
植物根际促生细菌定殖研究进展
2017-02-22 06:51:08孙真郑亮邱浩斌
生物技术通报 2017年2期
孙真郑亮邱浩斌
(1. 合肥工业大学,宣城 242000;2. 中国水产科学研究院东海水产研究所,上海 200090;3. 萍乡学院工程与管理学院,萍乡 337000)
植物根际促生细菌定殖研究进展
孙真1郑亮2邱浩斌3
(1. 合肥工业大学,宣城 242000;2. 中国水产科学研究院东海水产研究所,上海 200090;3. 萍乡学院工程与管理学院,萍乡 337000)
植物促生菌(plant grwth-promoting bacterium,PGPB)因可有效抑制根际病原菌,促进植物生长,增加作物产量等作用,在农业生产领域展现出巨大前景。以了解植物根际促生细菌的种类,理解定殖相关的促生机制为目的,展示了根际细菌的定殖过程及影响细菌在根部定殖的因素,通过抗生素标记、免疫学方法、外源基因标记等方法进行定殖微生物的检测,以高通量测序技术在根际定殖中的广泛应用为结果,得出结论应从遗传水平上对菌株进行解读,获得植物根际土壤微生物群落多样性,功能基因等研究,这项工作对预测根际促生细菌与植物的交互作用,生物菌剂的田间应用有重要意义,应是植物根际促生细菌今后的研究方向。
定殖;根际菌;促生;高通量测序
促生微生物与植物生长之间关系密切,其具有可抑制植物病原菌发生、促进植物对营养物质的吸收等功能。植物根际促生细菌兼具促进植物生长并控制病害发生的功能,成为有机农业发展的希望[1,2]。固氮作用、矿物质溶解、植物生长素释放等都是根际细菌促生的重要途径。但将实验室条件下促生效果显著的微生物应用到野外却不能发挥好的效果[3]。无论是温室或野外条件,微生物在根部的定殖过程都非常复杂,定殖研究将为根际细菌在复杂生态系统提供新视角,对生物菌剂的田间应用提供重要的理论支持。目前,越来越多的研究筛选并证实植物促生菌(plant grwth-promoting bacterium,PGPB)具有促生防病效果[4]。据此,笔者总结了植物根际促生细菌的种类、促生机制及定殖微生物的检测方法、影响微生物在根部定殖的因素等,认为传统形态学与分子生物学技术相结合可以更好的跟踪PGPB在根际的变化及所在生境的微生物群落演替规律,这对于更好的预测PGPB与植物的交互作用,以及生物菌剂的田间应用有重要意义。
1 植物根际定殖菌的种类
国内外已发现假单胞菌属(Pseudomonas)、芽孢杆菌属(Bacillus)、固氮菌属(Azotobacter)、肠杆菌属(Enterobacter)、节杆菌属(Arthrobacter)、伯克氏菌属(Burkholderia)等20多个种属具有促生防病潜能,研究最多是假单胞菌属、芽孢杆菌属[5]。枯草芽孢杆菌PTS-394[6]、芽孢杆菌YPP-9[7]均可定殖番茄根际,有效预防番茄枯萎病,枯草芽孢杆菌FB17[8]可发挥运动性、产生根系分泌物有效定殖拟南芥根系。巨大芽孢杆菌C4[9]在玉米、水稻根部有效定殖。解淀粉芽孢杆菌NJN-6[10]定殖香蕉根部促进生长。B.malacitensis Z-5[11]、B.subtilis GB03、B.amyloliquefaciens IN937a、B.polymyxa E681[12]、B.cereus[13,14]定殖寄主根系。绿针假单胞菌O6[12]产生挥发性代谢产物,如水杨酸和茉莉酸,并以产物为信号诱导植物产生免疫反应。荧光假单胞菌CV6可产IAA、嗜铁素、结瘤及拮抗作用促进黄瓜生长[15]。P.fluorescens 92rkG5[16]及P.aeruginosa IE-6S[17]定殖于番茄根部,前者依赖于根系分泌物,后者对逆境则有较好的耐性。假单胞菌DSMZ13134[18]可定殖于大麦根部且温室和大田的定殖特征相似。P.putida和P.fluorescens[19]通过天仙子碱和莨菪碱有效定殖。
