
4b型单核细胞增生李斯特菌内化素基因NTSN_0462的功能初探
2017-03-16 06:38蔡雪薛孔苏伟谈卫军潘志明殷月兰焦新安
中国人兽共患病学报 2017年2期
赵 丹,姚 浩,蔡雪薛,孔苏伟,恽 茜,谈卫军,潘志明,殷月兰,焦新安
4b型单核细胞增生李斯特菌内化素基因NTSN_0462的功能初探
赵 丹,姚 浩,蔡雪薛,孔苏伟,恽 茜,谈卫军,潘志明,殷月兰,焦新安
目的 单核细胞增生性李斯特菌(Listeriamonocytogenes,Lm)对宿主的粘附、侵袭作用与其多种内化素蛋白密切相关。本研究以4b型菌株LmNTSN的內化素基因NTSN_0462为研究对象,初步探究其致病作用。方法 利用同源重组技术构建该基因缺失的突变株,研究0462基因对NTSN在生长、细胞侵袭和体内定植中的作用。人结肠腺癌细胞Caco-2、人肝癌细胞HepG2的侵袭率,以及BALB/c小鼠体内肝、脾脏中定植能力的差异。结果 缺失株的生长曲线结果显示,NTSN_0462基因对Lm在BHI培养基中的生长及代谢未造成明显影响。以人结肠腺癌细胞Caco-2、人肝癌细胞HepG2进行的体外试验表明,缺失株的侵袭能力低于野生株(P<0.001)。以BALB/c小鼠进行的体内试验显示,缺失株在肝脏中定殖的能力显著低于野生株(P<0.001),在脾脏中无统计学差异。结论NTSN_0462基因在LmNTSN入侵肝脏和定植中发挥重要作用,是侵袭相关的重要毒力因子。
单核细胞增生李斯特菌;NTSN_0462;血清型4b;内化素
单核细胞增生性李斯特菌是一种兼性胞内寄生菌,其感染的宿主范围较广,可引发人和多种动物的李斯特菌病。作为食源性病原菌,Lm主要通过污染的食物进入肠道继而引发宿主的感染,感染后的临床症状主要表现为胃肠炎、脑膜炎、脑炎、流产等。新生儿、孕妇、老年人以及免疫功能缺陷者较容易感染Lm,对于人类致死率高达20%~30%[1-3]。20世纪90年代,Lm已被WHO世界卫生组织列为食品中的四大致病菌之一。
在导致宿主食源性感染过程中,Lm可以依赖其特有的表面蛋白侵入宿主非吞噬细胞,进而突破宿主肠道屏障、血胎屏障和血脑屏障。内化素家族蛋白是一类含有亮氨酸重复区域(LRRs)序列的表面蛋白,在Lm入侵宿主非吞噬细胞过程中发挥重要的作用[5]。依据内化素蛋白的一级结构,可以将其分为3类:含有LPXTG结构的内化素,含有GW/WxL结构的内化素和分泌型内化素。其中含有LPXTG结构的内化素的C末端LPXTG结构可以被转肽酶A(StrA)识别,并以共价键的方式结合于细胞壁磷壁酸;而GW结构是以甘氨酸和色氨酸为起始的含80个氨基酸序列的重复结构,WxL区域与GW结构类似,均通过非共价键的方式与磷壁酸结合,将蛋白锚定于细菌细胞壁表面[5]。
