
ω-3多不饱和脂肪酸在养猪生产中的研究进展
2017-02-10 02:02:51胡金杰吴志锋
粮食与饲料工业 2017年1期
胡金杰,曹 霞,吴志锋
(湖南九鼎科技(集团)有限公司技术中心,湖南 长沙 410003)
ω-3多不饱和脂肪酸在养猪生产中的研究进展
胡金杰,曹 霞,吴志锋
(湖南九鼎科技(集团)有限公司技术中心,湖南 长沙 410003)
ω-3多不饱和脂肪酸(ω-3 PUFAs)是机体不可缺少的一种必需脂肪酸和重要的能量来源,具有多种生理功能。介绍了畜牧业生产中富含ω-3 PUFAs的常见原料及含量,同时对ω-3 PUFAs在提高猪只免疫功能和改善猪肉品质、母猪生产性能、公猪精液品质方面的应用研究进行了综述,为ω-3 PUFAs在养猪生产中的进一步推广和应用提供参考。
ω-3 多不饱和脂肪酸;猪;必需脂肪酸;生理功能;能量来源
多不饱和脂肪酸(PUFA)营养功能以及开发利用成为近几年国际研究的热点。多不饱和脂肪酸之所以受到广泛关注,不仅因为它们很多是机体不可缺少的必需脂肪酸,是体内重要的能量来源,更重要的是因为它们对机体的多种生理功能,包括调节脂质代谢、调节机体免疫功能、抵抗癌症、影响细胞膜信息传递途径和转录因子活化途径等一些代谢相关酶基因的表达等方面。而ω-3多不饱和脂肪酸(ω-3 PUFAs)是机体不可缺少的必需脂肪酸之一,具有降低血液胆固醇,降低血栓形成,减少心脑血管病的发生,抗癌,提高机体免疫功能等[1]。由于ω-3 PUFAs在人体健康方面的重要作用,研究人员利用其特殊的营养作用在各阶段猪饲粮中添加,预期得到更好的经济效益。
1 ω-3多不饱和脂肪酸的来源及含量
ω-3 PUFAs属于长链多不饱和脂肪酸,第一个双键在三、四两个碳原子之间,主要包括α-亚麻酸(ALA),二十碳五烯酸(EPA),二十二碳五烯酸(DPA)和二十二碳六烯酸(DHA),机体可利用ALA为前体在Δ5、Δ6去饱和酶的作用下合成EPA和DHA。ω-3 PUFAs的来源十分广泛,富含ω-3 PUFAs的原料可分为植物性、动物性和微藻类等三大类。植物性原料主要有亚麻籽、大豆、菜籽、紫苏籽、大麻籽、紫花苜蓿粉、马齿苋及其榨出的植物油。ALA主要来源于植物油,常用油种中以亚麻籽油种含量最高,其次是豆油、菜籽油[2]。亚麻籽中ALA质量分数在12%左右,是生产富含ω-3 PUFAs畜禽饲料的常用原料之一[3]。紫苏籽含油35%~51%,其中ALA质量分数为63%~70%;大豆油、菜籽油、核桃油ALA质量分数大约为10%[4]。EPA和DHA的来源主要是鱼类、海洋真菌和微藻。动物性原料主要是各种鱼类提炼出的鱼油,鱼油中含25%~59%的ω-3 PUFAs,主要是DHA和EPA[5]。鲱鱼油中EPA和DHA分别占总脂肪酸的16.03%和10.83%[6]。研究认为微生物作为海洋食物链中的初级生产者,是ω-3 PUFAs的原始生产者[7]。目前已分离出的富含DHA和EPA的海洋微生物,主要是一些低等的海洋真菌和微藻,包括金藻纲、黄藻纲、硅藻纲、红藻纲、绿藻纲和隐藻纲中都有富含EPA的藻类。其中有些微藻所含的EPA占细胞总脂的40%以上[8]。而富含DHA的海洋藻类相对较少,主要集中在甲藻、金藻和硅藻中,目前商业上用于生产DHA的海洋微生物主要有裂殖壶菌和隐甲藻菌[9]。已有研究证明包括小球藻属、螺旋藻属、紫球藻属等微藻类,富含不饱和脂肪酸,可用于畜禽饲料[10]。在畜牧业生产中一些含有ω-3 PUFAs成分的常见原料见表1。
