
乳腺癌中BRCA通路相关基因的研究进展
2017-02-20 05:10刘伟玲郭永军郑州大学附属肿瘤医院分子病理科河南郑州450003
河南医学研究 2017年1期
刘伟玲 郭永军(郑州大学附属肿瘤医院 分子病理科 河南 郑州 450003)
乳腺癌中BRCA通路相关基因的研究进展
刘伟玲 郭永军
(郑州大学附属肿瘤医院 分子病理科 河南 郑州 450003)
BRCA;乳腺癌;FANCD2;PTEN;CHEK2
近年来乳腺癌是女性癌症中最常见和死亡率最高的恶性肿瘤[1]。乳腺癌易感基因BRCA1和BRCA2是与乳腺癌发生有关的重要的抑癌基因,虽然散发性乳腺癌中BRCA1和BRCA2基因突变率不高,但携带BRCA1和BRCA2基因突变的女性发生乳腺癌的概率却较高。BRCA基因已成为遗传性乳腺癌的筛查基因,此外也存在其他易感基因与乳腺癌的发生相关。二代测序的发展使多基因检测成为可能,本文就BRCA基因及其相互作用的乳腺癌易感基因的最新研究进展做相关综述,为乳腺癌的预防与治疗提供理论依据。
BRCA1基因由1990年Hall等[2]发现,定位于17 q21,由24个外显子组成,编码1 863个氨基酸。BRCA1具有转录调控活性及DNA损伤修复功能,参与细胞周期调控,DNA 双链损伤修复,基因的转录调节,细胞凋亡和泛素化等重要的细胞活动。在BRCA1基因中与DNA修复有关的结构域有3个,主要包括:①N-端的锌指结构域;②中间段Rad51结合区;③C端BRCT基序,3个区域任一区域出现突变均可使其基因产生功能,即产生BRCA1基因突变(见图1)。BRCA2基因在1994年由Wooster等[3]发现,定位于13号染色体q12~q13,由27个外显子组成,编码3 418个氨基酸,其基因序列与BRCA1无明显关系。BRCA2基因中与DNA修复有关的结构域主要为Rad51结合区(见图1)。BRCA2的功能主要是通过同源重组和同源重组为基础的DNA断链修复,并且还在细胞生长和DNA损伤监测点控制起作用。
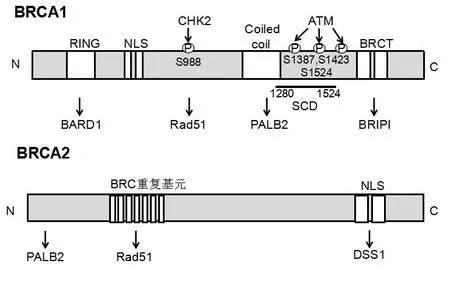
图1 BRCA1和BRCA2基因的结构
研究发现,BRCA1和BRCA2基因突变携带者有很高的乳腺癌发病风险。国内数据显示,BRCA1和BRCA2基因突变携带者70岁时单侧乳腺癌的累积发病风险分别为67.2%和76.8%。与BRCA1不同的是,BRCA2基因突变70岁后乳腺癌的发病率继续增加,到80岁时达93.1%。此外,BRCA1和BRCA2基因突变携带者对侧乳腺癌10 a和20 a的累积发病率分别为19.4%和50.3%[4]。而在西方人口中BRCA1和BRCA2基因携带者70岁时乳腺癌发病风险也明显增加,分别为35%和44%[5]。BRCA基因突变携带的高致癌风险,使得BRCA基因突变的研究更有意义。
1 BRCA通路相关基因
BRCA基因突变的乳腺癌中,除了BRCA1和BRCA2基因突变外,也有其他基因参与,不仅影响BRCA蛋白的表达,而且也影响了乳腺癌的预后。这里主要对乳腺癌FA/BRCA通路和PI3K/AKT通路对BRCA相互作用的基因及其作用机制加以阐述。
1.1 FA/BRCA通路 范可尼贫血(Fanconi anemia,FA)是一种常染色体或X染色体连锁隐性遗传病,表现为骨髓造血功能障碍,常伴发较高的癌症易感性,主要为急性髓性白血病及各种实体性肿瘤。目前发现FA家族有15个基因亚型(FANCA、FANCB、FANCC、FANCD1/BRCA2、FANCD2、FANCE、FANCF、FANCG、FANCI、FANCJ/BRIP1、FANCL、FANCM、FANCN/PALB2、FANCO/RAD51C、FANCP/SLX42),而这15个基因所编码的蛋白均参与同一条细胞信号转导通路,此通路被命名为“范可尼贫血症信号通路”(见图2)。正常细胞中FA/BRCA通路处于休眠状态,而在发生DNA 损伤时该通路被激活。DNA损伤后,由8种基因蛋白产物(FANC-A、B、C、E、F、G、L、M)组成FA复合体单泛素化FANCD2和FANCI构成的ID复合物,ID复合物激活后又被招募到DNA损伤位点,与下游蛋白PLAB2、BRCA2、BRIP1以及BRCA1、RAD51等多种DNA损伤反应蛋白相互作用,从而通过FA途径修复DNA损伤。
