
032基因缺失猪痘病毒载体的构建及鉴定
2017-01-17 00:46蔺辉星范红结
安徽科技学院学报 2016年6期
蔺辉星,周 红,范红结
(南京农业大学 动物医学院,江苏 南京 210095)
032基因缺失猪痘病毒载体的构建及鉴定
蔺辉星,周 红,范红结*
(南京农业大学 动物医学院,江苏 南京 210095)
目的:构建毒力减弱的猪痘病毒载体。方法:以猪痘病毒野生毒株NT1501株为亲本毒株,通过同源重组方法,构建缺失032毒力基因的猪痘病毒SPV△032。检测SPV△032在不同种属来源细胞系中的培养滴度,并通过动物试验来检测其毒力。结果:猪痘病毒SPV△032株与野生NT1501株在PK15和ST等猪源细胞中的培养滴度差异不显著,均可达到5.0×106TCID50左右。猪痘病毒SPV△032株在Vero、MDBK等其他物种来源的细胞系中不能进行复制,连续传代2~3代以后,细胞培养物中检测不到猪痘病毒基因,与野生NT1501株培养结果一致。动物试验结果表明,用高浓度SPV△032接种实验猪,接种部位皮肤未出现任何症状,比野生NT1501株的毒力显著降低。接种SPV△032实验猪的细胞免疫和体液免疫反应水平均显著高于野生NT1501株 (P<0.05)。接种SPV△032的实验猪不会通过粪便等途径向环境中排毒。结论:本研究构建了032基因缺失猪痘病毒SPV△032,该病毒的毒力与野生NT1501株相比显著下降,可以作为弱毒猪痘病毒载体,用于兽用疫苗研发。
猪痘病毒;032基因;载体
猪痘病毒(Swinepox virus, SPV)是脊椎动物痘病毒亚科(Chordopoxvirinae)猪痘病毒属(Suipoxvirus)的唯一成员[1]。猪是SPV唯一的天然宿主,其他动物均不被SPV感染[2]。猪感染SPV后可出现猪痘症状,该病于1842年首次在欧洲报道,目前在全世界范围内流行。SPV在表皮角质形成细胞中增殖,皮肤以外的其他组织很少受到影响,通常病变轻微[3]。由于SPV能够同时刺激免疫动物的体液免疫反应以及细胞免疫反应,具有基因组容量大以及生物安全性高等优点,故常作为载体被用于兽用疫苗研发。从临床分离到的SPV野生毒株对猪具有一定的致病力,因此不能直接作为疫苗载体,需要对其进行致弱,降低其毒力,以保证猪痘病毒载体疫苗的安全性。猪痘病毒基因组共编码150个基因,依次命名为001-150基因。通过与其他痘病毒(如痘苗病毒)毒力相关基因的同源性分析发现,009、032、135、145基因可能与SPV的毒力相关[4-6]。其中032基因全长534 bp,所编码的蛋白与痘苗病毒编码的双链RNA依赖性蛋白激酶抑制因子同源[7],能够抑制干扰素的生成,降低宿主的天然免疫应答,导致感染动物容易出现猪痘症状。本研究通过构建032基因缺失猪痘病毒SPV△032,对其毒力以及生物学特性进行评价,以期构建弱毒猪痘病毒载体,用于兽用疫苗研发。
1 材料与方法
1.1 病毒与细胞
猪痘病毒NT1501株,2015年分离自南通某猪场,保存于本实验室。猪肾细胞(PK15)、非洲绿猴肾细胞(Vero)以及牛肾细胞(MDBK)等分别购自美国典型培养物保存中心(ATCC)或中科院上海细胞库,用含5%~10%胎牛血清(FBS)的DMEM细胞培养液进行培养。