生癌肠杆菌定殖于木豆根际[13],阴沟肠杆菌[3]定殖于水稻根部。水生拉恩氏菌HX2,可溶磷并产生IAA,定殖于葡萄、番茄、向日葵根部[20]。醋杆菌属G.diazotrophicus[2]定殖于高粱和小麦根系,滋养节杆菌Ar13[21]防效达46.60%,定殖于绿豆根际。向日葵根际筛选出伯克氏菌(Burkholderia sp.)、固氮螺菌(Azospirillum sp.)、志贺氏杆菌(Shigella sp. )、新鞘脂菌(Novosphingobium sp.)、无色菌(Achromobacter sp.)等,均可产生铁细胞,新鞘脂菌、伯克氏菌、无色菌、固氮螺菌产生吲哚类物质,伯克氏菌和无色菌则具有溶磷作用[22]。克雷白氏杆菌NG14[23]通过nifH基因在水稻根系定殖参与固氮作用。草螺菌[24](H. frisingense)定殖大麦根系,草螺菌RAM10[25]利用piIT基因控制菠萝上的定殖。根瘤菌[26]定殖于大豆根系,其定殖能力受胁迫环境影响减弱。Boehm[27]发现固氮弧菌BH72菌株通过piIT基因控制在水稻根部的定殖能力。根瘤菌ORS-571、固氮菌MTCC-446可通过结瘤、固氮、耐性等促进植物生长[28]。苍白杆菌KUDC1013[29]定殖后可利用系统诱导抗性(ISR)和脂多糖诱导(LPS)抵御软腐病害。伯克氏菌KJ006的群体感应信号可防治水稻谷枯病[30]。
植物-微生物体系是一个完整的微生态系统,植物体内、体表和根际均分布大量的微生物,尽管PGPB在这一体系可发挥有益作用,其他微生物的种群和数量同样对植物的产量、品质、防病等具有极大影响,且其群落结构和数量也处于不断变化。以往PGPB的根际定殖研究多集中在单一或某几类微生物在根际的定殖效果,常见于从根际中筛选具有促生功能的细菌类群,且此类研究多由于田间应用效果而受到限制。了解根际定殖菌的种类,尤其是对该生态系统其他微生物群落结构的影响,对于田间促生效果的发挥具有重要意义。
2 定殖相关的促生机制
目前,PGPB的研究还集中在微生物在根际的促生防病机制[31],见表1。产生刺激性植物激素、挥发性化合物、ACC脱氨酶等,降低植物体乙烯水平,改进植物营养状况(如促进难溶性磷、钾和微量元素的释放及非共生固氮等)及诱导植物产生ISR均属于直接促生作用。间接促生作用则包括,产生抗生素、抗菌蛋白或铁载体等减轻病害,加强共生作用(根瘤和菌根的形成),降解农药等生物外源性物质。
荧光假单胞菌(P. fluorescens)FPT9601-T5及赛维瓦尔芽孢杆菌(P. thivervalensis)MLG45均可产生IAA,缓解定殖菌在寄主植物生理进程中受到的环境压力,直接刺激植物增长[32]。部分定殖菌可产乙烯,或利用外源乙烯和其前体ACC抑制病原菌感染,或形成根瘤,通过负反馈调节定殖[33]。部分PGPB产生铁细胞可螯合三价铁离子,能与LPS发生ISR[22]。水杨酸(SA)、茉莉酸(JA)可作为信号诱导植物产生免疫反应来调控微生物的定殖[12]。辅助因子吡咯喹啉醌(PQQ)与ISR辅助子相关[34]。细胞分裂素(CTK)促进根细胞分裂、增大根表面积,来影响菌株的定殖数量,赤霉素刺激叶芽生长,改变植物形态来影响定殖[35]。
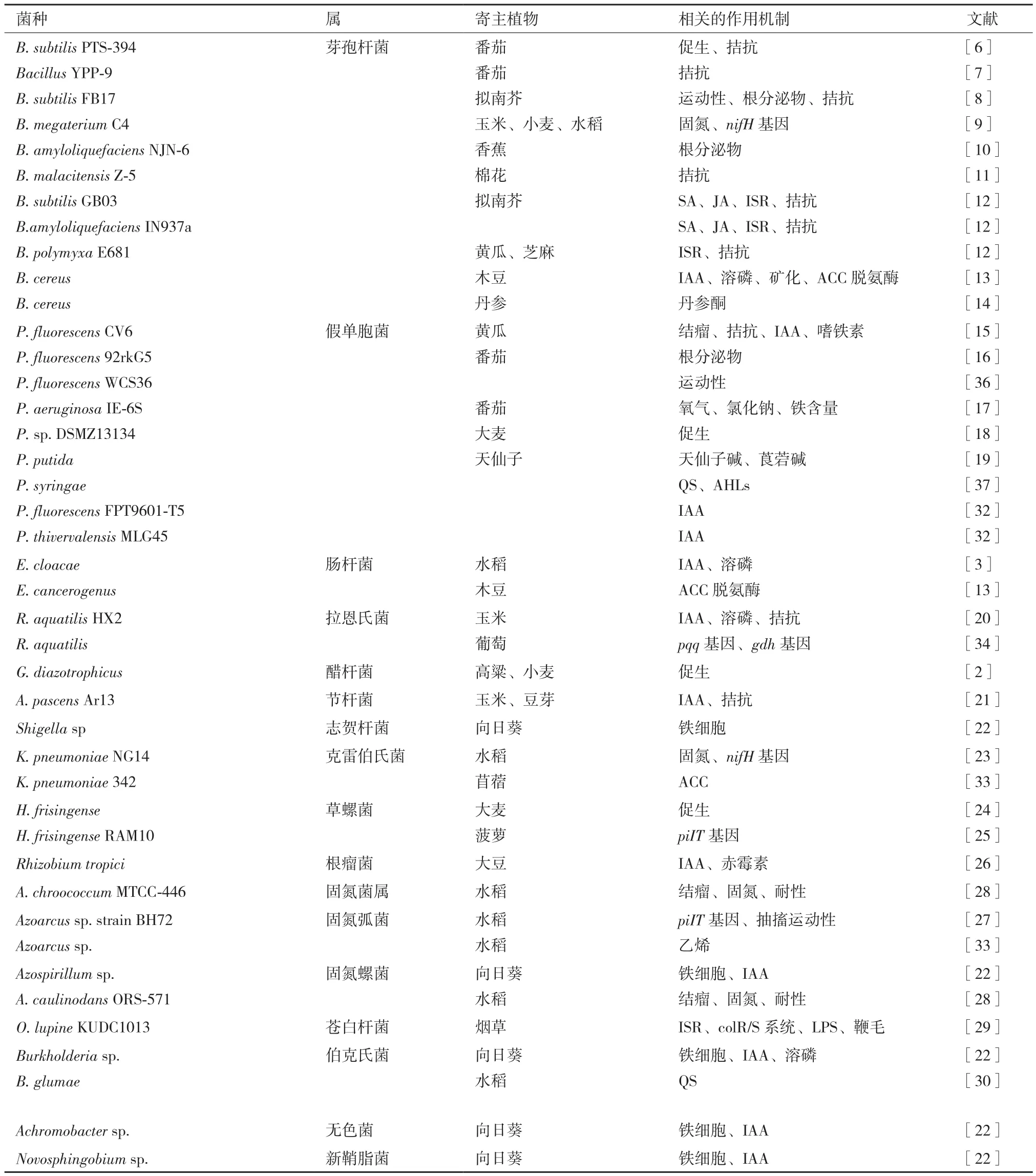
