
乙型肝炎病毒X蛋白的优势氨基酸序列与热点突变位点的生物信息学分析
2017-01-05 11:00:25王葳何永刚浦永兰潘少坤谢幼华
微生物与感染 2016年6期
王葳,何永刚,浦永兰,潘少坤,3,谢幼华
1. 复旦大学基础医学院教育部/卫生部医学分子病毒学重点实验室,上海 200032; 2. 太仓市第一人民医院感染科,太仓 215400; 3. 上海交通大学医学院附属第九人民医院口腔预防科,上海 200011
·论著·
乙型肝炎病毒X蛋白的优势氨基酸序列与热点突变位点的生物信息学分析
王葳1,何永刚1,浦永兰2,潘少坤1,3,谢幼华1
1. 复旦大学基础医学院教育部/卫生部医学分子病毒学重点实验室,上海 200032; 2. 太仓市第一人民医院感染科,太仓 215400; 3. 上海交通大学医学院附属第九人民医院口腔预防科,上海 200011
乙型肝炎病毒X蛋白(hepatitis B virus X protein,HBx)全长154个氨基酸,与肝癌发生密切相关。为确定HBx的优势氨基酸序列和热点突变位点,在GenBank中下载所有HBx的氨基酸序列13 950条,剔除插入突变、缺失突变和起始密码子非甲硫氨酸的序列,最后保留7 126条。通过分析这7 126条序列,计算出HBx每个位点的氨基酸分布情况,出现频率最高的氨基酸为该位点的优势氨基酸,其他氨基酸为突变氨基酸。154个位点的优势氨基酸组成HBx优势氨基酸序列。突变率>10.0%的热点突变位点有32个。其中第36、42、44、87、88和127位氨基酸有4种(突变率>1.0%)以上突变形式,具有较高的多态性。与肝癌密切相关的K130M/V131I双突变率为34.7%。通过7 126条HBx序列与优势序列的同源性比较,随机选出其中50条序列(2条与优势序列同源性<75%,48条同源性为76%~99%),与23条参考序列及优势序列共同构建系统发生树。结果显示,HBx优势氨基酸序列属于基因型C,这与基因型C为全球主要流行型一致。本研究首次系统性分析了GenBank中HBx的优势序列,确定了32个HBx热点突变位点和6个多态性较高的位点,为基于HBx突变的基础和应用研究奠定了基础。
乙型肝炎病毒X蛋白;热点突变;优势序列
乙型肝炎病毒(hepatitis B virus,HBV)感染引起的慢性肝炎、肝硬化(liver cirrhosis,LC)和肝癌(hepatocellular carcinoma,HCC)是全球性的公共卫生威胁,给患者尤其是发展中国家的患者带来沉重负担[1]。HBV X蛋白(HBV X protein,HBx)全长154个氨基酸,对HBV的感染和复制极为重要,是HBV诱发HCC的重要原因之一,且其相关突变与HCC病情进展密切相关[2]。HBx的热点突变位点主要集中在第26~45位氨基酸,其他突变位点较为散发[3]。HBx基因与核心启动子(core promoter,Cp)重叠。在Cp区常见的A1762T/G1764A双突变可导致HBx突变(K130M/V131I),与HCC密切相关[4-5]。因此,研究HBx的热点突变及频率有可能为发现新的HCC相关突变位点提供依据和启示。本研究利用生物信息学方法系统分析了GenBank中HBx的优势氨基酸序列、点突变发生率和基因型,为基于HBx突变的基础和应用研究奠定了基础。
1 材料与方法
1.1 材料