1.2 猪痘病毒032基因缺失载体构建
以猪痘病毒NT1501株基因组为模板,用引物LF1/LF2和RF1/RF2分别PCR扩增032基因的左、右同源重组交换臂LF和RF。以pEGFP-N1质粒为模板,用引物GFP1/GFP2通过PCR方法扩增出绿色荧光蛋白筛选标记基因gfp。以pUC19质粒(Takara)为骨架,通过酶切连接方法,依次插入左、右同源重组交换臂LF和RF以及绿色荧光蛋白筛选标记基因gfp,得到猪痘病毒032基因缺失载体pUCLGR。
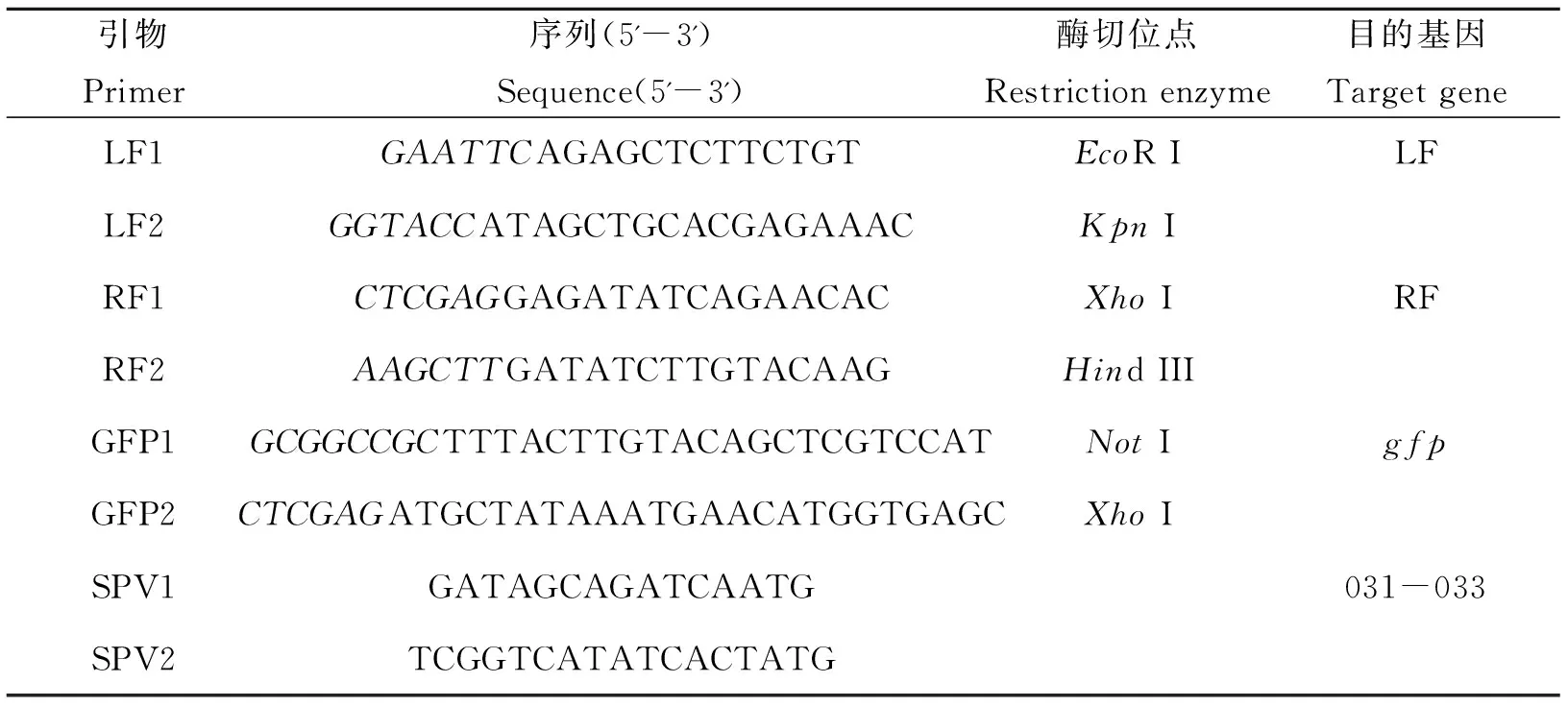
表1 本研究所用到的引物
斜体=限制性酶切位点。
1.3 032基因缺失猪痘病毒的构建
将PK15细胞培养于6孔细胞培养板中,待长成单层细胞后,加入0.01感染复数(MOI)的猪痘病毒NT1501株对细胞进行感染, 37 ℃孵育2 h后,用pUCLGR质粒进行转染,具体操作步骤按照转染试剂Lipofectamine 2000的说明书进行,转染后3 d将细胞培养物冻融两次后收集培养液。将收集的培养液接入6孔细胞板中的PK15单层细胞,进行新一轮的感染,37 ℃孵育2 h后,弃去孔中的细胞培养液,加入含1.5%甲基纤维素的DMEM细胞培养液,继续培养3~5 d。在荧光显微镜中观察到的绿色荧光噬斑即为032基因缺失猪痘病毒。在荧光显微镜下,用200 μL微量移液器挑取含绿色荧光的细胞,冻融2次后再次接种PK15单层细胞。重复10轮筛选纯化后,可得到纯化的032基因缺失猪痘病毒,命名为SPV△032。用引物SPV1/SPV2对该猪痘病毒进行PCR鉴定。
1.4 体外增殖能力检测以及遗传稳定性检测
对032基因缺失猪痘病毒SPV△032进行体外增殖能力进行检测。用0.01 MOI的病毒感染PK15、Vero和MDBK等细胞,37 ℃孵育2 h,中间摇动3次,弃去感染液,加入40 mL含2%胎牛血清的DMEM细胞培养液,继续培养至80%细胞发生病变(约72 h),将细胞反复冻融2次,收集细胞培养液,测定SPV△032的培养滴度。将收集的细胞培养液,再次接种同种细胞,进行连续传代,每隔10代取样,分别测定细胞培养液中SPV△032的培养滴度,并进行全基因组测序,以评价其遗传稳定性。
1.5 毒力评价
45头1月龄长白猪,随机分为3组,每组15头。前2组分别用SPV△032株以及NT1501株进行皮肤穿刺接种,第3组只注射生理盐水,作为阴性对照。每种病毒分别以1.0×106TCID50、1.0×105TCID50、1.0×104TCID50、1.0×103TCID50和1.0×102TCID50剂量进行接种,2 mL/头,每个剂量分别接种3头猪。每隔8 h观察猪皮肤表面症状,每隔48 h前腔静脉采血1 mL,提取血清中的病毒DNA,用引物SPV1/SPV2对猪痘病毒基因进行PCR检测。每隔3 d收集猪场内蚊虫、水源、饲料和粪便,提取样品中的病毒DNA,用引物SPV1/SPV2对猪痘病毒基因进行PCR检测,评估SPV△003的毒力以及扩散能力。
1.6 细胞因子检测