内化素InlB蛋白由630个氨基酸序列组成,其一级结构包括信号肽序列、7个LRRs序列、IR序列、B重复序列和3个GW序列,是含有GW/WxL结构的典型内化素蛋白。Bielecki的研究表明,inlA和inlB基因均由Lm毒力岛Ⅱ(LIPI-Ⅱ)编码,LIPI-Ⅱ主要介导Lm对宿主非吞噬细胞的粘附侵袭作用,InlA蛋白与细胞表面受体E-钙粘蛋白(E-cad)的相互作用介导Lm对上皮细胞的侵袭作用[2,4]。InlB蛋白主要有3种受体:补体分子受体(gC1qR)、肝细胞生长因子受体(Met)和葡糖氨基聚糖类(GAGs),其中Met为InlB蛋白的主要受体[5]。NTSN_0462基因是具有GW结构的內化素蛋白,为了研究该蛋白的功能,本研究以从暴发李斯特菌病的绵羊脑内分离株LmNTSN为亲本株,通过同源重组的方法构建了NTSN_0462的缺失突变株,并对其生物学特性进行了研究,为今后4b型Lm致病机理的研究提供理论依据。
1 材料与方法
1.1 材料
1.1.1 菌株和质粒 大肠杆菌(Escherichiacoli)DH5α、血清型4b的绵羊脑分离株LmNTSN、穿梭载体pKSV7均由本实验室保存,pGEM-T载体购自大连宝生物工程(TaKaRa)有限公司。
1.1.2 实验动物和细胞 6周龄雌性BALB/c小鼠购自北京维通利华公司。人结肠腺癌细胞系Caco-2由本实验室保存,人肝癌细胞系HepG2购自上海博古科技有限公司。
1.1.3 培养基 试剂BHI培养基(BactoTM Brain Heart Infusion)购自BD公司。
1.1.4 主要试剂和仪器 细菌基因组DNA抽提纯化试剂盒、质粒DNA抽提试剂盒购自北京天根生物科技有限公司,DNA纯化回收试剂盒、DNA聚合酶、高保真DNA聚合酶、限制性核酸内切酶(SalⅠ、KpnⅠ)、DL2000 DNA Marker、λ-14 DNA Marker、T4连接酶、氨苄青霉素(Amp)均购自大连宝生物工程(TaKaRa)有限公司,氯霉素购自生工生物工程公司,PCR扩增仪、凝胶成像系统购自Bio-RAD公司,离心机、电转化仪、Biophotometer分光光度计购自德国Eppendorf公司,恒温CO2培养箱购自Thermo公司。
1.1.5 引物 从NTSN全基因组中获得NTSN_0462基因及其上下游序列,利用Primer Premier 5.0软件设计其上游片段引物P1和P2,引物之间跨度为500 bp;设计其下游片段引物P3和P4,引物之间的跨度为700 bp。引物由南京金斯瑞生物科技有限公司合成,引物序列如表1所示。
表1 目的基因引物序列
Tab.1 Primers sequences of target genes
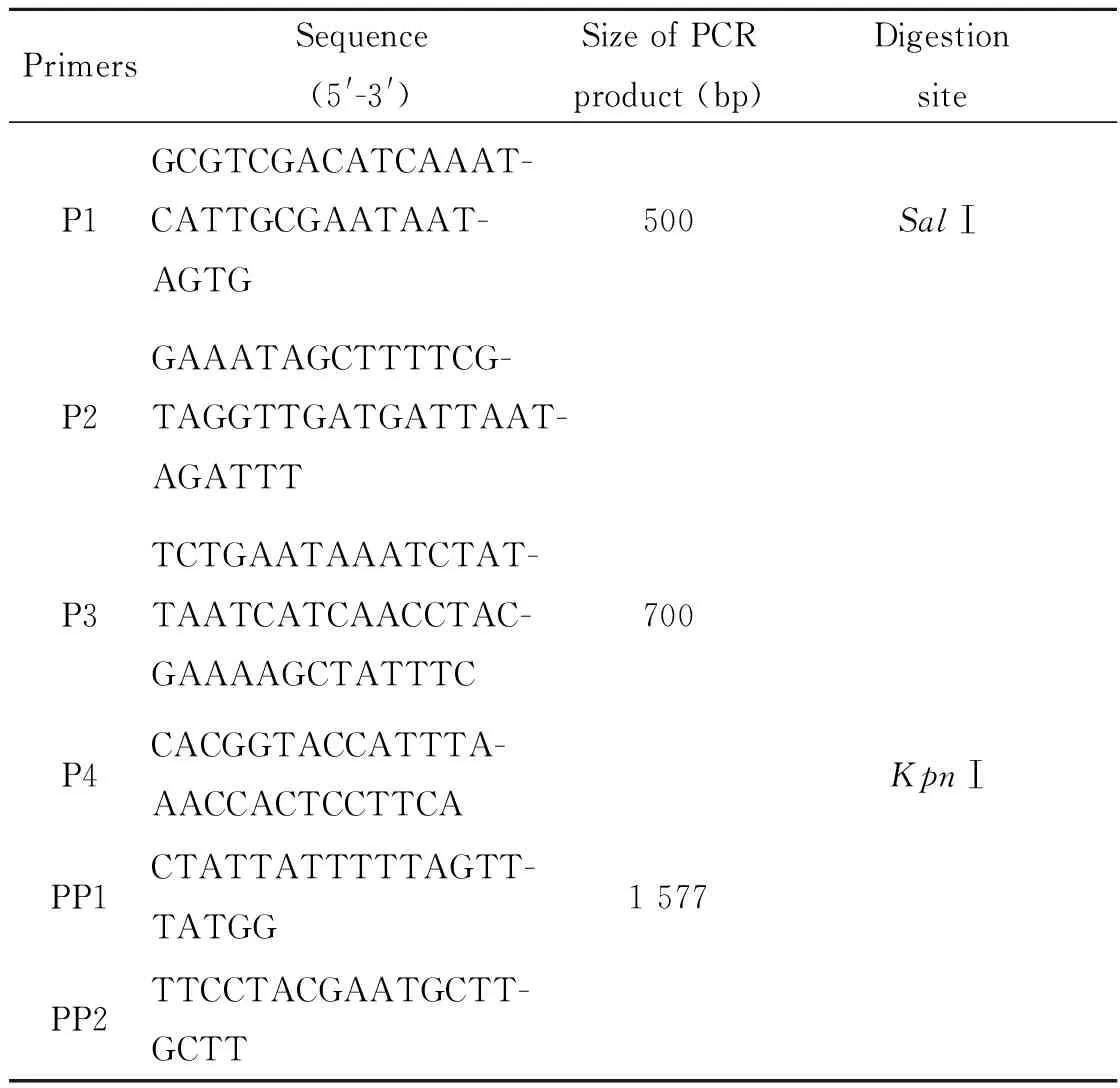
PrimersSequence(5'-3')SizeofPCRproduct(bp)DigestionsiteP1GCGTCGACATCAAAT-CATTGCGAATAAT-AGTG500SalⅠP2GAAATAGCTTTTCG-TAGGTTGATGATTAAT-AGATTTP3TCTGAATAAATCTAT-TAATCATCAACCTAC-GAAAAGCTATTTC700P4CACGGTACCATTTA-AACCACTCCTTCAKpnⅠPP1CTATTATTTTTAGTT-TATGG1577PP2TTCCTACGAATGCTT-GCTT