使用MEGA6、GenBank、Prism 6等软件。参考序列及基因型(序列号-基因型)如下:X02763.1-A、CAA36231.1-A、AAD40216.1-A、D00329.1-B、AB073846.1-B、BAK32996.1-B、CAA28288.1-C、AAM95244.1-C、BAA32912.1-C、CAA46360.1-D、AAA45504.1-D、CAA59513.1-D、CAA53341.1-E、BAA89320.1-E、CAA49453.1-F、BAB17959.1-F、AAG49731.1-F、AAF34736.1-G、BAB82385.1-G、AAL99435.1-G、AAM09038.1-H、AAM09050.1-H、AAM09062.1-H。
1.2 方法
1.2.1 HBx氨基酸序列的下载和筛选 在GenBank中下载所有HBx的氨基酸序列13 950条。剔除插入突变(>154个氨基酸)和缺失突变(<154个氨基酸)的序列,保留7 148条;再剔除起始氨基酸为非甲硫氨酸(methionine,M)的序列,保留7 128条。通过MEGA中的Align比对,剔除2条同时存在插入突变和缺失突变的长度为154个氨基酸的序列,最终保留7 126条HBx全长序列用于分析。
1.2.2 HBx优势氨基酸序列和热点突变位点的确定 通过计算得出7 126条HBx全长序列上每个位点的20种氨基酸的出现频率,指定出现频率最高的氨基酸为优势氨基酸,其余氨基酸为突变氨基酸,其出现频繁为该氨基酸的突变率。将每个位点优势氨基酸组成的虚拟序列定义为优势序列。
1.2.3 系统发生树的构建 通过对7 126条HBx全长序列与优势序列的同源性比较,得出每条序列与优势序列的同源性关系。随机选出与优势序列同源性递增的50条序列(2条同源性<75%和48条同源性为76%~99%的序列中,每升高1%选择两个序列),与23条参考序列(A~H共8种基因型)和优势序列构建系统发生树。使用近邻结合法(neighbor-joining,NJ),并用Bootstraping测试5 000次,其中重复性>70%者显示在节点上。
2 结果
2.1 HBx氨基酸序列的下载和筛选
为获得用于分析的HBx氨基酸序列,在GenBank中搜索关键词HBV X,设置条件为Protein,得到13 950条序列;剔除>154个氨基酸的序列(n=616)和<154个氨基酸的序列(n=6 186),得到7 148条序列;再剔除起始氨基酸为非甲硫氨酸的序列(n=20)及同时存在插入和缺失突变的长度为154个氨基酸的序列(n=2),得到7 126条序列(图1),用于后续分析。

图1 HBx氨基酸序列筛选流程图
Fig.1 Flowchart for screening of HBx protein sequences downloaded from GenBank
2.2 HBx优势氨基酸序列的构建和热点突变位点
分析7 126条HBx每一位点上20种氨基酸的出现频率,指定出现频率最高的氨基酸为优势氨基酸,将154个优势氨基酸组成的虚拟序列定义为HBx优势序列(图2)。相对优势序列,除起始氨基酸甲硫氨酸外,将7 126条HBx中第2~154位的每个位点除优势氨基酸以外的其他氨基酸均定义为突变氨基酸,分别计算每个位点的突变率。结果显示,就全序列而言,突变率为0.2%~60.6%。其中82个位点的氨基酸突变率<1.0%;24个位点的氨基酸突变率为1.0%~4.9%;15个位点的氨基酸突变率为5.0%~9.9%;11个位点的氨基酸突变率为10.0%~19.9%;8个位点的氨基酸突变率为20.0%~29.9%;5个位点的氨基酸突变率为30.0%~39.9%;5个位点的氨基酸突变率为40.0%~49.9%;1个位点的氨基酸突变率为50.0%~59.9%(第36位为54.2%);2个位点的氨基酸突变率为60.0%~69.9%(第30位为60.5%、第88位为60.6%)。