采集接种后第10 d、20 d的血清,用ELISA试剂盒(ExCell Bio)检测SPV△032株以及NT1501株接种猪血清中的IFN-γ和IL-4水平,具体操作步骤按照试剂盒说明书进行,根据相应的标准曲线进行计算。
1.7 统计学分析
所有的数据进行单因素方差分析,P<0.05被认为差异显著。
2 结果与分析
2.1 猪痘病毒032基因缺失载体构建及鉴定
用EcoR I/Hind III对猪痘病毒032基因缺失载体pUCLGR(图1A)进行双酶切鉴定,1 %琼脂糖凝胶电泳检测可见3824 bp和2158 bp 两个条带,酶切片段大小与预期结果一致(图1B)。测序结果表明,本试验所构建的猪痘病毒转移载体pUCLGR与目的基因序列完全一致。
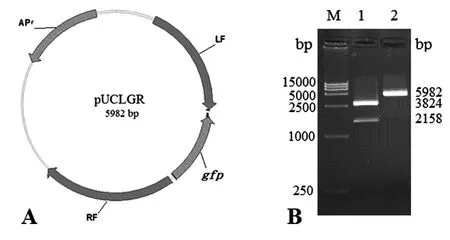
图1 猪痘病毒032基因缺失载体pUCLGR双酶切鉴定
2.2 032基因缺失猪痘病毒的纯化
重组猪痘病毒在含有甲基纤维素的培养基中培养约3~4 d,即可在荧光显微镜下观察到明显的绿色荧光噬斑(图2),挑取单个绿色荧光痘斑置于0.5 mL DMEM维持液中,反复冻融3次后用于下一轮纯化。纯化一直进行到出现在细胞板内的所有病毒噬斑均为绿色荧光噬斑,得到的032基因缺失猪痘病毒称作SPV△032株。结果表明,SPV△032株中检测不到032基因,032基因缺失猪痘病毒构建成功。
图2 032基因缺失猪痘病毒SPV△032的纯化
2.3 体外增殖能力检测以及遗传稳定性检测
在PK15和ST两种猪源细胞和十种非猪源细胞系中对032基因缺失猪痘病毒SPV△032进行遗传稳定性的评估。结果表明(表2),在PK15和ST猪源细胞系中,SPV△032株前4代的培养滴度有所下降,随传代次数增多,培养滴度逐渐提高,第6代以后接近野生毒株NT1501株的培养滴度(5.0×106TCID50)。而在非猪源细胞系中不能进行复制,传代2~3代以后就检测不到猪痘病毒基因。连续传代60代以上,SPV△032株的培养滴度始终维持在5.0×106TCID50左右。测序结果表明,各代次的SPV△032株未发生基因突变,遗传稳定性良好。
表2 基因缺失猪痘病毒SPV△032在不同细胞中的培养滴度(TCID50/log10)
a未检测到猪痘病毒基因。
2.4 毒力评价结果
动物试验结果表明,1.0×104TCID50以上剂量的SPV野生毒株NT1501株接种组试验猪注射部位皮肤出现直径约2~3 mm的痘疹,数量不多,第5 d明显可以观察到,16 d以内全部结痂脱落,除此之外未出现其他症状。SPV△032各浓度试验组均未出现皮肤症状,所有猪体温、饮食以及精神状态与阴性对照组猪相比,无显著差异。对接种猪血清中的病毒DNA进行检测,结果均为阴性,表明SPV△032只会感染皮肤组织,不会进入到血液中。对猪场内蚊虫、水源、饲料和粪便样品中的病毒DNA进行检测,结果均为阴性,表明SPV△032不会随粪便等释放到环境中,具有较高的生物安全性。
2.5 细胞因子检测结果