1.2LmNTSNΔ0462缺失突变株的构建
1.2.1 目的基因的制备 收集LmNTSN过夜培养物,利用细菌基因组DNA提取试剂盒提取NTSN基因组。以得到的基因组为模板,以P1、P2为引物合成NTSN_0462基因上游片段bu;以P3、P4为引物合成NTSN_0462基因下游片段bd。PCR扩增体系:10×PCR buffer 5 μL;dNTP 4 μL;DNA 模板2 μL;P1/P2 1 μL;P3/P4 1 μL;高保真酶0.5 μL;SW 36.5 μL。PCR扩增条件:Tm为55 ℃,30个循环。PCR 产物经1%琼脂糖凝胶电泳回收,以回收产物为模板,以P1、P4 为引物,经SOEing PCR 合成融合基因片段bu/bd。
融合片段bu/bd经DNA纯化回收试剂盒纯化后与pGEM-T载体相连,热击法转化至E.coliDH5α宿主菌中,经过蓝白斑筛选,挑取白色菌落培养,提取质粒进行PCR鉴定,PCR体系如前所述,验证正确后进行SalⅠ和KpnⅠ的双酶切鉴定。将初步鉴定为阳性的克隆pGEM-T-bu/bd送南京金斯瑞生物科技有限公司测序,测序结果通过NCBI中的BLAST进行比对分析,取比对正确的阳性克隆进行保存,并作为下一步构建的实验材料。
1.2.2 目的基因与pKSV7载体的连接 取测序正确的阳性克隆提取质粒,经SalⅠ、KpnⅠ双酶切,酶切产物经DNA纯化回收试剂盒回收,穿梭载体pKSV7经同样的方法处理。对回收后的载体片段和目的基因在T4连接酶作用下于16 ℃恒温器中过夜连接。次日,连接产物通过热击法转化至大肠杆菌E.coliDH5α感受态中。通过PCR和双酶切法鉴定重组质粒,将鉴定正确的质粒送南京金斯瑞生物科技有限公司测序,并将测序正确的质粒命名为pKSV7-bu/bd。
1.2.3LmNTSNΔ0462的构建及鉴定 挑取测序正确的pKSV7-bu/bd质粒,采用电转化的方法(2 100 V)将重组质粒转化至LmNTSN感受态中,电转化涂布于氯霉素(Cm+)抗性(10 μg/mL)BHI平板,置于30 ℃恒温培养箱中培养48 h。挑取菌落进行PCR鉴定,鉴定正确的菌株即为阳性克隆。将获得的阳性克隆在温度(42 ℃)及氯霉素抗性(10 μg/mL)双重压力下传至14代,使目的片段与细菌基因组发生同源重组,将同源重组成功的细菌在30 ℃无抗性压力下脱去抗性载体,获得在氯霉素(10 μg/mL)抗性平板上无法生长而在无抗BHI平板上正常生长的细菌。以NTSN_0462基因上下游臂外围引物PP1、PP2为引物,对获得的菌株进行PCR鉴定,PCR扩增序列送南京金斯瑞生物科技有限公司测序,测序结果经BLAST比对,将鉴定正确的细菌命名为LmNTSNΔ0462。