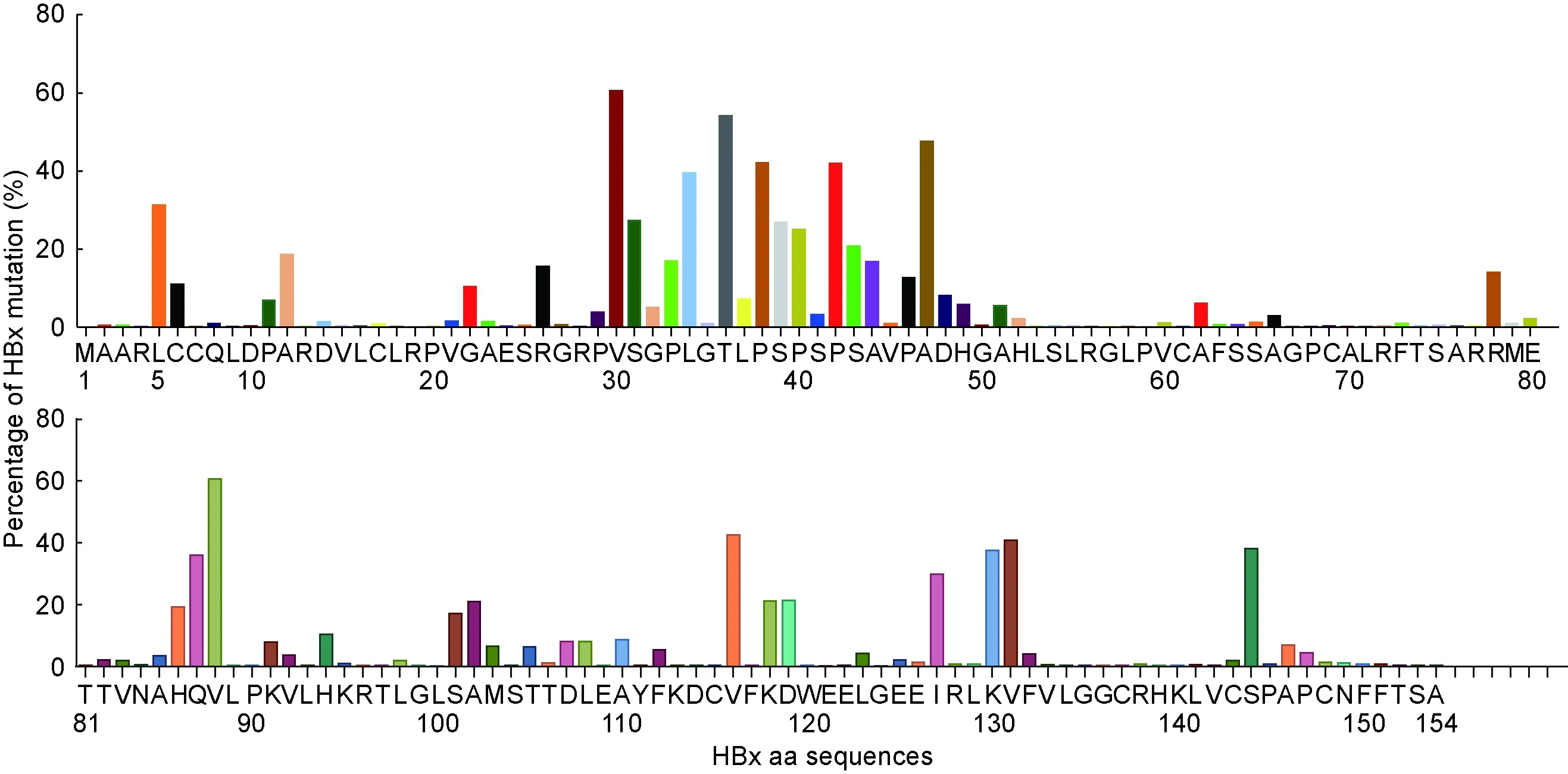
Each residue position and deduced dominant amino acid of HBx are displayed in the horizontal coordinate, and the mutation rate in each residue position is displayed in the vertical coordinate.
图2 HBx优势氨基酸序列和各位点的突变率
Fig.2 Deduced dominant amino acid sequence of HBx and mutation rate in each residue position
HBx氨基酸序列中有32个位点的总突变率>10.0%(表1)。这32个突变位点中,主要突变(单个氨基酸突变率>1.0%)形式只有1个的位点有12个:C6Y、G22S、P33S、L34F、P38S、S39P、S43P、P46S、H94Y、V116L、D119E和K130M;主要突变形式有2个的位点有9个:L5V/M、A12S/T、R26C/S、S31P/A、P40A/S、A47T/S、R78C/Y、V131I/L和S144A/V;主要突变形式有3个的位点有5个:V30L/F/I、H86R/P/S、A102V/G/T、S101P/A/F和K118T/N/A;主要突变形式有4个的位点只有1个:P42S/A/L/T;主要突变形式有5个的位点有4个:A44V/I/G/L/T、V88N/I/F/S/A、T36A/P/D/S/G和I127T/V/S/L/M;主要突变形式有6个的位点只有1个:Q87R/G/H/W/L/M(表2)。
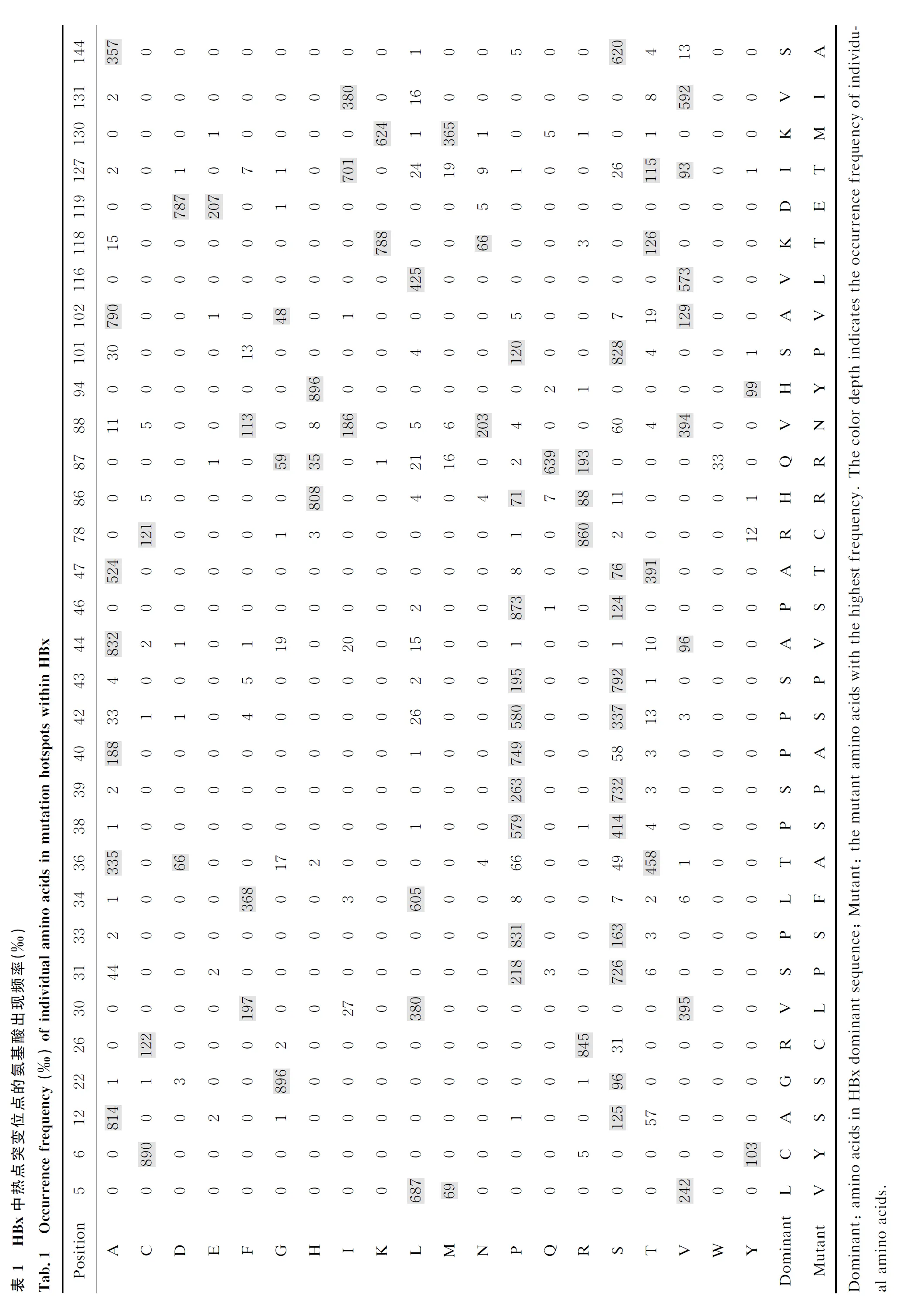

上述HBx热点突变位点中,K130M/V131I双突变率为34.7%,K130M单突变率为1.5%,V131I单突变率为3.1%。32个热点突变位点中,第36、42、44、87、88和127的突变种类在4种以上(突变率>1.0%),这6个热点突变位点表现出较高的单氨基酸多态性。
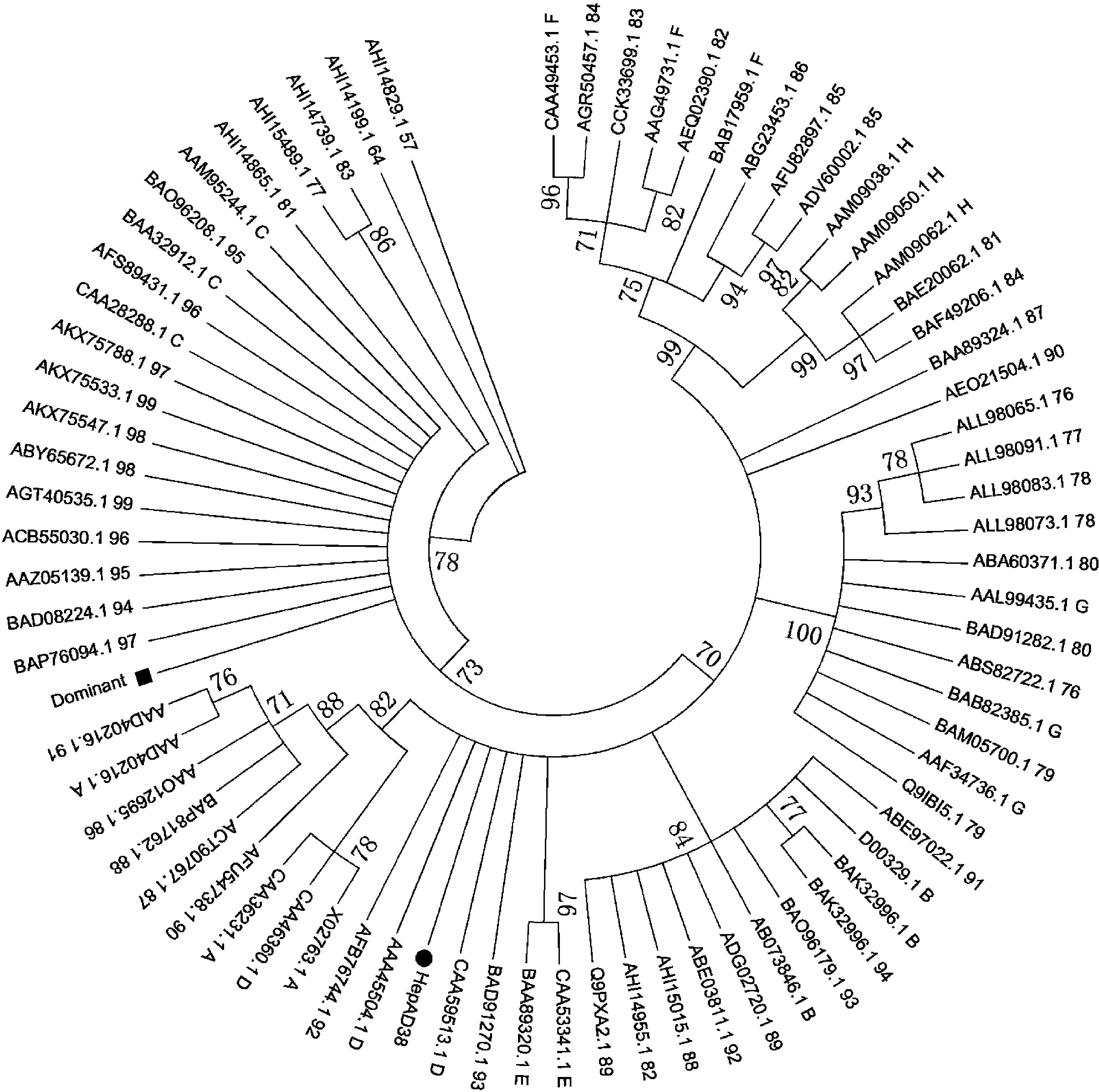
The relationships of the different amino acid sequences of HBx are indicated by phylogenetic tree constructed using the neighbor-joining (NJ) method, implemented in the MEGA6 package. The phylogenetic tree test is bootstrapped (5 000 replicates). The bootstrap values are shown below or above the branches. Each