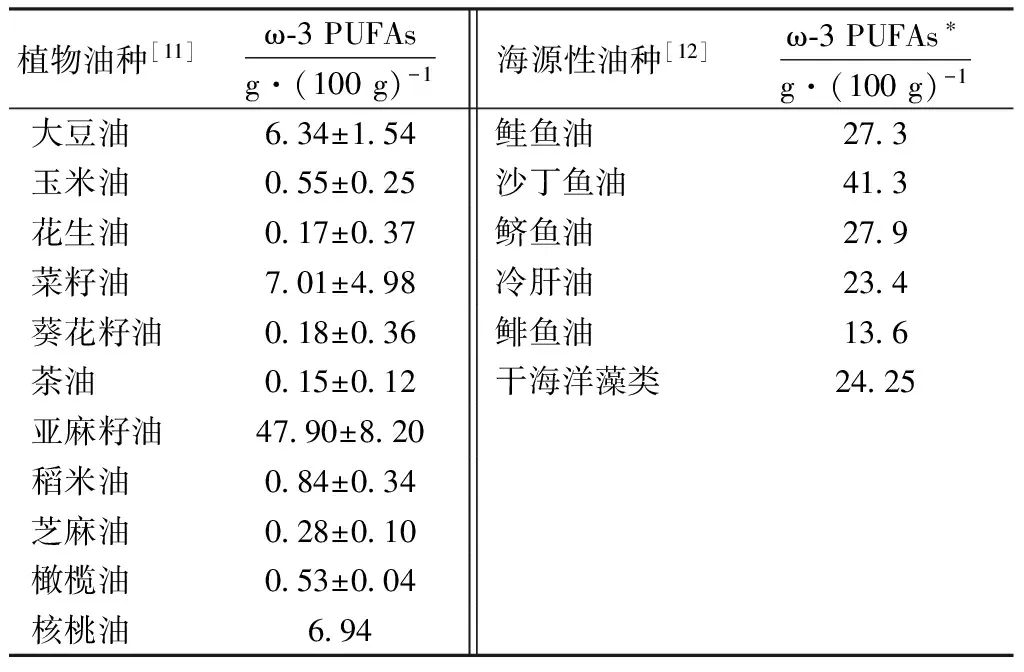
表1 常用油脂中ω-3 PUFAs的含量
注:*海源性油种中ω-3 PUFAs为ALA、花生四烯酸(AA)、EPA和DHA的总和。
2 ω-3多不饱和脂肪酸在养猪生产中的研究进展
2.1 ω-3多不饱和脂肪酸与猪只免疫功能
在养猪生产中,断奶仔猪因断奶失去了母乳中免疫球蛋白的供给以及断奶应激而导致未发育完全的肠道免疫系统应激很大,导致仔猪腹泻。仔猪刚断奶且采食固体日粮,处于较大的断奶应激和免疫应激条件下,任何一种应激(如环境、免疫或疾病侵袭)都会影响动物的生长。脂类的免疫作用主要是与细胞中脂类成分和二十烷类的合成有关。所有的免疫活性细胞都可利用PUFA合成二十烷类,多数二十烷类具有免疫调节功能。如花生四烯酸产生的类二十烷酸(如PGE2)是重要的炎性介质,参与炎症反应,而ω-3 PUFAs可降低这类介质的产生[13]。不同ω-3 PUFAs发挥不同的免疫调节作用,EPA比DHA的作用更广泛、更强,低水平的EPA就足以影响免疫反应[14]。ω-3 PUFAs能降低细胞因子的分泌,因此能够减轻一些自身免疫疾病、炎症等的恶化程度[15],还能抑制抗原递呈细胞发挥抗原递呈作用而抑制细胞免疫[16]。另外,ω-3 PUFAs还具有改变补体和免疫细胞功能的作用。刘玉兰和Liu等[17,18]发现,EPA和DHA剂量依赖性地抑制了断奶仔猪外周血淋巴细胞增殖,抑制白细胞介素-2(IL-2)的产生。Gaines等[19]发现,日粮添加鱼油能够抑制多糖(LPS)对断奶仔猪的刺激,抑制肿瘤坏死因子 -a(TNF-a)的分泌,阻止LPS对仔猪的生长抑制作用。日粮中添加富含ω-3 PUFAs的鱼油能够提高断奶仔猪日增重和饲料转化效率[17]。左磊等[20]研究发现,ω-3 PUFAs和ω-6 PUFAs比例不平衡时,ω-3 PUFAs含量高的免疫抑制作用要强于ω-6 PUFAs。ω-3/ω-6 PUFAs比值为10时,白细胞介素-1β (IL-1β)、白细胞介素-6(IL-6)和TNF-α在血浆中的含量达最高,同时仔猪的日增重和饲料转化效率最高。Upadhaya等[21]通过给平均体重为50.7 kg的生长肥育猪试验期6周,每周肌肉注射两次大肠杆菌脂LPS对猪进行免疫刺激,通过对照组,300 IU维生素E组和作为ω-3 PUFAs来源的亚麻籽油替代25%的动物油脂三组进行试验,研究发现,添加ω-3 PUFAs组比对照组中血液白细胞数量显著降低,维生素E组和ω-3 PUFAs组猪血浆中皮质醇和TNF-a也显著降低。身体对严重的免疫系统刺激的反应是增加促炎性细胞因子和PGE2的分泌,而ω-3 PUFAs能降低受免疫刺激的生长肥育猪TNF-a和PGE2的生成。LPS对猪只进行免疫刺激属于免疫应激,而免疫应激是畜禽的免疫系统不正常激活所致,也属于一种炎症反应。但也有研究指出,鱼油对炎性细胞因子的产生没有影响,产生这种不一致的原因可能是因为ω-3 PUFAs添加剂量不同导致的[22]。所以ω-3 PUFAs对急、慢性炎症具有良好的预防和治疗效果,对猪只免疫应激具有一定的缓解作用,但是,还要进一步确定ω-3 PUFAs在不同阶段猪饲粮的适宜添加量和适宜添加时间等,以发挥最大的抗炎症和抗免疫应激作用。