1.3 细菌生长曲线的测定 将Lm菌株NTSN、NTSNΔ0462分别接种到新鲜BHI培养基中过夜培养,次日取1 mL菌液,12 000 r/min 室温离心2 min。经PBS重悬,测定细菌的OD600值,将细菌分别转接到含有10 mL BHI 培养基的小锥形瓶中,每组细菌设立3个平行重复,并使其初始OD600值为0.05,放于37 ℃摇床培养,间隔1 h 测定1次各瓶的OD600值并做相应记录。
1.4 细胞侵袭实验 对生长状态良好的Caco-2或HepG2细胞进行计数,同时换用无抗DMEM培养基继续培养1 h。按细菌与细胞的数量比(MOI)=20加入新鲜培养的NTSN、NTSNΔ0462菌株继续培养1 h后,细胞用无菌PBS洗3次,加入含100 mg/mL硫酸庆大霉素的DMEM培养基继续培养2 h。PBS洗3次,加入1 mL 0.2% Triton X-100裂解细胞8 min,释放细菌,用无菌PBS稀释裂解液,涂布与BHI平板,置于37 ℃温箱培养20 h后菌落计数,每组设置3个重复。
1.5Lm对BALB/c小鼠的腹腔感染实验 挑单菌落接种于BHI液体培养中,于37 ℃、180 r/min过夜培养,次日将种子液按1∶50的比例重新接种于新鲜的BHI培养基中,于37 ℃、180 r/min振摇培养3 h,经无菌PBS洗涤2次后,调整细菌OD600值至0.8。将6周龄BALB/c雌性小鼠随机分为2,每组5只,腹腔注射免疫。每只小鼠的注射剂量为100 μL,1×104CFU细菌/只,同时将细菌适度稀释后涂布BHI平板,菌落计数。72 h后,分析小鼠肝脏和脾脏中的细菌载量,并进行统计学分析。
2 结 果
2.1LmNTSNΔ0462缺失株的构建
2.1.1 目的基因的制备 通过PCR的方法扩增出NTSN_0462基因上下游片段bu、bd,片段大小分别为500 bp、700 bp。对这两个DNA片段进行SOEing PCR,扩增出一条为1 200 bp的特异性条带。纯化回收,与pGEM-T载体16 ℃过夜连接后转化至E.coliDH5α感受态中。通过双酶切方法验证,目的基因条带大小为1 200 bp,载体条带大小约为3 000 bp,结果正确。将酶切鉴定正确的克隆送南京金斯瑞生物科技有限公司测序,对测序结果进行比对分析,结果表明bu/bd融合片段与pGEM-T载体正确连接。
2.1.2 目的基因与pKSV7载体的连接 双酶切显示目的基因条带大小为1 200 bp,载体条带大小为6 900 bp,结果正确。将酶切鉴定正确的质粒送南京金斯瑞生物科技有限公司测序,对测序结果进行比对分析,表明bu/bd融合片段与pKSV7载体正确连接。