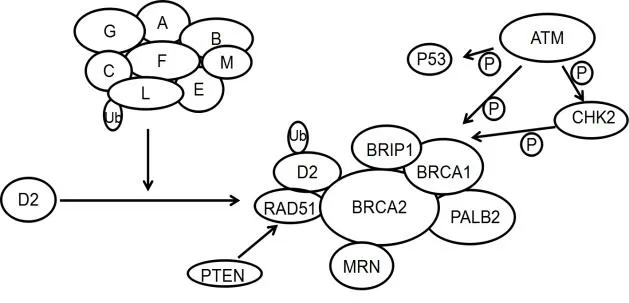
图2 FA/BRCA信号通路作用模式图
1.1.1 FANCD2 FANCD2是组成ID复合物的关键成分,FANCD2的泛素化是FA/BRCA通路的关键步骤,FANCD2泛素化后,在核内靶向与下游蛋白相互作用,促进乳腺癌的形成。FANCF作为一种衔接蛋白,其N端FANCC/FANCE亚单位相互作用,C端与FANCA/FANCG亚单位相互作用,发挥FA复合体的作用,可影响FANCD2的泛素化过程。研究发现,通过沉默FANCF基因,使得FANCD2表达降低,FA/BRCA通路失活,结果显示乳腺癌细胞生长抑制、侵袭力下降,而细胞对化疗药物米托蒽醌、阿霉素、丝裂霉素的敏感性增加[6]。但研究也发现[7],在家族性乳腺癌中BRCA1蛋白的表达与FANCD2蛋白的表达差异有统计学意义,而散发性乳腺癌中,两种蛋白表达差异无统计学意义。未来FANCD2可作为一种靶点来改变FA/BRCA通路,从而有效治疗BRCA基因突变型乳腺癌。
1.1.2 FANCN(PALB2) FANCN/PALB2(BRCA2的定位和伴随蛋白)是通过与BRCA2的相互作用而被发现的,因其二等位基因的截断突变可导致新型的范可尼贫血(FA-N)。DNA损伤后BRCA1召集PALB2,PALB2再通过与BRCA2的相互作用促进BRCA2以及RAD51在DNA损伤点的积累,促进同源重组修复过程。在遗传性乳腺癌中PALB2基因突变率为5.5%,甚至高于BRCA2基因突变率,而PALB2基因突变携带者发生乳腺癌的风险为33%~58%,提示PALB2可作为BRCA1/2之外的新型筛查基因[8]。研究发现,PALB2基因突变能导致其与BRCA2的结合作用丧失,应用PARP抑制剂能导致合成致死性[9]。
1.1.3 FANCJ(BRIP1) FANCJ/BRIP1定位于17号染色体,紧邻BRCA1(17q21),基因编码一种新型蛋白BACH1,可以和BRCA1中两个BRCT结构域相互作用。BACH1属于DEAH DNA解螺旋酶家族成员,BACH1和BRCT突变可以抑制BRCA1的DNA断裂修复过程。研究发现,BRIP1突变能导致乳腺癌的发生,BRIP1的高突变率使得BRIP1成为一种新型的靶点,BRIP1抑制剂用于乳腺癌的治疗仍处于研究中[10]。
1.1.4 CHEK2 细胞周期检测点激酶 2(cell cycle checkpoint kinase 2,CHK2或CHEK2)是一种抑癌基因,编码的蛋白为Chk2激酶,其结构特异性区域包含丝氨酸-谷氨酸/丝氨酸-谷氨酰胺氨基酸对(SQ/TQ)富含区,叉头相关结构域(FHA)和靠近C端的蛋白激酶结构域(KD)。DNA损伤诱导ATM基因磷酸化后,进一步激活CHK2,与CHK2激酶N端的SQ/TQ区相互作用,使一系列位点磷酸化,包括BRCA1的Ser988,使细胞周期进程发生阻滞,同时激活修复基因的转录,促进细胞对损伤进行修复。此外,BRCA1能够维持染色体的稳定性,然而只有CHK2介导的BRCA1磷酸化才能保证有丝分裂纺锤体准确的形成。CHK2的缺失会影响BRCA1对微管着丝粒的形成作用,导致染色体不稳定[11]。CHK2在有丝分裂的这种功能为癌症治疗提供了新的观点。研究发现,CHK2抑制剂AZD7762联合顺铂治疗,有效抑制了肿瘤细胞的生长[12]。
1.1.5 BARD1 BARD1是1996年由研究人员发现和BRCA1具有相似的环指功能域,可以和BRCA1蛋白结合,并因此而命名。BARD1通过其环指区和BRCA1氨基末端的环指区相互结合,并增强BRCA1的E3连接酶活性。BARD1和BRCA1在多数组织中共表达,在小鼠中两者基因的敲除有相似的表型,BARD1和BRCA1形成的这种稳定的复合体,由此猜想BARD1在BRCA1基因的表达过程中扮演着辅助角色[13]。此外,BARD1在散发性乳腺癌以及BRCA1突变的家族性肿瘤中突变并过度表达[14]。当BRCA1和BARD1的基因产物数量相称时,诱导细胞存活和DNA的修复,如果BARD1产物超过BRCA1则诱导凋亡[15]。然而在BARD1突变和乳腺癌的风险评估的Meta分析中,并没有发现BARD1突变能增加乳腺癌的风险[16],具体作用机制需要进一步研究。