reference sequence (n=27) is represented by a specific letter, which represents it’s genotype; Each selected sequence (n=50) is represented by a specific number, which represents its homology to HBx dominant sequence. The dominant sequence using “Dominant” instead is marked with black box; HBx sequence in HepaAD38 is marked with black circle.
图3 HBx优势序列和50条按同源性递增随机筛选的序列系统发生树
Fig.3 Phylogenetic tree for the dominant HBx and 50 selected HBx sequences based on an increasing homology
2.3 HBx的系统发生关系
7 126条HBx氨基酸序列与优势氨基酸序列之间的同源性为54.5%~98.7%。其中同源性<75%有2条(同源性分别为54.5%和57.1%);75%≤同源性<80%有95条(占1.3%);80%≤同源性<85%有332条(占4.7%);85%≤同源性<90%有733条(占10.3%);90%≤同源性<95%有4 433条(占62.2%);同源性≥95%有1 531条(占21.5%)。
通过NJ系统发生树(图3)可看出,HBx优势氨基酸序列、基因型C的参考序列与同源性为94%~99%的11条序列归为一支,属基因型C;同源性为82%~86%的6条序列与基因型F的参考序列归为一支,属基因型F;同源性为81%和84%的2条序列与基因型H的参考序列归为一支,属基因型H;同源性为76%~80%的9条序列与基因型G的参考序列归为一支,属基因型G;同源性为82%~94%的8条序列与基因型B的参考序列归为一支,属基因型B;基因型为86%~93%的7条序列和HepAD38与基因型A和D的参考序列归为一支,分型不明确;同源性为87%和90%的2条序列介于基因型G与H之间,分型不明确;同源性为57%、64%、77%、81%和83%的5条序列离8种基因型参考序列分支均较远,分型不明确。
3 讨论
HBx相对分子质量约为16 500,含154个氨基酸[6]。HBx是一个多功能蛋白,在HBV生活史及HCC的发生和发展中起重要作用。HBx具有广泛的生物学功能,可通过与多个细胞内转录因子如活化蛋白1(activator protein 1,AP-1)等作用调控基因表达,也可与多条信号通路中的关键因子作用而影响细胞周期、细胞凋亡和自吞噬等多个生理过程[7]。最新研究发现,HBx能结合Smc5/6并将其带至溶酶体降解,从而保护病毒基因转录免受宿主细胞的抑制[8]。