2.1.3LmNTSNΔ0462的构建及鉴定 取测序正确的的重组质粒pKSV7-bu/bd电转化至NTSN感受态中。PCR鉴定转化产物。取PCR结果为阳性的克隆在温度和氯霉素抗性双重选择压力下传至14代,使目的片段与细菌基因组之间发生同源重组。将同源重组成功的细菌在30 ℃无抗性压力下脱去抗性载体,获得在氯霉素(10 μg/mL)的抗性平板上无法生长,在无抗BHI平板可正常生长的菌株。对获得的细菌进行PCR验证,结果如图1所示,在1 200 bp处有一条清晰的目的条带。再以外侧引物PP1、PP2 PCR鉴定获得的细菌,如图2所示,缺失株扩增的目的条带大小为1 577 bp,将PCR扩增产物送南京金斯瑞生物科技有限公司测序,对测序结果进行比对分析,结果表明LmNTSN的inlB基因已成功缺失。

M: DL2000 DNA marker;1-2: PCR product of NTSN-pKSV7-bu/bd.图1 重组菌的PCR筛选Fig.1 Result of mutant strains identified by PCR
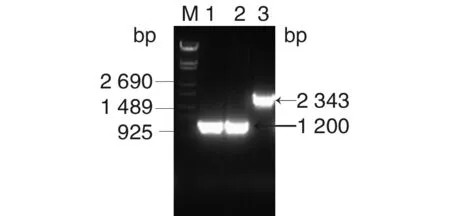
M: λ-14 DNA marker;1-2: PCR product of NTSN Δ0462;3: PCR product of NTSN图2 突变株的鉴定图Fig.2 Identification of mutant strains by PCR
2.2 细菌生长曲线的测定Lm的生长曲线如图3所示,NTSN和NTSNΔ0462的生长趋势基本一致:37 ℃培养环境中,6 h内细菌呈指数形式增长,6 h后,细菌增殖进入平稳期。这表明NTSN_0462基因的缺失未对细菌生长造成明显的影响。
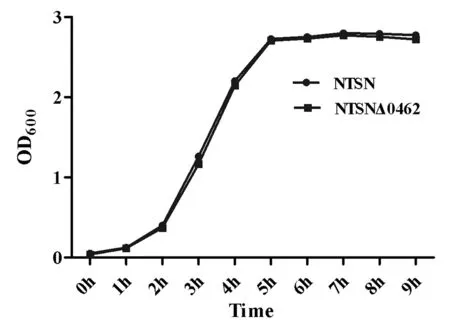
图3 细菌生长曲线的测定Fig.3 The growth curve of Lm at 37 ℃
2.3 细胞侵袭实验 本研究通过相对侵袭率直观地显示出野生株与缺失株对Caco-2以及HepG2细胞侵袭的差别。结果如图4,图5所示,NTSNΔ0462对Caco-2和HepG2的侵袭率低于野生株(P<0.001),表明NTSN_0462基因在Lm对Caco-2和HepG2细胞侵袭过程中发挥重要作用。

图4 Lm对Caco-2细胞系的侵袭Fig.4 Invasion of Lm into Caco-2 cell line

图5 Lm对HepG2细胞系的侵袭Fig.5 Invasion of Lm into HepG2 cell line
2.4Lm对BALB/c小鼠的腹腔感染实验 用NTSN和NTSNΔ0462分别腹腔感染BALB/c小鼠,感染剂量为0.1 LD50,即1×104CFU/只。感染72 h后分析野生株与缺失株在小鼠脾脏、肝脏中的细菌载量。结果如图6所示,NTSNΔ0462缺失株在肝脏中的分离率低于野生株(P<0.01),在脾脏中则没有明显变化,结果表明NTSN_0462基因的缺失导致Lm对小鼠致病力明显下降。

图6 Lm腹腔感染小鼠后组织中的载菌量Fig.6 Bacterial counts in organ homogenates of mice at 72 h i.p. Infected with Lm
3 讨 论