表1 PGPB及定殖相关的生物学特征
荧光假单胞菌WCS365野生型菌株和非营养缺陷的Tn5随机转座突变株的根尖定殖对比试验表明细菌运动性相关基因可影响定殖[36]。运动性和对根分泌物的趋化性是细菌的重要定植特性,如苹果酸内酯是生防枯草芽孢杆菌FB17菌株的主要化学引诱物[8]。非特异性磷酸酶、植酸酶和C-P 裂解酶分解土壤中有机磷化合物以释放可溶性磷,无机磷酸盐中可溶性磷的释放与有机酸形成有关。QS信号扩散由N-酰基高丝氨酸内酯(AHLs)和N-3-羰基六酰基高丝氨酸内酯(HSL)两种物质传递[37]。 苍白杆菌KUDC1013利用ISR抵御软腐病害,colR/S系统、LPS氧化位点、鞭毛等都参与ISR[29]。piIT基因[25,27]可控制菌毛收缩,缺失该基因的突变体丧失抽搐运动性,影响菌株定殖能力。
综上所述,IAA、乙烯、ACC脱氨酶、铁细胞、赤霉素、细胞分裂素、水杨酸、茉莉酸、类黄酮及细胞脱落酸等物质均可参与调控根际微生物定殖,此类分泌物的产生通常是细菌或植物体内编码基因及酶的调控。生物固氮、溶磷、群体感应、ISR机制、NADH脱氢酶、LPS氧化位点、IV-菌毛及抽搐运动性等机制也可调控植物的定殖,此类机制是否发生效应除了依靠信号分子外,基因的作用至关重要。因此,在植物根际促生菌研究中,除了回答所在生境有何物种,哪些物种有促生作用,生境中微生物群落的演替变化、还需要弄清楚这些物种具有怎样的基因功能,各种促生机制如何发挥作用,则需要高通量测序技术,尤其是宏基因组学在PGPB研究中的应用。
3 根际细菌的定殖过程及影响因素
土壤接种微生物后单个零星细胞先到达根系表面,成对的包裹于整个根被皮,并成线状排列于表面,后密集覆盖于整个根系,甚至在寄主植物上形成包裹的生物膜。根表细菌的侵入多依靠被动方式,可通过侧根裂隙、气孔、水孔及茎皮孔等自然开口,以及因土壤摩擦、动物取食或人为伤害的伤口进入。如笔者将接种了HX2gfp的幼苗根部,用水冲洗在显微镜下能观察到大量菌株,同时进行消毒处理后制成冰冻切片,横截面却未观测到菌株,该细菌仅定殖于根表[39]。内生细菌多以主动方式侵入,不但在根围成功定殖还可进入植物体内,从根部一直侵染到茎、叶、花和果实等部位,发挥促生效应[33]。如巨大芽孢杆菌C4[9],荧光假单胞菌92rkG5[16],内生细菌01-144[38],与根表细菌相比,内生细菌对寄主要求更高、专一性更强。
PGPB的定殖受一系列生物环境因素影响,包括根系分泌物、植物生长状况、细菌趋化性、自我调节机制及细菌营养型等。如香蕉根系分泌物吸引解淀粉芽孢杆菌(B. amyloliquefaciens)NJN-6定殖[10],假高粱根系分泌物对PGPB的定殖有选择性[40]。植物改变根系建筑学结构可诱导不同种群定殖[41],根系伸长速率影响假单胞菌积累[42]。此外,细菌通过遗传控制鞭毛/运动性、趋化性、多糖、位点特异重组酶/菌落阶段变异、NADH脱氢酶、根系分泌物和植物种类[43],水生拉恩示菌pqq和gdh基因被破坏后生防效果降低[21],细菌B3-7重新获得运动性基因时定殖生防能力恢复[44]。同时,番茄根部易分化部位的荧光假单胞菌92rkG5与根尖的细菌生理型不同[16]。PGPB能否定殖,还受到土壤温度、质地、含水量、衬质势及含氧量等非生物因素的影响。如干旱胁迫影响大豆根际接种多黏类芽孢杆菌(P. polymyxa)及热带根瘤菌(R. tropici)的定殖[26],低温下植物生长缓慢,土壤空隙可为根际菌提供保护位点,细菌扩散借助于水分,合适的氧气、氯化钠浓度、铁含量可促进铜绿假单胞菌(P. aeruginosa)IE-6S[17]在番茄的定殖。因此,多种外界因素影响着根际菌竞争能力的发挥。
