P<0.05)。说明轻度干旱胁迫对草莓蒸腾速率的影响不大,但是重度干旱胁迫会导致草莓水分不足,进而减缓蒸腾作用以减轻伤害。
2.4 干旱胁迫对不同草莓品种气孔导度的影响
从图4可以看出,草莓在受到干旱胁迫后气孔导度随着胁迫的加重逐渐减小。在轻度干旱胁迫下,草莓气孔导度缓慢下降;方差分析结果显示,轻度干旱胁迫下5个草莓品种的气孔导度均与对照无显著差异(P>0.05)。在重度干旱胁迫下,草莓气孔导度快速下降,其中,艳丽气孔导度下降幅度最大,为71.63%,其他草莓品种分别为宁玉63.31%、晶瑶56.49%、晶玉47.26%、甜查理46.29%;方差分析结果显示,重度干旱胁迫下5个草莓品种的气孔导度均与对照存在显著差异(P<0.05)。说明草莓气孔导度受到重度干旱胁迫的影响较大,且不同草莓品种气孔导度对干旱胁迫的响应能力不同。
2.5 干旱胁迫对不同草莓品种胞间CO2浓度的影响
从图5可以看出,草莓在受到干旱胁迫后胞间CO2浓度随着胁迫的加重先降低再增加。在轻度干旱胁迫下,草莓胞间CO2浓度急剧下降,5个草莓品种的胞间CO2浓度下降幅度分别为艳丽38.47%、宁玉39.89%、晶瑶32.32%、晶玉22.48%、甜查理24.59%。方差分析结果显示,轻度干旱胁迫下5个草莓品种的胞间CO2浓度均与对照存在显著差异(P<0.05),说明草莓胞间CO2浓度在轻度干旱胁迫下已受到显著影响,这可能与气孔导度的下降有关,干旱会引起气孔的关闭,使气孔导度下降,从而引起CO2的吸收减少,导致胞间CO2浓度下降。在重度干旱胁迫下,5个草莓品种的胞间CO2浓度较轻度干旱胁迫下有很大的回升,与对照相比,甜查理胞间CO2浓度下降最小,仅为7.24%,艳丽的最大为15.05%。方差分析结果显示,重度干旱胁迫下5个草莓品种的胞间CO2浓度均与对照无显著差异(P>0.05)。说明草莓胞间CO2浓度受到轻度干旱胁迫的影响较大,且草莓胞间CO2浓度在轻度和重度干旱胁迫下对光合作用的影响机制不同。不同草莓品种胞间CO2浓度下降幅度也显示其对干旱胁迫的响应能力不同。
3 讨论
光合作用是植物生命活动中的重要合成代谢过程之一,会直接影响植株的生长、发育状况,而干旱胁迫明显影响植物的光合作用[12,13]。植物通过叶绿素进行光合作用,叶绿素的合成与分解平衡会影响光合作用的進行。因此叶绿素的含量变化情况可以用来反映植物受干旱胁迫的程度[14,15]。本试验结果显示,在干旱胁迫下,5个草莓品种的叶绿素含量均下降,且随着干旱胁迫的加剧叶绿素含量的下降幅度增大。可能是由于干旱胁迫导致蛋白质合成受阻或破坏了叶绿体的结构,使叶绿素的合成减少,导致叶绿素含量下降[15,16]。其中甜查理的叶绿素含量降幅最小,说明甜查理具有较强的水分亏缺自身调节能力,能有效地防止或减少水分亏缺对光合作用的破坏,具有较强的抗旱性;艳丽的叶绿素含量下降幅度最大,说明其不能维持一定水平的光合作用,抗旱性较差。
干旱胁迫会直接影响植物的光合特性,会造成植物叶片的气体交换参数降低,这是植物对干旱胁迫的一种生理反应[13]。本试验结果表明,在干旱胁迫下,5个草莓品种叶片的净光合速率、气孔导度、胞间CO2浓度、蒸腾速率均下降,这是草莓对干旱胁迫的一种生理反应,是通过降低光合特性来缓解或降低受到的干旱胁迫伤害。这与徐金娥等[2]的研究结果是一致的。通常认为影响植物光合作用的因素分为气孔因素和非气孔因素。如果净光合速率随着胞间CO2浓度的降低而减少,那么气孔限制就是造成净光合速率下降的主要因素;净光合速率下降的同时将伴随着胞间CO2浓度的上升,那么非气孔限制就是造成净光合速率下降的主要因素[17-19]。本研究中,5个草莓品种在轻度干旱胁迫下,草莓叶片净光合速率和胞间CO2浓度随着干旱胁迫的下降而下降,说明草莓叶片净光合速率的下降是气孔限制造成的。在轻度到重度干旱胁迫下,草莓叶片净光合速率随着干旱胁迫的下降而下降,胞间CO2浓度随着干旱胁迫的下降而上升,说明草莓叶片净光合速率的下降是非气孔限制造成的。
在干旱胁迫下,5个草莓品种中甜查理能较好地通过自身来响应干旱胁迫,叶绿素含量的变化幅度不大,且净光合速率、气孔导度、蒸腾速率、胞间CO2浓度在干旱胁迫下变化幅度最小,而艳丽各参数变化幅度最大。说明甜查理表现出优良的抗旱性,艳丽抗旱性较差。
参考文献:
[1] 王玉坤,张 放,祝庭耀.国内草莓生产现状与发展趋势[J].北方园艺,2003(6):6-7.