单核细胞增生性李斯特菌为食源性兼性胞内寄生菌,通过污染的食物进入肠道,在其毒力因子作用下突破宿主的肠道屏障、胎盘屏障和血脑屏障进行扩散和传播,从而引发人和40多种动物的感染,具有较高的死亡率[1]。在Lm的致病作用中,内化素蛋白家族(Internalins)是介导该菌粘附与入侵宿主细胞的主要因子,在其致病力方面发挥着重要的作用。NTSN菌株中共有26个內化素家族蛋白,本研究以LmNTSN_0462基因为研究对象,对其致病作用进行了初步的探讨。
InlB是内化素家族中含有GW/WxL结构的典型内化素蛋白,目前对该蛋白的研究主要基于血清型1/2a的EGDe菌株开展的,NTSN与该模式菌株InlB蛋白的同源性虽然达到94%,但在该蛋白的多个LRR结构域、B重复区和GW重复区共存在28个氨基酸的明显差异,为此,本研究利用同源重组的方法成功构建了NTSNΔ0462缺失株,在对缺失株和野生株体内外感染特性进行研究的基础上,探究4b型菌株含有GW结构的內化素蛋白的功能。
在造成宿主食源性感染过程中,Lm可依赖其表面蛋白入侵宿主的巨噬细胞及多种非吞噬细胞,并在胞内存活和增殖[6-9]。内化素蛋白作为细菌表面蛋白,与Lm粘附侵袭非吞噬细胞的过程密切相关[10-13]。本研究中以人结肠腺癌细胞系Caco-2和人肝癌细胞系HepG2细胞系为研究材料,对NTSNΔ0462的粘附侵袭特性进行了研究。结果显示,与亲本株相比,NTSNΔ0462缺失突变株对Caco-2细胞系的侵袭率降低了78%,对HepG2细胞系的侵袭率则降低了98%。缺失突变株对两株细胞系的侵袭率均低于野生株,表明NTSN_0462编码蛋白在LmNTSN侵入上皮细胞和肝细胞时均发挥内化作用,但在入侵肝细胞时的内化作用更为明显。
内化素蛋白InlA、InlB的作用受体具有种属特异性,InlA蛋白不能识别小鼠的细胞表面受体E-钙粘蛋白(E-cad),却可以作用于人、豚鼠和兔子的E-cad;而InlB蛋白与肝细胞生长因子受体(Met)的相互作用则正好与之相反[14-15]。本研究选用BALB/c小鼠作为研究模型,选择腹腔注射途径,对NTSNΔ0462缺失突变株和亲本株体内感染特性进行了测定,结果显示缺失突变株在小鼠肝脏中的细菌载量低于野生株,提示NTSN_0462基因在NTSN侵入肝脏进行定植中发挥重要作用,与体外侵袭试验的结果一致,表明该基因参与了NTSN的体内致病作用。
本研究成功构建了单核细胞增生李斯特菌NTSN_0462基因缺失突变株,并对缺失株的生长、代谢、对细胞的侵袭以及对小鼠的毒力进行了研究,表明该內化素基因在Lm侵袭Caco-2和HepG2细胞系及小鼠肝脏定植中发挥重要作用,是NTSN菌株的重要致病因子,也为揭示NTSN_0462编码蛋白在Lm感染过程中作用机制奠定了理论基础。
[1] Hedberg C. Food-related illness and death in the United States[J]. Emerg Infect Dis, 1999, 5(6): 840-842.