PGPB定殖特征具有普遍性,其影响因素也被众多学者认可。然而,涉及微生物遗传层面,影响PGPB定殖的因素如何与自身的基因功能作用,对相应的细菌基因进行注释,了解定殖菌的代谢通路并进行功能注释则需要研究者更多的探索。
4 定殖微生物的检测方法
植物根际促生细菌的定殖检测方法包括抗生素标记、免疫学方法及外源基因标记等。抗生素标记法可保持原始菌株的稳定遗传性,抗利福平和抗病原真菌法鉴定出B. malacitensis Z-5有效定殖棉花根际[11]。免疫学方法包括荧光标记免疫,酶联免疫吸附(ELISA)等。例如,ELISA方法证明PRPG菌株可减少黄瓜花叶病毒对番茄类植物的感染[45]。外源及标记基因多为编码酶的基因,通过检查表达产物来监测定殖状况,此类基因包括lacZ(编码大肠杆菌的β-半乳糖苷酶)、gusA(编码大肠杆菌的β-葡萄糖醛酸糖苷酶)、luxAB(编码细菌荧光素酶)和gfp(绿色荧光蛋白基因)。目前,使用最多的还是gfp标记,该方法不需要底物及辅助因子即能表达,对细胞及组织无毒害作用,稳定性好,有多种颜色和强度的突变体[46]。如标记了gfp基因的枯草芽孢杆菌PTS-394在番茄根际有效定殖[6],笔者通过电击转入带有gfp表达的质粒pSMC21,在488 nm激发波长下观察到HX2gfp菌株为亮绿色荧光短杆状菌体[39]。另外,根据gfp基因的特点还可使用基因探针、分子杂交、PCR等分子生物学方法。
各种微生物的定殖检测方法可明确微生物在植物根际定殖的机制和微生物定殖的特性,在微生物与环境相互作用具有较好的应用价值,其中分子生物方法对于PGPB在根际的定殖研究起到了重要的推动作用。
5 高通量测序技术在根际定殖中的应用
21 世纪以来,人类基因组计划开启大数据时代,各国研究者对微生物的研究已从形态特征分析、生理生化分析、分子生物学技术这些传统手段向高通量测序技术分析逐渐转化,随着高通量测序技术和数据处理能力的发展,目前研究包括:针对rRNA为主要对象的群落多样性研究;以环境中所有遗传物质为对象的群落功能研究及以环境中所有转录本为对象的群落转录调控规律研究。
针对16S rRNA的微生物群落多样性研究有助于准确分析定殖在植物根际和非根际土壤的细菌组成和分布,测序的结果能够更加深入地了解细菌群落组分在科、属和OUT水平上的变化。如利用16S rRNA对南极洲维管束植物根际土壤细群落的研究[47],利用rRNA测序发现施用生物有机肥可调节根际中微生物群落结构,增强黄瓜枯萎病的抵抗力[48]。以环境中所有遗传物质为对象的群落功能研究,除了有微生物的分类信息外更包含了环境中微生物的全部基因信息,有助于对群落潜在的功能进行挖掘,可通过测序序列找到特定环境下富集的功能基因。固氮菌等广泛分布于植物根际,兼具促生、抑制病原菌可定殖于植物根系,如利用高通量测序获得了土壤中涉及β-内酰胺酶的细菌功能类群,这些类群决定了土壤细菌对药物的抗性[49]。利用MiSeq平台的高通量测序虽获得nifH固氮菌[50]、nirS反硝化菌[51]根际的群落多样性,却较少涉及根际中影响N元素代谢途径的调控基因,以及根际中涉及固氮功能的组学研究。以环境中所有转录本为对象的群落转录调控规律研究,可实时、实地的反映微生物群落的基因表达,如对解淀粉芽孢杆菌SQR9的全转录组分析表明,该菌株定殖根部期间,受玉米分泌物影响生物膜的形成能力增强[52]。
6 展望