1.1.6 RAD51 DNA修复蛋白Rad51是由1993年首次被Shinohara等[17]克隆,是一种DNA依赖性ATP酶。研究者把Rad51、BRCA1和BRCA2作为一个复合体研究其对乳腺癌的预后和预测的作用,发现在早期乳腺癌中Rad51/BRCA1/BRCA2复合体低表达的乳腺癌预后差,但对放疗敏感。Jakubowska等[18]已证实,Rad51单核苷酸多态135G/C是BRCA1/2突变乳腺癌的重要调节因素。Rad51可通过BRCA1使细胞周期停滞在某一特定阶段,再进行DNA修复。BRCA2则通过与Rad51结合,对Rad51进行功能调控。研究表明,RAD51单核苷酸多态可增加BRCA2突变的携带者的致病率,而对BRCA1突变者或非突变者无影响[19]。具体机制是复杂的,有待进一步研究。
1.2 PI3K/AKT通路
1.2.1 ATM ATM是引起毛细血管扩张性共济失调症(ataxinet-lanigestia,AT)的常染色体隐性突变基因。由Savitsky等[20]在1995年通过原位克隆技术发现。ATM属于磷脂酰肌醇酶(PI3K)相关的蛋白家族的成员。DNA损伤后,ATM受刺激而活化,催化BRCA1基因的磷酸化,进一步激活下游通路,引起一系列的级联反应。研究发现,ATM缺失表达是乳腺上皮细胞恶性转化的一个早期事件,随着乳腺癌进展,ATM缺失也随之进展[21]。ATM抑制剂也能对于BRCA1缺陷的细胞通过累积的DNA双链断裂产生合成致死性,为BRCA1突变型的乳腺癌提供了一种新型疗法[22]。
1.2.2 PTEN PTEN是一种抑癌基因,DNA损伤后,PTEN作用于染色体并上调RAD51基因的表达,减少双链断裂的发生率。PTEN在PI3K/AKT信号通路起着重要作用,PTEN基因突变,能激活PI3K/AKT通路导致细胞增殖及分化的紊乱,促进肿瘤的发生。此外,BRCA1缺陷也能够激活AKT信号通路[23]。BRCA1和PTEN对PI3K/AKT信号通路具有类似的抑制作用,而两者之间的相关作用机制需要我们进一步研究。研究发现,一些带有BRCA1突变的乳腺癌有较高的PTEN突变率[24]。Martins等[25]猜测在BRCA1相关性乳腺癌中,PTEN是促进乳腺癌进展的初始因素。PTEN和BRCA1在PI3K/AKT通路类似的作用,以及乳腺癌中两种基因共突变的发生,未来联合应用BRCA1和PTEN抑制剂将很有可能更有效促进肿瘤的治疗。
1.2.3P53P53是一种抑癌基因,在细胞周期调控和维持基因组稳定性具有重要作用,也被称为“基因管家”。BRCA1可直接与P53结合,激活P21、Gadd45等下游靶基因,增加DNA修复能力。而且P53也可以调控BRCA2转录[26]。研究发现P53基因突变患者对EC(表阿霉素+环磷酰胺)、TC(紫杉醇+环磷酰胺)及TAC(多西他赛+阿霉素+环磷酰胺)方案新辅助化疗并不敏感,未突变患者则对TAC方案新辅助化疗有较高的敏感性,可能为指导实际临床用药提供一定的理论依据[27]。
1.3 其他 研究表明,除了上述基因在乳腺癌的发生中可能对BRCA1和BRCA2基因产生影响,还有其他基因也可能产生影响,如由MRE11、NBS1、RAD50组成的MRN复合物及OGG1基因也会与BRCA1和BRCA2基因发生作用[28-29]。具体机制有待进一步研究。
2 结语
BRCA1和BRCA2在乳腺癌的发生中和多种基因发生关联,共同促进乳腺癌的发生,基因的发展是复杂的,但上述基因和BRCA基因相互作用都是直接或间接通过FA/BRCA通路,或者通过影响PI3K/AKT通路发挥作用,提示我们需更加深入研究上述两个通路的具体作用机制。二代测序、PARP抑制剂及PI3K抑制剂的发展给BRCA突变乳腺癌的治疗带来了新的进展。我们只有深入了解BRCA基因通路各种基因的相互作用,找到更多的基因靶点,才能为BRCA基因突变型乳腺癌的靶向治疗提供更多的研究方向。
[1] Chen W,Zheng R,Baade P D,et al.Cancer statistics in China,2015[J]. CA Cancer J Clin,2016,66(2):115-132.