HCC是病死率最高的癌症之一[9-10]。HBx基因突变与HCC的发生和进程密切相关[2]。目前已报道的与HCC相关的突变位点有V5M(G1386A)、P38S(C1485T)[11]、G50R(G1521C/A)[12]、D80E(G1613A)[13]、H94Y(C1653T)[11,13]、S101P(T1674C)[14]、I127T/N/S(T1753V)[13-14]、K130M(A1762T)和V131I(G1764A)[11,13]。与HCC相关的双突变位点有K130M/V131I(A1762T/G1764A)和L30F/S144A[12],3个位点共突变有V5M/K130M/V131I[11]。未发现与HCC有明显相关性的热点突变位点有A23T(G1440A)[14]、G31R、G32R(G1467C)[14]、A/P36T(G/C1479A)[14]、S43P和Q87R[12]。
深入研究较多的HCC相关突变位点是K130M/V131I (A1762T/G1764A)。A1762T/G1764A位于Cp区,首先在HBV e抗原(HBV e antigen,HBeAg)阴性患者中发现[15]。该双突变能抑制HBeAg表达,增强HBV复制[5]。自然发生的K130M/V131I双突变频率在HCC患者与HBV慢性无症状携带者(chronic asymptomatic carrier,ASC)、慢性乙型肝炎(chronic hepatitis B,CHB)患者和LC患者中存在显著差异,且随着病情进展在ASC及CHB、LC、HCC患者中显著增加,此双突变可能无须进展至LC就能直接诱发HCC[5]。
除与基础核心启动子区(basal core promoter, BCP)和EnⅡ重叠外,HBx基因的5′端与聚合酶的RNase H功能域部分重叠,3′端与PreC部分重叠,但HBx基因与这两个基因的编码框均不同框。HBx与RNase H重叠部分长250 bp(1374~1623位核苷酸,对应HBx的1~84位氨基酸);与PreC重叠部分长15 bp(1814~1828位核苷酸,对应HBx的147~154位氨基酸)。RNase H在HBV复制过程中主要起负链DNA合成时降解pgRNA的作用。目前对RNase H突变的临床研究较少。细胞水平研究发现,基因型D的RNase H中D689、R703、E718、D737、D777和R781突变会抑制HBV DNA合成[16]。其中Asp777和Arg781落在与HBx的重叠部分,分别对应27~28位和31~32位氨基酸。27~28位氨基酸在HBx上的突变率很低,而31~32位氨基酸在HBx上的突变率相对较高。HBx第26~52位氨基酸的突变率较高,RNase H在此区域的突变率也较高。RNase H与HBx的编码框并不相同,在一定程度上可限制RNase H和HBx突变,但两者可能在此区域的保守性均较低。PreC/C基因编码的HBe与HBV建立慢性感染密切相关。影响HBeAg表达的主要原因为BCP突变(A1762T/G1764A)和基因型依赖的G1896A突变[17]。在PreC中与HBx的重叠部位为1~8位氨基酸,处于信号肽的N端,在HBeAg分泌过程中被切割[18]。HBx第147位氨基酸为脯氨酸(P,CCA),其对应的最后一个碱基为A,单独A突变为147位同义突变,对HBx无影响。HBx第148~154位氨基酸突变率较低,可能因为其与PreC重叠但不同框。