目前,筛选新的可定殖于植物根际的促生菌株研究、利用gfp基因展示促生菌在植物根系的迁移运动规律、判定PGPB促生机制的种类,以及实验室条件下PGPB对植物的促生防病效果研究,虽丰富了PGPB的数据库给农业应用带来希望,但越来越多的同类型研究报道已难在高质量期刊上占有一席地位,依靠高通量测序等技术对涉及定殖行为的基因调控研究,以及根际中影响菌株代谢途径的宏基因组学研究逐渐成为热点。弄清楚实验室环境下的促生防病效果显著的PGPB难以在自然环境下发挥功效的原因仍具有巨大挑战,首先弄清限菌与大田环境根际微生物的群落差异,通过引入PGPB根据微生物群落的变化,找到影响PGPB效果发挥的关键类群,进一步优化菌株的田间应用效果。此外,加深涉及定殖性状的基因研究,了解信号分子如何在定殖过程发挥调控作用,以及一系列中间产物的迁移转化规律也将促进PGPB在农业的应用。
因此,利用高通量测序技术从遗传水平上对菌株进行解读,获得植物根际土壤微生物群落多样性,功能基因及代谢通路分析有助于加深研究者对植物-根际菌相互关系的理解,传统形态学、生理学与分子生物学技术的结合,对于更好的预测PGPB与植物的交互作用,解决PGPB在田间与限菌条件下定殖能力的差异,推动生物菌剂的商业化发展意义重大,应为未来研究主要方向。
[1]Bhattacharyya PN, Jha DK. Plant growth-promoting rhizobacteria(PGPR):emergence in agriculture[J]. World Journal of Microbiology Biotechnology, 2012, 28(4):1327-1350.
[2]Luna MF, Galar ML, Aprea J, et al. Colonization of sorghum and wheat by seed inoculation with Gluconacetobacter diazotrophicus[J]. Biotechnology Letters, 2010, 32(8):1071-1076.
[3]Shankar M, Ponraj P, Llakkiam D, et al. Root colonization of a rice growth promoting strain of Enterobacter cloacae[J]. Journal of Basic Microbiology, 2011, 51(5):523-530.
[4]Cindy HW, Stéphanie MB, Gary LA, et al. Developing microbe-plant interactions for applications in plant-growth promotion and disease control, production of useful compounds, remediation and carbon sequestration[J]. Microbial Biotechnology, 2009, 2(4):428-440.
[5]Compant S, Clement C, Sessitsch A. Plant growth-promoting bacteria in the rhizo- and endosphere of plants:Their role, colonization, mechanisms involved and prospects for utilization[J]. Soil Biology Biochemistry, 2010, 42(5):669-678.