2.2 ω-3多不饱和脂肪酸与猪肉品质
随着生活水平的提高,人们对肉质的要求越来越高,目前大多肉制品富含饱和脂肪酸,相对缺乏多不饱和脂肪酸,而研究表明,由于PUFA经消化道吸收后不经氢化,直接合成胴体脂肪,因此猪肉中的脂肪酸类型受日粮脂肪酸类型的影响很大。Karin等[23]给猪饲喂含5%亚麻籽油日粮后发现,猪的所有组织中ω-3 PUFAs表现为增长的趋势,猪肌肉脂质组织中ALA含量显著增加到13.2%,随着ω-3 PUFAs的富集,肌肉中花生四烯酸(AA)含量降低,ω-6/ω-3 PUFAs比值降低。刘则学[24]在生长肥育猪屠宰前0、30、60和90 d饲粮中添加10%亚麻籽,发现猪肌内脂肪和皮下脂肪中ω-3 PUFAs含量随着亚麻籽添加时间的延长而极显著的增加,添加90 d时ω-3 PUFAs在100 g肌内脂肪中占7.13%,而对照组仅占1.11%。Romans等[25]在屠宰前7、14、21和28 d时在猪饲粮中添加15%的亚麻籽,发现猪肌肉中ω-3 PUFAs含量(0.2~0.6 mg/g)随添加时间长短而发生变化,时间越长含量越高,但添加21和28 d的ω-3 PUFAs含量相似。这一结果与Kouba等得出的结果一致,Kouba等[26]发现在猪饲粮中添加6%压碎的亚麻籽可以降低猪最长肌中ω-6/ω-3 PUFAs的比值,而饲喂60和100 d,ω-6/ω-3 PUFAs的比值分别为3.0和3.1。Sardi等[27]在猪饲粮中不同时期内添加两个不同水平(2.5 g/kg添加8周,2.5 g/kg或5.0 g/kg添加4周)的富含DHA的海水藻产品(17.1 g/kg总FA),结果发现添加海水藻的猪背最长肌DHA含量显著增加;对比添加量和饲喂时间发现,添加量为5.0 g/kg时,背最长肌DHA含量最高,而添加2.5 g/kg饲喂8周与2.5 g/kg或5.0 g/kg饲喂4周肉中DHA含量无显著差异。
上述研究表明,可以通过调整猪日粮中的多不饱和脂肪酸含量来改善肉制品的多不饱和脂肪酸含量和不同多不饱和脂肪酸之间的比值。但是Wood等[28]研究发现肉中脂肪酸影响脂肪组织硬度、货架期和风味。多不饱和脂肪酸含量升高时,肌肉的脂肪随之变软,贮存、加工过程中易氧化腐败,而氧化腐败主要是由于脂质氧化产物与美拉德反应产物的相互作用,导致风味物质的浓度和本质发生改变,动物内源或由饲粮摄取的不饱和脂肪酸比例、活性氧物质产生的数量以及抗氧化产物水平决定了氧化反应的强弱[29]。不饱和脂肪酸分子内含有很多双键,易被氧化成饱和脂肪酸,肌肉易氧化腐败,货架期缩短、产生异味,使肉品质下降,影响经济效益。