[2] 徐金娥,陈 静.干旱胁迫对草莓光合作用的影响[J].保定学院学报,2008,21(2):35-37.
[3] 古丽江·许库尔汗,阿依古丽·铁木儿,孙雅丽,等.不同蓝靛果忍冬品种对干旱胁迫的光合生理响应比较分析[J].核农学报,2015,29(9):1833-1843.
[4] 眭晓蕾,毛胜利,王立浩,等.弱光条件下辣椒幼苗叶片的气体交换和叶绿素荧光特性[J].园艺学报,2007,34(3):615-622.
[5] MAXWELL K,JOHNSON G N. Chlorophyll fluorescence-A practical guide[J].J Exp Bot,2000,51(345):659-688.
[6] 卢琼琼,宋新山,严登华.干旱胁迫对大豆苗期光合生理特性的影响[J].中国农学通报,2012,28(9):42-47.
[7] 张云海,何念鹏,张光明,等.氮沉降强度和频率对羊草叶绿素含量的影响[J].生态学报,2013,33(21):6786-6794.
[8] QIU Z Y,WANG L H,ZHOU Q. Effects of bisphenol A on growth,photosynthesis and chlorophyll fluorescence in above-ground organs of soybean seedlings[J].Chemosphere,2013,90(3):1274-1280.
[9] 安玉艳,梁宗锁.植物应对干旱胁迫的阶段性策略[J].应用生态学报,2012,23(10):2907-2915.
[10] 党晓宏,高 永,虞 毅,等.3种滨藜属牧草苗期叶片解剖结构及生理特性对干旱的响应[J].西北植物学报,2014,34(5):976-987.
[11] FU X Y,MO W P,ZHANG J Y,et al. Shoot growth pattern and quantifying flush maturity with SPAD value in litchi (Litchi chinensis Sonn.)[J].Scientia Horticulturae,2014,174:29-35.
[12] 王玉珏,付秋实,郑 禾,等.干旱胁迫对黄瓜幼苗生长、光合生理及气孔特征的影响[J].中国农业大学学报,2010,15(5):12-18.
[13] LAFITTE H R,GUAN Y S,SHI Y,et al. Whole plant responses,key processes and adaptation to drought stress:The case of rice[J].J Exp Bot,2007,58(2):169-175.
[14] 王群英,胡昌浩.玉米不同叶位叶片叶绿体超微结构与光合性能的研究[J].植物学报,1988,30(2):146-150.
[15] 任安之,高玉葆,李 侠.内生真菌感染对黑麦草若干抗旱生理特征的影响[J].应用与环境生物学报,2002,8(5):535-539.
[16] CALOS G M,LORENZO L. Nitric oxide induces stomatal closure and enhances the adaptive plant response against drought stress[J].Plant Physiology,2001,126(3):1196-1204.
[17] FARQUHAR G D,SHARKEY T D. Stomatal conductance and photosynthesis[J].Annual Review of Plant Physiology,1982, 33(3):317-345.
[18] 郭卫华,李 波,黄永梅,等.不同程度的水分胁迫对中间锦鸡儿幼苗气体交换特征的影响[J].生态学报,2004,24(12):2716-2722.
[19] 张光灿,刘 霞,贺康宁,等.金矮生苹果叶片气体交换参数对土壤水分胁迫的响应[J].植物生态学报,2004,8(1):66-72.
猜你喜欢
山东农业科学(2016年12期)2017-01-21
山东农业科学(2016年12期)2017-01-21
山东农业科学(2016年11期)2016-12-17
现代园艺(2016年2期)2016-03-15