[2] Hall J M,Lee M K,Newman B,et al.Linkage of early-onset familial breast cancer to chromosome 17q21[J].Science,1990,250(4988):1684-1689.
[3] Wooster R,Neuhausen S L,Mangion J,et al.Localization of a breast cancer susceptibility gene,BRCA2,to chromosome 13q12-13[J].Science,1994,265(5181):2088-2090.
[4] 杨晓晨,胡震,吴炅,等.中国汉族人群中BRCA1和BRCA2基因突变携带者患乳腺癌风险的研究[J].中国癌症杂志,2015,(4):247-252.
[5] Nielsen H R,Nilbert M,Petersen J,et al.BRCA1/BRCA2 founder mutations and cancer risks: impact in the western Danish population[J]. Fam Cancer,2016,15(4):507-512.
[6] Yu J,Zhao L,Li Y,et al.Silencing of fanconi anemia complementation group f exhibits potent chemosensitization of mitomycin C activity in breast cancer cells[J].J Breast Cancer,2013,16(3):291-299.
[7] 冯亮,李兴东,陈浩,等.家族性与散发性乳腺癌BRCA1与FANCD2基因相关性的研究[J].中华肿瘤防治杂志,2011,(22):1752-1754.
[8] Janatova M,Borecka M,Soukupova J,et al.PALB2 as another candidate gene for genetic testing in patients with hereditary breast cancer in Czech republic.[J].Klin Onkol,2016,29(Suppl 1):S31-S34.
[9] Smith M A,Hampton O A,Reynolds C P,et al.Initial testing (stage 1) of the PARP inhibitor BMN 673 by the pediatric preclinical testing program:PALB2 mutation predicts exceptional in vivo response to BMN 673[J].Pediatr Blood Cancer,2015,62(1):91-98.
[10]Ouhtit A,Gupta I,Shaikh Z.BRIP1,a potential candidate gene in development of non-BRCA1/2 breast cancer[J].Front Biosci (Elite Ed),2016,(8):289-298.
[11]Stolz A,Ertych N,Bastians H.Tumor suppressor CHK2:regulator of DNA damage response and mediator of chromosomal stability[J].Clin Cancer Res,2011,17(3):401-405.
[12]Itamochi H,Nishimura M,Oumi N,et al.Checkpoint kinase inhibitor AZD7762 overcomes cisplatin resistance in clear cell carcinoma of the ovary[J].Int J Gynecol Cancer,2014,24(1):61-69.