为了增加猪肉抗氧化能力,很多研究者进而在饲粮中添加抗氧化剂如维生素E和矿物质硒进行研究,Guillevic等[30]在每千克饲粮中添加170 mg维生素E,170 μg亚硒酸盐和80 μg硒-蛋氨酸,还有很多研究在饲粮中添加不同剂量的维生素E(40~200 mg/kg)来增加猪肉的抗氧化能力。Botsoglou等[31]认为饲粮中添加亚麻籽,再加入200 mg/kg的维生素E就足以保护背最长肌免于氧化,能增加组织中α-生育酚水平。而Cannata等[32]研究发现,在添加亚麻籽饲粮中每千克饲粮中增加170 mg维生素E和250 μg硒,饲养出的大肥猪用于制作干腌火腿,火腿的感官品质仍受到影响。近来有研究认为,在猪饲粮中长期添加植物提取物(包括毛蕊花苷)能增加背最长肌维生素E含量和增强抗氧化能力[33]。Mairesse等[34]在含有丰富的ω-3 PUFAs猪饲粮中添加植物抗氧化混合物,能够减少干腌火腿中丙二醛含量,并且不影响其他质量参数。
通过诸多研究发现,在添加亚麻籽或其他来源增加饲粮ω-3 PUFAs时,为了不影响猪肉品质,可添加一定剂量的维生素E、硒和植物提取物来达到保持猪肉抗氧化能力,增加货架期和加工性能。
2.3 ω-3多不饱和脂肪酸与母猪生产
PUFA是脂肪的重要组成成分,同时还广泛参与类固醇激素如雌二醇、睾酮、孕酮和糖皮质激素等的合成,因此不同种类和比例的PUFA对家畜繁殖性能具有重要影响[35]。Moreira等[36]在后备母猪饲粮中添加鱼油(含ω-3 PUFAs 6.88 g/d)与添加豆油的对照组进行比较,发现饲喂45 d后,鱼油组子宫重量稍有增加(P=0.09),并能降低血浆中胆固醇含量(P=0.06)。另外脂肪酸也是卵母细胞成熟过程中的重要能量来源,所以不同类型的脂肪酸对卵母细胞的质量也会产生影响,Spencer等[37]研究发现,在母猪分娩前30 d至分娩间饲粮添加ω-3PUFAs能够增加窝产仔数。Lauridsen等[38]也发现母猪饲粮中添加ω-3 PUFAs对胚胎存活率和窝产仔数的增加有益,这可能是因为ω-3 PUFAs可抑制前列腺素的合成,提高母体对胎儿的保护作用,进而提高胎儿成活率。Papadopoulos等[39]报道称,在分娩前3 d或者8 d提高母猪日粮中ω-3 PUFAs含量,会影响胰岛素和瘦素的分泌,从而提高母猪在哺乳期的采食量和奶水质量来提高哺乳仔猪生长性能。Smit等[40]也研究发现,在初次妊娠的母猪基础饲粮中添加ω-3 PUFAs,母猪每天摄入84 g,试验从妊娠60 d开始至21 d哺乳期结束,结果发现添加ω-3 PUFAs的试验组不影响母猪产仔数和出生体重,而能增加仔猪断奶时体重,之后的研究进一步证实了这一结果,在母猪饲粮中添加0.5%的海源性ω-3 PUFAs可以增加仔猪断奶时体重和日增重。肖成林等[41]在妊娠后期(103 d)母猪饲粮中添加鱼油至仔猪21 d断奶,研究乳汁中脂肪酸含量以及对仔猪生长性能的影响,结果发现,添加5%鱼油极显著增加乳中ω-3 PUFAs含量,极显著降低ω-6 PUFAs含量;与添加7%猪油相比,7%鱼油饲粮极显著提高乳中ω-3 PUFAs含量,提高初生重小于等于1.5 kg仔猪的育成率,提高仔猪21日龄ADG,显著降低初生重小于等于1.2 kg较小仔猪比例。