[6]刘邮洲, 梁雪杰, 乔俊卿, 等. 枯草芽孢杆菌PTS-394的GFP标记及定殖能力[J]. 植物保护学报, 2014, 41(4):416-421.
[7]黎志坤, 朱红惠. 一株番茄青枯病生防菌的鉴定与防病、定殖能力[J]. 微生物学报, 2010, 50(3):342-349.
[8]Rudrappa T, Czymmek KJ, Pare PW, et al. Root-secreted malic acid recruits beneficial soil bacteria[J]. Plant Physiol, 2008, 148(3):1547-1556.
[9]Liu XM, Zhao HX, Chen SF. Colonization of maize and rice plants by strain Bacillus megaterium C4[J]. Current Microbiology, 2005, 52:186-190.
[10]Yuan J, Zhang N, Huang Q, et al. Organic acids from root exudates of banana help root colonization of PGPR strain Bacillus amyloliquefaciens NJN-6[J]. Scientific Reports, 2015, 5:13438.
[11]张冬冬, 刘涛, 高同国, 等. 棉花黄萎病拮抗细菌Z-5菌株的定殖能力检测[J]. 棉花学报, 2013, 25(6):510-516.
[12]Farag MA, Zhang H, Ryu CM. Dynamic chemical communication between plants and bacteria through airborne signals:induced resistance by bacterial volatiles[J]. Journal of Microbiology, 50(3):380-385.
[13]Rahi MU, Arundhathi, Reddy G. Bacillus cereus and Enterobacter cancerogenus screened for their efficient plant growth promoting traits rhizobacteria(PGPR)and antagonistic traits among sixteen bacterial isolated from rhizospheric soils of Pigeon pea[J]. African Journal of Microbiology Research, 2011, 5(15):2090-2094.
[14]Zhao J, Zhou L, Wub J. Promotion of Salvia miltiorrhiza hairy root growth and tanshinone production by polysaccharide-protein fractions of plant growth-promoting rhizobacterium Bacillus cereus[J]. Process Biochemistry, 2010, 45:1517-1522.
[15]Maleki M, Mostafaee S, Mokhtarnejad L, et al. Characterization of Pseudomonas fluorescens strain CV6 isolated from cucumber rhizosphere in Varamin as a potential biocontrol agent[J]. Australian Journal of Crop Science, 2010, 4(9):676-683.
[16]Gamalero E, Lingua G, Tombolini R, et al. Colonization of tomato root seedling by Pseudomonas fluorescens 92rkG5:Spatiotemporal dynamics, localization, organization, viability, and culturability[J]. Microbial Ecology, 2005, 50:2, 289-297.
[17]Siddiqui IA, Shaukat SS, Khan GH, et al. Suppression of Meloidogyne javanica by Pseudomonas aeruginosa IE-6S+in tomato:the influence of NaCl, oxygen and iron levels[J]. Soil Biology and Biochemistry, 2003, 35(12):1625-1634.
18]Buddrus-Schiemann K, Schmid M, Schreiner K, et al. Root colonization by Pseudomonas sp. DSMZ13134 and impact on the indigenous rhizosphere bacterial community of barley[J]. Microbiology Ecology, 2010, 60(2):381-393.
[19]Ghorbanpour M, Hosseini S, Rezazadeh M, et al. Hyoscyamine and scopolamine production of black henbane(Hyoscyamus niger)infected with Pseudomonas putida and P. fluorescens strains under water deficit stress[J]. Planta Medica, 2010, 76(12):167.
[20]Chen F, Guo YB, Wang JH, et al. Biological control of grape crown gall by Rahnella aquatilis HX2[J]. Plant Disease, 2007, 91(8):957-963.
[21]郭岩彬, 吴文良, 孙真, 等. 滋养节杆菌Ar13及其作为植物根际促生细菌的应用:中国, 201210103736. 2[P]. 2012-08-22.
[22]Ambrosini A, Beneduzi A, Stefanski T, et al. Screening of plant growth promoting Rhizobacteria isolated from sunflower(Helianthusannuus L.)[J]. Plant and Soil, 2012, 356(1):245-264.
[23]Liu Y, Wang H, Sun XL, et al. Study on Mechanisms of colonization of Nitrogen-Fixing PGPB, Klebsiella pneumonia NG14 on the root surface of rice and the formation of biofilm[J]. Current Microbiology, 2011, 62:1113-1122.
[24] Rothballer M, Eckert B, Schmid M, et al. Endophytic root colonization of gramineous plants by Herbaspirillum frisingense[J]. FEMS Microbiology Ecology, 2008, 66:85-95.