目前研究主要集中在少数热点突变位点,尚缺乏对HBx所有位点进行系统的突变研究。本研究从GenBank中筛选到7 126条HBx序列,首次系统分析了HBx的优势氨基酸序列,并对HBx每个氨基酸位点的突变率进行了分析。结果显示,热点突变位点基本与他人研究一致,集中在26~52位氨基酸的反式激活因子区域(图2),主要热点突变位点有R26C/S、V30L/F/I、S31P/A、P33S、T36A/P/D/S/G、P38S、S39P、P42S/A/L/T、S43P、A44V/I/G/L/T、P46S和A47T/S;其他热点突变位点相对较为分散,主要有L5V/M、C6Y、A12S/T、G22S、R78C/Y、H86R/P/S、Q87R/G/H/W/L/M、V88N/I/F/S/A、H94Y、S101P/A/F、A102V/G/T、V116L、K118T/N/A、D119E、I127T/V/S/L/M、K130M、V131I/L和S144A/V。其中与HCC密切相关的K130M/V131I双突变发生频率为34.7%,与其他研究较为吻合[5,11,14]。
HBx第5位优势氨基酸是L,其出现频率为68.7%,主要突变位点有L5V(24.2%)和L5M(6.9%);但有报道为V5M突变,且V5M与HCC密切相关[11]。第30位优势氨基酸为V,其出现频率为39.5%,主要突变位点有V30L(38.0%)、V30F(19.7%)和V30I(2.7%);有研究报道为V/L30F突变[14],认为V和L均为野生型。第36位优势氨基酸为T,其出现频率为45.8%,主要突变位点有T36A(33.5%)、T36D(6.6%)、T36P(6.6%)、T36S(4.9%)和T36G(17.0%);有研究报道为A/P36T突变[14],认为A和P为野生型。上述3个位点中第5位和第36位优势氨基酸的出现频率比突变频率显著高,表明以前认为的野生型可能要做调整。第30位优势氨基酸的出现频率(39.5%)与主要突变频率(38.0%)接近,可认为优势氨基酸有两种。
HBx第36、42、44、87、88和127位氨基酸的多态性频率较高,突变率>1.0%的有T36A/P/D/S/G、P42S/A/L/T、A44V/I/G/L/T、Q87R/G/H/W/L/M、V88N/I/F/S/A和I127T/V/S/L/M,尚无报道。
HBx优势序列及与其同源性较高(94%~99%)的序列归于基因型C,表明提交到数据库中的序列为基因型C的序列数量最多,这与国内HBV主要流行型为C型和B型[19]及全球主要流行型为C型[20]一致;与HBx优势氨基酸序列同源性较低(76%~86%)的序列归于流行性较低的F、G和H型。有研究报道,HBV基因型B比基因型C发生K130M/V131I双突变的概率低,且HCC患者中HBV基因型C的K130M/V131I双突变比例比基因型B高[5]。在母婴垂直传播研究中发现,HBV基因型C比基因型B突变更快,更易发生K130M/V131I双突变[21]。由此可见,HBV基因型C的K130M/V131I双突变与HCC相关性较高,也提示对HBV基因型C的ASC及CHB、LC患者检测K130M/V131I(A1762T/G1764A)双突变具有重要意义。
综上所述,本研究系统性研究了HBx优势氨基酸序列和每个位点氨基酸的突变情况,确定了32个HBx热点突变位点,首次报道了6个氨基酸多态性位点,为HBx突变研究及基于HBx突变的临床诊断与治疗奠定了基础。
[1] Lavanchy D. Hepatitis B virus epidemiology, disease burden, treatment, and current and emerging prevention and control measures [J]. J Viral Hepat, 2004, 11(2): 97-107.