[2] Jiang LL, Chen JS, Fang WH. Advances in the research of main virulence factors ofListeriamonocytogenes[J]. Chin J Zoonoses, 2007, 23(7): 733-735.
江玲丽, 陈健舜, 方维焕. 单核细胞增多性李斯特菌主要毒力因子研究进展[J]. 中国人兽共患病学报, 2007, 23(7): 733-735.[3] de las Heras A, Cain RJ, Bielecka MK, et al. Regulation ofListeriavirulence: PrfA master and commander[J]. Curr Opin Microbiol, 2011, 14(2): 118-127.
[4] Bielecki J, Younqman P, Connelly P, et al. Bacillus subtilis expressing a haemolysin gene fromListeriamonocytogenescan grow in mammalian cells[J]. Nature, 1990, 345(6271): 175-176.
[5] Bierne H, Sabet C, Personnic N, et al. Internalins: a complex family of leucine-rich repeat-containing proteins inListeriamonocytogenes[J]. Microbes Infect, 2007, 9(10): 1156-1166.
[6] Niqhtinqale KK, Windham K, Martin KE, et al. SelectListeriamonocytogenessubtypes commonly found in foods carry distinct nonsense mutations in inlA, leading to expression of truncated and secreted internalin A, and are associated with a reduced invasion phenotype for human intestinal epithelial cells[J]. Appl Environ Microbiol, 2005, 71(12): 8764-8772.
[7] Greqory SH, Saqnimeni AJ, Winq EJ. Internalin B promotes the replication ofListeriamonocytogenesin mouse hepatocytes[J]. Infect Immun, 1997, 65(12): 5137-5141.
[8] Greiffenberg L, Goebel W, Kim KS, et al. Interaction ofListeriamonocytogeneswith human brain microvascular endothelial cells: InlB-dependent invasion,long-term intracellular growth, and spread from macrophages to endothelial cells[J]. Infect Immun, 1998, 66(11): 5260-5267.
[9] Cabanes D, Sousa S, Cebria A, et al. GP96 is a receptor for a novelListeriamonocytogenesvirulance factor, Vip, a surface protein[J]. EMBO J, 2005, 24(15): 2827-2838.
[10] Sahu SC, Gaines DW, Williams KM, et al. A synthetic polypeptide based on Human E-cadherin inhibits invasion of human intestinal and liver cell lines byListeriamonocytogenes[J]. J Med Microbiol, 2007, 56(8): 1011-1016.
[11] Chiba S, Naqai T, Hayashi T, et al.Listerialinvasion protein internalin InlB promotes entry into ideal Peyer's patchesinvivo[J]. Microbiol Immunol, 55(2): 123-129.