因此,饲粮中添加ω-3 PUFAs的饲料原料能提高母猪繁殖性能,改善乳汁成分,提高仔猪生长性能等。其作用机理可能是日粮中添加ω-3 PUFAs会影响母体的激素代谢[42],进而通过乳汁,改善哺乳仔猪的免疫状态和生长性能[43]。
2.4 ω-3多不饱和脂肪酸与公猪精液品质
精液品质包括射精量,精子浓度、精子活力、畸形率,在规模化养猪生产中,公猪的精液品质直接影响母猪的繁殖性能,影响着猪场的生产成绩。在哺乳动物精子的脂肪酸组成中,多不饱和脂肪酸占比例较高,而在公猪、公牛、公羊等哺乳动物精子的多不饱和脂肪中DHA含量最高[44]。研究发现,在公猪饲粮中添加鱼油可以提高睾丸中卵磷脂、磷脂酰乙醇胺含量和鞘磷脂中ω-3 PUFAs含量以及DHA与DPA的比值,降低鞘磷脂中ω-6/ω-3 PUFAs的比值,但猪吸收饲粮中添加的DHA存在一个饱和剂量,超过了则可能不会被吸收[45]。Liu等[46]在长白公猪饲料中添加不同比例ω-6/ω-3 PUFAs (14.4、6.6和2.2),结果发现ω-6/ω-3 PUFAs比值为6.6和2.2饲粮可以有效增加精子中DHA和ω-3 PUFAs含量,降低DPA含量和ω-6/ω-3 PUFAs比值;6.6比值饲粮可以显著促进精子活力,并能增强公猪血浆中超氧化物歧化酶(SOD)和总抗氧化能力(TAC)。Estienne等[47]在公猪饲粮中添加脂肪酸93 g/d ω-3 PUFAs,发现在试验的第9~16 周,精子数量和密度均显著高于试验第1~8 周,但是与对照组相比各项指标均有所提高,但未达到显著差异。Yeste等[48]在公猪饲粮中添加300 g/d鱼油(含91.44 g ω-3 PUFAs),饲喂26 周,结果并未发现公猪射精量、精子密度等方面有差异,也并未发现精子活力存在差异,但使大约克和皮特兰公猪精子畸形率显著降低,杜洛克公猪精子的畸形率则无显著变化。
所以,目前饲粮中添加不同剂量的ω-3 PUFAs对公猪精子数量、密度、畸形率的影响研究结果并不一致,这可能与ω-3 PUFAs的来源不同有关,而且ω-6/ω-3 PUFAs比值不同对公猪精液影响不同,为达到最优效果,不同品种公猪饲粮最佳的ω-6/ω-3 PUFAs比值还需进一步研究。
3 小结
以ω-3 PUFAs为代表的不饱和脂肪酸对人体有重要的生理功能,在脂质代谢、改善免疫力、抵抗癌症以及一些代谢相关酶基因的表达等方面都有调节功能。由于猪肉中的脂肪酸组成受日粮的影响较大,因此通过向日粮中添加富含ω-3 PUFAs等多不饱和脂肪酸可以明显提高猪肉制品中的多不饱和脂肪酸含量以及比例,同时ω-3 PUFAs还具有提高断奶仔猪免疫应激能力、提高母猪生产性能以及公猪精液质量等作用。但由于ω-3 PUFAs等多不饱和脂肪酸也会降低猪肉脂质的抗氧化能力,影响肉品质和货架期,因此一般会通过添加维生素E、硒和植物提取物来提高猪肉抗氧化能力。笔者相信随着关于ω-3 PUFAs的作用机理、适宜添加量等方面研究的继续完善,ω-3 PUFAs在畜牧行业的应用将越加广泛。
[1] 刘庆生,王加启,卜登攀,等.多不饱和脂肪酸对动物血脂代谢与抗氧化性能影响研究进展[J].中国饲料,2009(8):7-10.