机体的免疫反应主要是由Th1和Th2 类T细胞亚群诱导。Th1细胞主要产生IFN-γ,IL-2和TNF-β等,主要诱导细胞免疫反应以及炎症反应;而Th2细胞主要产生IL-4、IL-6和IL-10等,主要诱导体液免疫反应和嗜酸性粒细胞积聚。SPV△032所诱导的机体免疫反应的类型可以通过测定血清中IFN-γ和IL-4的水平来进行间接评估。分别用ELISA检测试剂盒(ExCell Bio)对采集的试验猪血清中的IFN-γ和IL-4进行测定,结果表明(图3),SPV△032接种猪血清中IFN-γ和IL-4含量显著高于其他两组(P<0.05),表明SPV△032能够同时增强细胞免疫反应和体液免疫反应。
图3 细胞因子IFN-γ和IL-4水平检测
3 结论与讨论
SPV是脊椎动物痘病毒亚科猪痘病毒属的唯一成员,在自然条件下仅感染猪,通常病变轻微[8-9]。当前对SPV的研究主要集中在将其作为兽用疫苗载体方面[10-13]。Van der Leek等构建了表达伪狂犬病毒糖蛋白gp50-gp63的重组猪痘病毒,经皮肤穿刺或肌肉注射途径接种实验猪,21 d后分别有90%、100%的实验猪产生了伪狂犬病毒血清中和抗体[14]。本课题组构建了表达猪圆环病毒2型(PCV2)Cap蛋白的重组猪痘病毒,该重组猪痘病毒可诱导免疫仔猪产生高水平的PCV2中和抗体,可有效防制PCV2强毒株的攻击[15]。上述研究结果表明,重组猪痘病毒载体可高效表达外源蛋白,能诱导免疫动物产生高效的细胞免疫应答和体液免疫应答,是一种具有巨大应用前景的疫苗载体。
SPV载体疫苗的安全性问题一直是人们关注的焦点。SPV 032基因是其重要的毒力基因之一,能够降低宿主的天然免疫应答,导致感染动物容易出现猪痘症状。本研究构建了缺失032基因的猪痘病毒SPV△032、SPV△032株与野生NT1501株在PK15和ST等猪源细胞中的培养滴度差异不显著,均可达到5.0×106TCID50左右。猪痘病毒SPV△032株在Vero、MDBK等其他物种来源的细胞系中不能进行复制,连续传代2~3代以后,细胞培养物中检测不到猪痘病毒基因,与野生NT1501株培养结果一致。动物试验结果表明,1.0×104TCID50以上剂量的SPV野生毒株NT1501株会引起接种猪的皮肤出现局部痘斑,但不会导致其他症状。而1.0×106TCID50以上剂量的SPV△032株接种猪的皮肤未出现任何病理症状,SPV△032株比野生NT1501株的毒力显著降低。接种SPV△032实验猪的细胞免疫和体液免疫反应水平均显著高于野生NT1501株。接种SPV△032的实验猪不会通过粪便等途径向环境中排毒。上述研究结果表明,032基因缺失猪痘病毒SPV△032与野生NT1501株相比毒力显著下降,可以作为弱毒猪痘病毒载体,用于兽用疫苗研发。
[1]AITKEN W A. Swine pox[J]. The North American Veterinarian, 1948, 29(6): 353.
[2]Kasza L, Griesemer R A. Experimental swine pox.[J]. American Journal of Veterinary Research, 1962, 23(4):443-451.