[25]Baldotto LEB, Olivares FL, Bressan-Smith R. Structural interaction between gfp-labeled diazotrophic endophic bacterium Herbaspirillum seropedicae RAM10 and pineapple plantlets vitoria[J]. Brazillian Journal of Microbiology, 2011, 42:114-125.
[26]Figueiredo MVB, Burity HA, Martinez CR, et al. Alleviation of drought stress in the common bean(Phaseolus vulgaris L.)by co-inoculation with Paenibacillus polymyxa and Rhizobium tropici[J]. Applied Soil Ecology, 2008, 40(1):182-188.
[27] Boehm M, Hurek T, Reinhold HB. Twitching motility is essential for endophytic rice colonization by the N-2-fixing endophyte Azoarcus sp. strain BH72[J]. Molecular Plant-Microbe Interactions, 2007, 20(5):526-533.
[28]Joe MM, Jaleel CA, Sivakumar PK, et al. Co-aggregation in Azospirillum brasilensense MTCC-125 with other PGPR strains:effect of physical and chemical factors and stress endurance ability[J]. Journal of Taiwan Institute of Chemical Engineers, 2009, 40(5):491-499.
[29]Sumayo M, Hahm MS, Ghim SY, et al. Determinants of plant growth-promoting Ochrobactrum lupini KUDC1013 involved in induction of systemic resistance against Pectobacterium carotovorum subsp carotovorum in tobacco leaves[J]. Plant Pathology Journal, 2013, 29(2):174-181.
[30]Cho HS. Park SY, Ryu CM, et al. Interference of quorum sensing and virulence of the rice pathogen Burkholderia glumae by an engineered endophytic bacterium[J]. FEMS Microbiology Ecology, 2007, 60(1):14-23.
[31]Ahemad M, Kibret M. Mechanisms and applications of plant growth promoting rhizobacteria:current perspective[J]. Journal of King Saud University:Science, 2014, 26(1):1-20.
[32]Spaepen S, Vanderleyden J. Auxin and plant-microbe interactions[J]. Cold Spring Harbor Perspectives in Biology, 2011, 3(4):a001438.
[33]Pablo RH, Leo SVO, Jan DVE. Properties of bacterial endophytes and their proposed role in plant growth[J]. Trends in Microbiology, 2008, 16:463-471.
[34]Guo YB, Li JY, Li L, et al. Mutations that disrupt either the pqq or the gdh gene of Rahnella aquatilis abolish the production of an antibacterial substance and result in reduced biological control of grapevine crown gall[J]. Applied and Environmental Microbiology, 2009, 75(21):6792-6803.
[35]Chauhan H, Bagyaraj DJ, Selvakumar G, et al. Novel plant growth promoting rhizobacteria-prospects and potential[J]. Applied Soil Ecology, 2015, 95:38-53.
[36] Lugtenberg B, Kamilova F. Plant-Growth-Promoting Rhizobacteria[J]. Annual Review Microbiology, 2009. 63:541-556.
[37]Shephard RW, Lindow S. Two dissimilar N-acyl-Homoserine lactone acylases of Pseudomonas syringae influence colony and biofilm morphology[J]. Applied Environment Microbiology, 2009, 75(1):45-53.
[38]龙良鲲, 肖崇刚. 内生细菌01-144在番茄根茎内定殖的初步研究[J]. 微生物学通报, 2003, 30(5):53-56.
[39]孙真. 拉恩氏水生菌(Rahnella aquatilis)HX2在玉米根系的定殖研究[D]. 北京:中国农业大学, 2010.
[40]刘纯, 黄红娟, 张朝贤, 等. 假高粱根系分泌物对土壤细菌群落多样性的影响[J]. 生态环境学报, 2013, 22(7):1124-1128.
[41]Hardoim PR, van Overbeek S, van Elsas JD. Properties of bacterial endophytes and their proposed role in plant growth[J]. Trends in Microbiology, 2008, 16(10):463-471.
[42]Watt M, McCully ME, Kirkegaard JA. Soil strength and rate of root elongation alter the accumulation of Pseudomonas spp. and other bacteria in the rhizosphere of wheat[J]. Functional Plant Biology, 2003, 30(5):483-491.
[43]年洪娟, 陈丽梅. 土壤有益细菌在植物根际竞争定殖的影响因素[J]. 生态学杂志, 2010, 29(6):1235-1239.
[44]van Loon LC. Plant responses to plant growth-promoting bacteria[J]. European Journal of Plant Pathology, 2007, 119:243-254.