[2] 郭金枝,李藏兰.多不饱和脂肪酸在养猪生产中应用的研究进展[J].中国畜牧杂志,2012,48(11):76-78.
[3] 邓兴照.多不饱和脂肪酸日粮对蛋鸡脂类代谢及蛋黄脂肪酸组成的影响[D].杨凌:西北农林科技大学,2005.
[4] 赵春艳,敖长金.多不饱和脂肪酸对机体免疫机能的影响[J].饲料研究,2005,11:33-36.
[5] 郎 婧,石宝明,张宏宇,等.亚麻籽中n-3多不饱和脂肪酸对猪肌体脂肪酸组成和肉品质的影响[J].中国饲料,2010(8):9-12.
[6] 宋 芸,丁华,吴葆杰.n-3多不饱和脂肪酸的研究进展[J].中国海洋药物,2005,24(6):57-60.
[7] 朱路英,张学成,宋晓金等.n-3多不饱和脂肪酸DHA、EPA研究进展[J].海洋科学,2007,31(11):78-85.
[8] SERVEL M O, CLAIRE C, DERRIEN A, et al. Fatty acid composition of some marine microalgae [J]. Phytochemistry, 1994, 36: 691-693.
[9] 朱路英,张学成,宋晓金,等.n-3多不饱和脂肪酸DHA、EPA研究进展[J].海洋科学,2007,11(31):78-64.
[10] LUM K K, KIM J, XIN G L. Dual potential of microalgae as a sustainable biofuel feedstock and animal feed [J]. Journal of Animal Science and Biotechnology, 2013, 4(1):53.
[11] 廖静.我国代表性油种脂肪酸组成分析及热稳定性研究[M].长沙:湖南农业大学.
[12] ROSSI R, PASTORELLI G P, CANNATA S, et al. Recent advances in the use of fatty acids as supplements in pig diets: a review [J].Animal Feed Science and Technology, 2010, 162:1-11.
[13] 刘玉兰,黄晶晶.N-3多不饱和脂肪酸对畜禽免疫、炎症及免疫应激的调节作用[J].畜牧与兽医,2007,39(9):33-35.
[14] 王珊珊,李 秋,徐田彬,等.N-3多不饱和脂肪酸的生理功能特性及应用[J].中国食物与营养,2009,15(10):51-54.
[15] 魏 毅,张秋海.脂肪酸与免疫功能的关系[J].潍坊医学院学报,2005,27(3):217-220.
[16] HUGHUS D A, PINDER A C, CONNOR W E, et al. Supplements n-3 polyunsaturated fatty acids inhibit the antigen- presenting function of human monocytes [J]. Am J Clin Nutr, 2000, 71(1):357S- 360S.
[17] 刘玉兰.鱼油对断奶仔猪抗免疫应激机理研究[D].北京:中国农业大学,2003.
[18] LIU Y L, GONG L M, LI D F, et al. Effects of fish oil only lymphocyte proliferation, cytokine production and intracellular signaling in weanling pigs [J].Arch Anim Nutr, 2003, 57:151-165.
[19] GAUNES A M, CARROLL J A, YI G F, et al. Effect of menhaden fish oil Supplementation and lipopolysaccharide exposure on nursery pigs :II. Effects on the immune axis when fed simple or complex diets containing no spray-dried plasma [J].Domest Anim Endocrinol, 2003,24:353-365.
[20] 左 磊,李藏兰,赖长华.不同n-6/n-3多不饱和脂肪酸比值对断奶仔猪生长性能和免疫反应的影响[J].中国畜牧杂志,2010,46(23):48-50.
[21] UPADHAYA S D, KIM J C, MULLAN B P, et al. Vitamin E and omega-3 fatty acids independently attenuate plasma concentrations of proinflammatory cytokines and prostaglandin E3 in Escherichia coli lipopolysaccharide-challenged growing-finishing pigs [J]. J Anim Sci, 2015,93:2 926-2 934.