[3]CHEVILLE N F. The cytopathology of swine pox in the skin of swine[J]. American Journal of Pathology, 1966, 49(2): 339-352.
[4]AFONSO C L, TULMAN E R, LU Z, et al. The genome of swinepox virus.[J]. Journal of Virology, 2002, 76(2):783-790.
[5]ISHIDO S, CHOI J K, LEE B S, et al. Inhibition of natural killer cell-mediated cytotoxicity by kaposi's sarcoma-associated herpesvirus K5 protein[J]. Immunity, 2000, 13(3): 365-374.
[6]STEVENSON P G, EFSTATHIOU S, DOHERTY P C, et al. Inhibition of MHC class I-restricted antigen presentation by gamma 2-herpesviruses[J]. Proceedings of the National Academy of Sciences of the United States of America, 2000, 97(15): 8455-8460.
[7]LANGLAND J O, JACOBS B L. The role of the PKR-inhibitory genes,E3L and K3L,in determining vaccinia virus host range[J]. Virology, 2002, 299(1): 133-141.
[8]RIYESH T, BARUA S, KUMAR N, et al. Isolation and genetic characterization of swinepox virus from pigs in India[J]. Comparative Immunology Microbiology and Infectious Diseases, 2016, 46: 60-65.
[9]MEDAGLIA M L, PEREIRA A D, FREITAS T R, et al. Swinepox virus outbreak, Brazil, 2011[J]. Emerging Infectious Diseases, 2011, 17(10): 1976-1978.
[10]FANG Yi-zhen, LIN Hui-xing, MA Zhe, et al. Construction and immunogenicity of recombinant swinepox virus expressing outer membrane protein L of salmonella[J]. Journal of Microbiology and Biotechnology, 2016, 26(7): 1173-1181.