[13]Irminger-Finger I,Ratajska M,Pilyugin M.New concepts on BARD1:Regulator of BRCA pathways and beyond[J].Int J Biochem Cell Biol,2015,72:1-17.
[14]Wu J Y,Vlastos A T,Pelte M F,et al.Aberrant expression of BARD1 in breast and ovarian cancers with poor prognosis.[J].Int J Cancer,2006,118(5):1215-1226.
[15]Feki A,Jefford C E,Berardi P,et al.BARD1 induces apoptosis by catalysing phosphorylation of p53 by DNA-damage response kinase[J].Oncogene,2005,24(23):3726-3736.
[16]Ding D P,Zhang Y,Ma W L,et al.Lack of association between BARD1 Cys557Ser variant and breast cancer risk:a meta-analysis of 11,870 cases and 7,687 controls[J].J Cancer Res Clin Oncol,2011,137(10):1463-1468.
[17]Shinohara A,Ogawa H,Matsuda Y,et al.Cloning of human,mouse and fission yeast recombination genes homologous to RAD51 and recA[J]. Nat Genet,1993,4(3):239-243.
[18]Jakubowska A,Gronwald J,Menkiszak J,et al.The RAD51 135 G>C polymorphism modifies breast cancer and ovarian cancer risk in Polish BRCA1 mutation carriers[J].Cancer Epidemiol Biomarkers Prev,2007,16(2):270-275.
[19]Kadouri L,Kote-Jarai Z,Hubert A,et al.A single-nucleotide polymorphism in the RAD51 gene modifies breast cancer risk in BRCA2 carriers,but not in BRCA1 carriers or noncarriers[J].Br J Cancer,2004,90(10):2002-2005.
[20]Savitsky K,Bar-Shira A,Gilad S,et al.A single ataxia telangiectasia gene with a product similar to PI-3 kinase[J].Science,1995,268(5218):1749-1753.
[21]Shiloh Y,Ziv Y.The ATM protein kinase:regulating the cellular response to genotoxic stress,and more[J].Nat Rev Mol Cell Biol,2013,14(4):197-210.
[22]Albarakati N,Abdel-Fatah T M,Doherty R,et al.Targeting BRCA1-BER deficient breast cancer by ATM or DNA-PKcs blockade either alone or in combination with cisplatin for personalized therapy[J].Mol Oncol,2015,9(1):204-217.
[23]Xiang T,Ohashi A,Huang Y,et al.Negative Regulation of AKT Activation by BRCA1[J].Cancer Res,2008,68(24):10040-10044.
[24]Saal L H,Gruvberger-Saal S K,Persson C,et al.Recurrent gross mutations of the PTEN tumor suppressor gene in breast cancers with deficient DSB repair[J].Nat Genet,2008,40(1):102-107.
[25]Martins F C,De S,Almendro V,et al.Evolutionary pathways in BRCA1-associated breast tumors[J].Cancer Discov,2012,2(6):503-511.
[26]Verma S,Rao B J.p53 suppresses BRCA2-stimulated ATPase and strand exchange functions of human RAD51[J].J Biochem,2013,154(3):237-248.
[27]张丝媛.p53基因突变与乳腺癌新辅助化疗疗效关系的研究[D].长沙:中南大学,2012.
[28]Damiola F,Pertesi M,Oliver J,et al.Rare key functional domain missense substitutions in MRE11A,RAD50,and NBN contribute to breast cancer susceptibility:results from a Breast Cancer Family Registry case-control mutation-screening study[J].Breast Cancer Res,2014,16(3):R58.
[29]Benitez-Buelga C,Vaclova T,Ferreira S,et al.Molecular insights into the OGG1 gene,a cancer risk modifier in BRCA1 and BRCA2 mutations carriers[J].Oncotarget,2016,7(18):25815-25825.
·经验交流·
R 655.8
10.3969/j.issn.1004-437X.2017.01.024
2016-04-22)
猜你喜欢
现代临床医学(2022年4期)2022-09-29
湖北农业科学(2022年11期)2022-07-18
中老年保健(2021年12期)2021-08-24
中国生殖健康(2020年2期)2021-01-18
医学与法学(2020年3期)2020-09-18
生物信息学(2020年1期)2020-05-16
小学生导刊(2018年13期)2018-06-29
中国生殖健康(2018年2期)2018-01-12
故事作文·高年级(2017年3期)2017-04-12
天津科技大学学报(2016年1期)2016-02-28