[2] Geng M, Xin X, Bi LQ, Zhou LT, Liu XH. Molecular mechanism of hepatitis B virus X protein function in hepatocarcinogenesis [J]. World J Gastroenterol, 2015, 21(38): 10732-10738.
[3] Hwang GY, Huang CJ, Lin CY, Wu CC. Dominant mutations of hepatitis B virus variants in hepatoma accumulate in B-cell and T-cell epitopes of the HBx antigen [J]. Virus Res, 2003, 92(2): 157-164.
[4] Baptista M, Kramvis A, Kew MC. High prevalence of 1762(T) 1764(A) mutations in the basic core promoter of hepatitis B virus isolated from black Africans with hepatocellular carcinoma compared with asymptomatic carriers [J]. Hepatology, 1999, 29(3): 946-953.
[5] Yang Z, Zhuang L, Lu Y, Xu Q, Tang B, Chen X. Naturally occurring basal core promoter A1762T/G1764A dual mutations increase the risk of HBV-related hepatocellular carcinoma: a meta-analysis [J/OL]. Oncotarget, 2016. http://www.impactjournals.com/oncotarget/index.php?journal=oncotarget&page=article&op=view&path%5B%5D=7123&path%5B%5D=20283.
[6] Tiollais P, Pourcel C, Dejean A. The hepatitis B virus [J]. Nature, 1985, 317(6037): 489-495.
[7] Bouchard MJ, Schneider RJ. The enigmatic X gene of hepatitis B virus [J]. J Virol, 2004, 78(23): 12725-12734.
[8] Decorsière A, Mueller H, van Breugel PC, Abdul F, Gerossier L, Beran RK, Livingston CM, Niu C, Fletcher SP, Hantz O, Strubin M. Hepatitis B virus X protein identifies the Smc5/6 complex as a host restriction factor [J]. Nature, 2016, 531(7594): 386-389.
[9] Zhuang LP, Xu LT, Wang P, Jiang Y, Yong P, Zhang CY, Zhang HB, Meng ZQ, Yang PY. Na+/K+-ATPase alpha 1 subunit, a novel therapeutic target for hepatocellular carcinoma [J]. Oncotarget, 2015, 6(29): 28183-28193.
[10] Yang ZG, Zhang JL, Lu YF, Xu QA, Wang Q, Zhang WS, Chen SS, Lu LQ, Chen XR. Aspartate aminotransferase-lymphocyte ratio index and systemic immune-inflammation index predict overall survival in HBV-related hepatocellular carcinoma patients after transcatheter arterial chemoembolization [J]. Oncotarget, 2015, 6(40): 43090-43098.
[11] Lee JH, Han KH, Lee JM, Park JH, Kim HS. Impact of hepatitis B virus (HBV) X gene mutations on hepatocellular carcinoma development in chronic HBV infection [J]. Clin Vaccine Immunol, 2011, 18(6): 914-921.
[12] Zhang X, Ding HG. Key role of hepatitis B virus mutation in chronic hepatitis B development to hepatocellular carcinoma [J]. World J Hepatol, 2015, 7(9): 1282-1286.
[13] Park YM. Clinical utility of complex mutations in the core promoter and proximal precore regions of the hepatitis B virus genome [J]. World J Hepatol, 2015, 7(1): 113-120.
[14] 王淼,胡艳芳,史为涛,王勤英. 原发性肝细胞癌患者乙型肝炎病毒X基因突变检测及其临床意义 [J]. 中华肝脏病杂志, 2015,23(8):599-603.
[15] Okamoto H, Tsuda F, Akahane Y, Sugai Y, Yoshiba M, Moriyama K, Tanaka T, Miyakawa Y, Mayumi M. Hepatitis B virus with mutations in the core promoter for an e antigen-negative phenotype in carriers with antibody to e antigen [J]. J Virol, 1994, 68(12): 8102-8110.