[22] 徐晓燕,王加启,卜登攀,等.脂肪酸免疫调节功能研究进展[J].华北农学报,2011,26(S2):239-242.
[23] NUERNBERG K, FISCHER K, NUERNBERG G, et al. Effects of dietary olive and linseed oil on lipid composition,meat quality,sensory characteristics and muscle structure in pigs[J].Meat Science,2005,70:63-74.
[24] 刘则学.亚麻籽中多不饱和脂肪酸在猪不同组织中的富集规律及对猪胴体品质的影响[D].武汉:华中农业大学,2006.
[25] ROMANS J R, JOHNSON R C, WULF D M, et al. Effects of ground flaxseed in swine diets on pig performance and on physical and sensory characteristics and omega-3 fatty acid content of pork: II. Duration of 15% dietary flaxseed[J]. Anim. Sci. 1995,73(7):1 987-1 999.
[26] KOUBA M, ENSER M, WHITTINGTON F M, et al. Effect of a high-linolenic acid diet on lipogenic enzyme activities, fatty acid composition,and meat quality in the growing pig [J]. Journal of Animal Science, 2003, 81(8):1 967-1 979.
[27] SARDI L, MARTELLI G, LAMBERTINI L, et al. Effects of a dietary supplement of DHA-rich marine algae on Italian heavy pig production parameters [J]. Livestock Science, 2006, 103(1-2):95-103.
[28] WOOD J D, RICHARDSON R I, NUTE G R, et al. Effects of fatty acids on meat quality [J]. Meat Science, 2004,66(1):21-32.)
[29] 马燕芬,高 民,卢德勋. 动物产品中多不饱和脂肪酸和其他脂肪酸含量变化及其影响因素[J].动物营养学报,2011, 32(9):1 131-1 137.
[30] GUILLEVIC M, KOUBA M, MOUROT J. Effect of a linseed diet or a sunflower diet on performances, fatty acid composition, lipogenic enzyme activities and stearoyl-CoA-desaturase activity in the pig [J]. Livestock Science, 2009, 124(1), 288-294.
[31] BOTSOGLOU E, GOVARIS A, AMBROSIADI I, et al. Lipid and protein oxidation of α-linolenic acid-enriched pork during refrigerated storage as influenced by diet supplementation with olive leaves (Olea europea L.) or α-tocopheryl acetate [J]. Meat Science, 2013, 92(4): 525-532.
[32] CANNATA S, RATTI S, METEAU K, et al. Evaluation of different types of dry-cured ham by Italian and French consumers [J]. Meat Science, 2010, 84(4):601-606.
[33] ROSSI R, PASTORELLI G, CANNATA S, et al. Effect of long term dietary supplementation with plant extract on carcass characteristics meat quality and oxidative stability in pork [J]. Meat Science, 2013, 95(3) :542-548.
[34] MAIRESSE G, BENET M,METEAU K, et al. Effect of plant antioxidant in n-3 polyunsaturated fatty acid-enriched diet on fatty acid composition and sensorial attributes of dry-cured ham [J]. International Journal of Food Science & Technoloqy, 2011, 46(12):2 656-2 662.
[35] 何 钦,宋代军.日粮n-3多不饱和脂肪酸对母猪繁殖性能的影响[J].中国饲料,2012,12(2):8-10.
[36] MOREIRA F, CHEUICHE Z M, RIZZOTO G, et al. Metabolic and reproductive parameters in prepubertal gilts after omega-3 supplementation in the diet[J]. Anim Reprod Science, 2016,170:178-183.
[37] SPENCER J D,WILSON L,WEBEL S K,et al.Effect of feeding protected n-3 polyunsaturated fatty acids (FertiliumTM)on litter size in gilts [J].J Anim Sci,2004,82(Suppl.1):211.
[38] LAURIDSEN C,DDNIELSEN V. Lactational dietary fat levels and sources influence milk composition and performance of sows and their progeny [J]. Livestock Production Science, 2005, 91(1-2):95-105.
[39] PAPADOPOULOS G A, MAES D G, VAN WEYENBERG S,et al. Peripartal feeding strategy with different n-6:n-3 ratios in sows:effects on sows’ performance,inflammatory and periparturient metabolic parameters[J].British Journal of Nutrition,2009,101(3):348-357.