[11]FAN H J, LIN H X. Recombinant Swinepox Virus for Veterinary Vaccine Development[J]. Methods Mol Biol, 2016,1349:163-175.
[12]YUAN Xiao-min, LIN Hui-xing, FAN Hong-jie. Efficacy and immunogenicity of recombinant swinepox virus expressing the A epitope of the TGEV S protein[J]. Vaccine, 2015, 33(32): 3900-3906.
[13]XU Jia-rong, YANG De-ji, HUANG Dong-yan, et al. Protection of Guinea pigs by vaccination with a recombinant swinepox virus co-expressing HA1 genes of swine H1N1 and H3N2 influenza viruses[J]. Archives of Virology, 2013, 158(3): 629-637.
[14]VAN DER LEEK M L, FELLER J A, SORENSEN G, et al. Evaluation of swinepox virus as a vaccine vector in pigs using an Aujeszky's disease(pseudorabies)virus gene insert coding for glycoproteins gp50 and gp63[J]. The Veterinary Record, 1994, 134(1): 13-18.
[15]LIN Hui-xing, MA Zhe, FAN Hong-jie, et al. Construction and immunogenicity of recombinant swinepox virus expressing capsid protein of PCV2[J]. Vaccine, 2012, 30(44): 6307-6313.
(责任编辑:马世堂)
Construction and Identification of the 032 Gene Deletion in Swinepox Virus
LIN Hui-xing, ZHOU Hong, FAN Hong-jie*
(College of Veterinary Medicine, Nanjing Agricultural University, Nanjing 210095,China)
Objective: To construct an attenuated swinepox virus (SPV) vector. Methods: The 032 gene deletion SPV△032 was constructed by homologous recombination with SPV NT1501 wild strain. The titers of the SPV△032 culture in different species-derived cell lines were measured, and the virulence of the SPV△032 was detected by animal experiment. Results: There was no significant difference in the culture titers between the SPV△032 strain and the wild NT1501 strain in PK15 and ST cell lines, and the culture titer of the SPV△032 was about 5.0×106TCID50. The SPV△032 could not be replicated in Vero, MDBK and other species-derived cell lines. After 2-3 passages, the swinepox virus gene could not be detected in the cell culture of the SPV△032, which was consistent with the culture of wild NT1501 strain . The results of animal experiments showed that the inoculation site of the SPV△032 inoculated pigs with high concentration did not show any symptoms, and the vi-rulence of the SPV△032 was significantly lower than that of the wild NT1501 strain. The cellular immune response and humoral immune response of the SPV△032 inoculated pigs were significantly higher than those of wild NT1501 strain inoculated pigs (P<0.05). The SPV△032 virus will not give emission to the environment through the feces of the SPV△032 inoculated pigs. Conclusion: The 032 gene deletion SPV△032 was constructed in this study. The virulence of the SPV△032 was significantly lower than that of the wild NT1501 strain, and could be used as attenuated vector in veterinary vaccine development.
Swinepox virus; 032 gene; Vector
2016-09-15
江苏省农业科技自主创新资金项目(CX(15)1056,CX(16)1028)。
蔺辉星(1985-),男,河北省石家庄市人,博士,讲师,主要从事兽医微生物学及免疫学、兽医生物技术研究。*通讯作者:范红结,教授,E-mail: fhj@njau.edu.cn。
S828.9
A
1673-8772(2016)06-0001-06
猜你喜欢
甘肃畜牧兽医(2022年11期)2023-01-05
昆明医科大学学报(2022年2期)2022-03-29
现代畜牧科技(2021年9期)2021-10-13
华侨大学学报(自然科学版)(2021年4期)2021-07-30
阅读(科学探秘)(2020年4期)2020-07-04
农药科学与管理(2019年9期)2019-11-23
农药科学与管理(2019年6期)2019-11-23
爱你(2019年21期)2019-06-21
农民致富之友(2017年12期)2018-01-31
现代检验医学杂志(2015年2期)2015-02-06