[45]Dashti NH, Ali NY, Cherian VM, et al. Application of plant growthpromoting rhizobacteria(PGPR)in combination with a mild strain of cucumber virus(CMV)associated with viral satelliteRNAs to enhance growth and protection against a virulent strain of CMV in tomato[J]. Canadian Journal of Plant Pathology-Revue Canadienne de Phytopathologie, 2012, 34(2):177-186.
[46]Krzyanowska D, Obuchowski M, Bikowski M, et al. Colonization of potato rhizosphere by GFP-Tagged Bacillus subtilis MB73/2, Pseudomonas sp. P482 and Ochrobactrum sp. A44 shown on large sections of roots using enrichment sample preparation and confocal laser scanning microscopy[J]. Sensors, 2012, 12(12):17608-17619.
[47]Teixeira LC, Peixoto RS, Cury JC, et al. Bacterial diversity in rhizosphere soil from Antarctic vascular plants of Admiralty Bay, maritime Antarctica[J]. The ISME Journal, 2010, 4(8):989-1001.
[48]Qiu MH, Zhang RF, Xue C, et al. Application of bio-organic fertilizer can control Fusarium wilt of cucumber plants by regulating microbial community of rhizosphere soil[J]. Biology and Fertility of Soils, 2012, 48(7):807-816.
[49]Allen HK, Moe LA, Rodbumrer J, et al. Functional metagenomics reveals diverse beta-lactamases in a remote Alaskan soil[J]. ISME J, 2009, 3(2):243-251.
[50]Dong L, Meng Y, Wang J, et al. Evaluation of droplet digital PCR for characterizing plasmid reference material used for quantifying ammonia oxidizers and denitrifiers[J]. Analytical and Bioanalytical Chemistry, 2014, 406(6):1701-1712.
[51]Rosch C, Mergel A, Bothe H. Biodiversity of denitrifying and dinitrogen-fixing bacteria in an acid forest soil[J]. Applied Environmental Microbiology, 2002, 68(8):3818-3829.
[52]Zhang N, Yang DQ, Wang DD, et al. Whole transcriptomic analysis of the plant-beneficial rhizobacterium Bacillus amyloliquefaciens SQR9 during enhanced biofilm formation regulated by maize root exudates[J]. BMC Genomics, 2015, 16:685.
(责任编辑 狄艳红)
Research Advances on Colonization of Plant Growth-promoting Rhizobacteria
SUN Zhen1ZHENG Liang2QIU Hao-bin3
(1. Hefei University of Technology,Xuancheng 242000;2. East China Sea Fisheries Research Institute,Chinese Academy of Fishery Sciences,Shanghai 200090;3. College of Engineering and Management,Pingxiang University,Pingxiang 337000)
The PGPB(plant growth-promoting bacterium)presents a promising prospect in agriculture due to its advantages such as inhibiting rhizosphere pathogens,promoting plant growth,and increasing crop production. This paper first summarizes the species of plant growth-promoting rhizobacteria and the promoting mechanisms of involved colonization,and demonstrates the colonization process of rhizobacteria as well as the factors influencing the colonization of bacteria in rhizosphere. Then the paper reviews the detection methods of microorganisms in the colonization of bacteria,including antibiotics marking,immunological methods,and exogenous gene marking methods. Further,the paper discusses high-throughput sequencing technologies widely used in the rhizosphere colonization. Finally,it is concluded that the future researches on interpreting strains in genetic level,gaining microbial community diversity in plant rhizosphere and the functional genes should be of significance to better predict the interaction of PGPB and its host plant,and to apply bio-fertilizers in field.
colonization;rhizobacteria;growth-promoting;high-throughput sequencing
10.13560/j.cnki.biotech.bull.1985.2017.02.002
2016-05-11
中央高校基本科研业务费专项资金资助项目(JZ2015HGBZ0527),合肥工业大学2016年度“学术新人提升A计划”项目(JZ2016HGTA0682)
孙真,女,博士,研究方向:微生物生态;E-mail:sunzhen@hfut.edu.cn
猜你喜欢
昆明医科大学学报(2022年2期)2022-03-29 00:51:58
中国土壤与肥料(2021年5期)2021-12-02 01:06:26
食品安全导刊(2021年20期)2021-08-30 06:40:50
天然产物研究与开发(2018年3期)2018-05-07 06:38:35
山东农业科学(2017年11期)2017-12-26 02:14:06
农业与技术(2017年17期)2017-09-30 19:27:54
热带农业科学(2017年8期)2017-09-18 23:47:33
江苏农业科学(2017年7期)2017-05-23 08:46:38
中国蔬菜(2016年8期)2017-01-15 14:23:38
水生生物学报(2015年1期)2015-02-28 16:01:05