[12] Pentecost M, Kumaran J, Ghosh P, et al.Listeriamonocytogenesinternalin B activates junctional endocytosis to accelerate intestinal invasion[J]. PLoS Pathoq, 2010, 6(5): e1000900.
[13] Maetins M, Custodio R, Camejo A, et al.Listeriamonocytogenestriggers the cell surface expression of Gp96 protein and interacts with its N terminus tosupport cellular infection[J]. J Biol Chem, 2012, 287(51): 43083-43093.
[14] Bonazzi M, Lecuit M, Cossart P.Listeriamonocytogenesinternalin and E-cadherin: from structure to pathogenesis[J]. Cell Microbiol, 2009, 11(5): 693-702.
[15] Sahu SC, Gaines DW, Williams KM, et al. A synthetic polypeptide based on human E-cadherin inhibits invasion of human intestinal and liver cell lines byListeriamonocytogenes[J]. J Med Microbiol, 2007, 56(8): 1011-1016.
Function of an internalin geneNTSN_0462 inListeriamonocytognesserotype 4b
ZHAO Dan, YAO Hao, CAI Xue-xue, KONG Su-wei, YUN-Xi, TAN Wei-jun, PAN Zhi-ming, YIN Yue-lan, JIAO Xin-an
(JiangsuCo-InnovationCenterforPreventionandControlofImportantAnimalInfectiousDiseaseandZoonoses,JiangsuKeyLaboratoryofZoonosis,YangzhouUniversity,JointInternationalResearchLaboratoryofAgricultureAgri-ProductSafety,Yangzhou225009,China)
Internalins play a key role in the adhesion and invasion ofL.monocytogenesinto host cells. In this study, the function of internalin geneNTSN_0462 in serotype 4bLmNTSN was probed into. Firstly,NTSN_0462 was knocked out by homology recombination, then the pathogenic characterization of mutant strainLmNTSNΔ0462 was investigatedinvivoandinvitro. The growth curve showed that the deletion ofNTSN_0462 gene didn't affect growth and metabolism ofLmin BHI medium. The invasion of the mutant strain into human intestinal epithelial cell line Caco-2 and human hepatocyte cell line HepG2 significantly reduced comparing with the WT strains (P<0.001). Furthermore, the infection ability to BALB/c mouse was analyzedinvivo, compared with the WT strains, the colonization ofLmNTSNΔ0462 in liver decreased significantly (P<0.001), wheareas there was no significant difference in the spleen, indicating thatNTSN_0462 plays an important role in the invasion and multiplication in host livers, heretofore it is an invasion-related virulent factor ofL.monocytogenes.
Listeriamonocytogenes;NTSN_0462; serotype 4; internalin
Yin Yue-lan, Email: yylan@yzu.edu.cn
10.3969/j.issn.1002-2694.2017.02.005
国家自然科学基金 (Nos. 31472193, 31101841),江苏高校优势学科建设工程项目资助
扬州大学/江苏省人兽共患病重点实验室/江苏省动物重要疫病和人兽共患病防控协同创新中心/农业与农产品安全国际合作联合实验室,扬州 225009
R378
A
1002-2694(2017)02-0115-05
2016-09-26 编辑:刘岱伟
Supported by the National Natural Science Foundation of China (Nos. 31472193 & 31101841) and the Priority Academic Program Development of Jiangsu Higher Education Institutions
猜你喜欢
——一道江苏高考题的奥秘解读和拓展
中学生物学(2022年7期)2022-09-07
成都医学院学报(2022年4期)2022-08-19
科学与生活(2021年16期)2021-11-25
江西农业学报(2021年4期)2021-04-20
浙江医学(2020年19期)2020-10-20
少先队活动(2020年8期)2020-09-11
少先队活动(2020年7期)2020-08-14
三农资讯半月报(2020年11期)2020-06-21
福建基础教育研究(2019年9期)2019-05-28
华人时刊(2018年15期)2018-11-10