[40] SMIT M N, SPENCER J D, PATTERSON J L, et al. Effects of dietary enrichment with a marine oil-based n-3 LCPUFA supplement in sows with predicted birth weight phenotypes on birth litter quality and growth performance to weaning[J]. Animal An International Journal of Animal Bioscience, 2014, 9(3):471-480.
[41] 肖成林,田春庄,黄飞若,等. 母猪日粮中的鱼油对乳中脂肪酸和仔猪生长性能的影响[J]. 动物营养学报,2008,20(1):8-15.
[42] BRAZLE A E, JOHNSON B J, TITGEMEYER E C, et al. Fatty acid composition of the pig conceptus and endometrium in response to maternal omega-3 fatty acid supplementation[J].Journal of Animal Science, 2006, 84:66-66.
[43] TAUGBOL O,FRAMSTAD T,SAAREM K. Supplements of Cod Liver Oil to Lactating Sows. Influence on Milk Fatty Acid Composition and Growth Performance of Piglets [J]. Transboundary and Emerging Diseases, 1993, 40(1-10):437-443.
[44] MALDJIAN A, PENNY P C , NOBLE R C, et al. Docosa-hexaenoic acid-rich marine oils and improved reproductive efficiency in pigs[M]∥De VRIESE S R, CHRISTOPHE A B. Male fertility and lipid metabolism.Champaign:AOCS Press,2003.
[45] CASTELLANO C A, AAUDE I, LAFOREST J P, et al. Fish oil diets alter the phospholipid balance, fatty acid composition, and steroid hormone concentrations in testes of adult pigs [J]. Theriogenology, 2011, 76( 6) : 1 134-1 145.
[46] LIU Q, ZHOU Y F, DUAN R J, et al. Effects of dietary n-6:n-3 fatty acid ratio and vitamin E on semen quality, fatty acid composition and antioxidant status in boars[J]. Animal Reproduction Science, 2015, 162:11-19.
[47] ESTIENNE M J, HARPER A F, CRAWFORD R J. Dietary supplementation with a source of omega-3 fatty acids increases sperm number and the duration of ejaculation in boars[J]. Theriogenology, 2008, 70(1):70-76.
[48] YESTE M, BARRERA X, COLL D, et al. The effects on boar sperm quality of dietary supplementation with omega-3 polyunsaturated fatty acids differ among porcine breeds[J]. Theriogenology, 2011, 76(1):184-196.
(责任编辑:舒莲梅)
Research progress of ω-3 polyunsaturated fatty acid in swine production
HU Jin-jie,CAO Xia,WU Zhi-feng
(Hunan Jiuding Technology (Group) Co., Ltd.,Changsha 410003,China )
ω-3 polyunsaturated fatty acid (ω-3 PUFAs) is a kind of essential fatty acids and an important source of energy in the body, with a variety of physiological functions. We introduced the common raw materials and its content in the swine production which was rich in ω-3 PUFAs, and reviewed the immune function of pigs and its application in improving pork quality, production quality and boar semen quality,which provided some reference in the further promotion and application of ω-3 PUFAs.
ω-3 polyunsaturated fatty acids;swine;essential fatty acids;physiological functions; source of energy
2016-10-17;
2016-11-08
胡金杰 (1987-),男,硕士,研究方向为动物营养与饲料。
10.7633/j.issn.1003-6202.2017.01.012
S816.4
A
1003-6202(2017)01-0048-06
猜你喜欢
纺织科学研究(2023年9期)2023-10-23 11:18:12
中国生殖健康(2020年5期)2021-01-18 03:00:04
中国生殖健康(2018年5期)2018-11-06 07:15:54
天然产物研究与开发(2018年9期)2018-10-08 03:26:00
中国麻业科学(2018年6期)2018-04-09 11:22:12
商周刊(2017年6期)2017-08-22 03:42:51
纺织科学研究(2017年4期)2017-05-17 04:00:02
纺织服装流行趋势展望(2016年6期)2016-05-04 03:52:43
食品界(2016年4期)2016-02-27 07:36:48
合成化学(2015年10期)2016-01-17 08:56:06