[16] Ko C, Shin YC, Park WJ, Kim SA, Ryu WS. Residues Arg703, Asp777, and Arg781 of the RNase H domain of hepatitis B virus polymerase are critical for viral DNA synthesis [J]. J Virol, 2014, 88(1): 154-163.
[17] Tong S, Revill P. Overview of hepatitis B viral replication and genetic variability [J]. J Hepatol, 2016, 64(1 Suppl): S4-S16.
[18] Ito K, Kim KH, Lok AS, Tong S. Characterization of genotype-specific carboxyl-terminal cleavage sites of hepatitis B virus e antigen precursor and identification of furin as the candidate enzyme [J]. J Virol, 2009, 83(8): 3507-3517.
[19] Li HM, Wang JQ, Wang R, Zhao Q, Li L, Zhang JP, Shen T. Hepatitis B virus genotypes and genome characteristics in China [J]. World J Gastroenterol, 2015, 21(21): 6684-6697.
[20] Jazayeri SM, Alavian SM, Carman WF. Hepatitis B virus: origin and evolution [J]. J Viral Hepat, 2010, 17(4): 229-235.
[21] Wu Q, Xu C, Li J, Li L, Yan G, Yue L, Zeng Y, Huang H, Deng G, Wang Y. Evolution and mutations of hepatitis B virus quasispecies in genotype B and C during vertical transmission [J]. J Med Virol, 2016, 88(6): 1018-1026.
s. PAN Shaokun, E-mail: span10@fudan.edu.cn;XIE Youhua, E-mail: yhxie@fudan.edu.cn
Bioinformatic determination of dominant amino acid sequence and mutation hotspots in hepatitis B virus X protein
WANG Wei1, HE Yonggang1, PU Yonglan2, PAN Shaokun1,3, XIE Youhua1
1.KeyLaboratoryofMedicalMolecularVirologyofMinistriesofEducationandHealth,SchoolofBasicMedicalSciences,FudanUniversity,Shanghai200032,China; 2.DepartmentofInfectiousDiseases,TaicangFirstPeople’sHospital,Taicang215400,China; 3.DepartmentofPreventiveDentistry,ShanghaiNinthPeople’sHospital,ShanghaiJiaoTongUniversitySchoolofMedicine,ShanghaiKeyLaboratoryofStomatology,Shanghai200011,China
Hepatitis B virus X protein (HBx), a 154-amino acid protein encoded by hepatitis B virus (HBV), has been implicated in the development of hepatocellular carcinoma (HCC). The aim of the present study is to establish a complete and dominant peptide sequence of HBx. A total of 13 950 HBx protein sequences were retrieved from GenBank. After excluding the non-complete ones that harbor insertions, deletions and non-methionine start amino acid, 7 126 were included in the study. Occurrence frequencies of the 20 amino acids at each position of HBx were calculated. The amino acid with the highest frequency at each position was designated as the dominant amino acid at the position and all the dominant residues constituted the complete and dominant peptide sequence of HBx. All the non-dominant amino acids were taken as the mutants. 32 hotspots with a mutation rate over 10.0% were detected. The positions 36, 42, 44, 87, 88, and 127 showed a higher polymorphism, having more than 4 kinds of mutations with a rate over 1.0%. The K130M/V131I dual mutation, regarded as a high risk factor of HBV-related HCC, showed an occurrence frequency at 34.7% in this study. The homology between the dominant sequence and each of the 7 126 HBx sequences was calculated. 50 sequences, which included 2 sequences with a homology to the dominant below 75% and 48 sequences with a homology between 76%-99%, were chosen for phylogenetic tree construction. The phylogenetic tree with the dominant HBx, 50 selected HBx and 23 reference sequences showed that the dominant HBx belonged to genotype C. This study has systematically constituted a calculated HBx peptide sequence with dominant amino acid at each position, laying the foundation for the basic and applied research on HBx.
Hepatitis B virus X protein; Mutation hotspot; Dominant sequence
国家重点基础研究发展计划(2012CB519002)
潘少坤,谢幼华
2016-04-05)
猜你喜欢
加油站服务指南(2022年6期)2022-07-28 06:07:02
遗传(2021年10期)2021-11-01 10:30:08
中国医学创新(2020年11期)2020-06-08 10:38:37
中华肺部疾病杂志(电子版)(2020年2期)2020-05-07 00:34:24
中国实验诊断学(2020年4期)2020-04-29 14:30:28
车迷(2019年10期)2019-06-24 05:43:28
快乐语文(2018年7期)2018-05-25 02:32:00
现代检验医学杂志(2015年6期)2015-02-06 01:44:02
中国记者(2014年6期)2014-03-01 01:39:53
实验动物与比较医学(2014年5期)2014-02-28 14:53:10
