
栗斑腹鹀对寄生和捕食风险的行为反应
2016-12-29 03:50陈艳东姜云垒张立世刘丽秋
东北师大学报(自然科学版) 2016年4期
陈艳东,姜云垒,2,秦 博,张立世,刘丽秋
(1.吉林农业大学动物科学与技术学院,吉林 长春 130118;2.吉林农业大学动物生产及产品质量安全教育部重点实验室,吉林 长春 130118;3.吉林省莫莫格国家级自然保护区管理局,吉林 白城 137316)
栗斑腹鹀对寄生和捕食风险的行为反应
陈艳东1,姜云垒1,2,秦 博3,张立世1,刘丽秋1
(1.吉林农业大学动物科学与技术学院,吉林 长春 130118;2.吉林农业大学动物生产及产品质量安全教育部重点实验室,吉林 长春 130118;3.吉林省莫莫格国家级自然保护区管理局,吉林 白城 137316)
捕食和巢寄生是影响栗斑腹鹀存活和繁殖成功的重要因素,面对不同风险,其行为对策可能不同.在内蒙古赤峰市阿鲁科尔沁旗敖伦花和内蒙古通辽市扎鲁特旗嘎亥图,通过摆放标本实验,探讨了栗斑腹鹀在捕食和寄生风险下的行为反应差异.结果表明:栗斑腹鹀对捕食者红隼(Falcotinnunculus)反应最强烈;其次为寄生者大杜鹃(Cuculuscanorus);对中性对照山斑鸠(Streptopeliaorientalis)反应强度最弱.说明栗斑腹鹀具有识别不同类型风险的能力,并能采取不同的行为对策应对不同风险.
栗斑腹鹀;标本;行为对策;报警声
捕食可定义为一种生物摄取其他种生物个体的全部或部分为食,前者称为捕食者,后者称为猎物或被捕食者[1].对动物反捕食对策的研究始于19世纪末[2],研究发现捕食者与猎物长期协同进化,猎物形成了保护色、假死、逃跑、集体防御、报警等行为对策以逃避被捕食[1-5].而采取反捕食对策的前提就是能够识别捕食者[6],大多数动物都能够识别环境中的捕食者,并做出一定的反捕食行为来降低捕食风险[7-9],因此,猎物需要评估捕食风险的大小从而选择合适的行为对策[10-11].
鸟类种间巢寄生降低了宿主的繁殖成功率和适合度[12].宿主主要的反寄生策略为识别寄生卵和雏鸟并丢弃或弃巢[13-15].实际上,最好的反寄生策略是寻找安全的繁殖地[16-17]或阻止巢寄生者于宿主巢中产卵[18-20],当寄生者出现在巢附近时,亲鸟增加警戒并进行防御,能够降低巢寄生发生的概率[21-22].
栗斑腹鹀(Emberizajankowskii)隶属于雀形目(Passeriformes)、鹀科(Emberizidae)、鹀属(Emberiza),单型种,无亚种分化,是世界上稀有的鸟类之一.由于栗斑腹鹀较小的分布范围和种群数量急剧下降,2010年被世界自然保护联盟(IUCN)列为濒危物种[23].现已对栗斑腹鹀分布和繁殖习性[24]、种群数量变化[25]、巢址选择[26]、双亲递食率[27]、现状和保护对策[28]等开展了研究,研究发现捕食和寄生是影响栗斑腹鹀繁殖成功率的主要因素[29].本文以栗斑腹鹀为研究对象,在孵化期采用摆放标本实验,检验栗斑腹鹀是否能够识别捕食和寄生风险,分析其是否采取不同的行为对策应对不同的风险,以为栗斑腹鹀的保护研究提供基础资料.
1 方法
1.1 研究区域
研究样地位于内蒙古自治区赤峰市阿鲁科尔沁旗(43°21′43″~45°24′20″N,119°02′15″~121°01′E)和内蒙古通辽市扎鲁特旗(43°50′13″~45°35′31″N,119°34′48″~121°56′50″E),海拔179.2~1 444.2 m.研究区东邻科尔沁右翼中旗,南与赤峰市翁牛特旗接壤,西与巴林左旗、巴林右旗交界,北与锡林郭勒盟西乌珠穆沁旗毗邻;以低山丘陵和倾斜冲积平原为主要地貌特征;属温带大陆性气候;降雨主要集中在6—8月份,年平均降雨量为300~400 mm.研究区域内草本植被主要有贝加尔针茅(StipaBaicalensis)、羊草(Leymuschinensis)、北京隐子草(Cleistogeneshancei)、万年蒿(Artenmisiagmelinii)、猪毛蒿(Artemisiascoparia)等,木本植被主要有西伯利亚山杏(Prunussibirica)和蒙古黄榆(Ulmusmacrocarpa)两种[27].
1.2 实验方法
2014—2015年5—7月,在研究区域内采用巢位统计法确定栗斑腹鹀繁殖巢的位置,监测首枚卵日期、孵化期等.对孵化期内栗斑腹鹀亲鸟(n=20对)进行摆放标本实验:选择红隼标本代表捕食风险,大杜鹃标本代表寄生风险,山斑鸠标本作为中性对照,代表无风险[30].选择晴朗无风鸟类活动高峰期(6:00~9:00,16:00 ~19:00 )进行实验.标本摆放在距巢1 m远,距地1 m高的山杏树上.实验开始前将标本用布遮盖,一人移除遮盖物,并立刻退到10 m以外的位置,另一人在15 m处记录亲鸟行为,实验持续10 min.从至少一只亲鸟飞回距巢25 m处开始计时[5].为了确定亲鸟的位置,以巢为中心,在巢四周5,10,15,20,25 m处做标记,从标本摆放到第一只亲鸟飞回时间间隔不超过30 min.标本摆放顺序是随机的,针对同一巢的两次实验至少间隔1 d.观察者和受试亲鸟之间的接触尽量最小化,采用焦点动物取样法和所有事件取样法结合记录亲鸟行为并选取以下行为变量:
进巢:标本摆放后,栗斑腹鹀是否回巢继续孵化[30];回巢时间:从第一只亲鸟到达至其进巢的时间[30];报警:亲鸟返回后是否报警[6];报警级别:0(无报警声),1(报警时间<2 min),2(报警时间<6 min),3(报警时间>6 min)[31];快速靠近标本:亲鸟返回后是否快速飞过或靠近标本[6];快速靠近标本次数;返回时间:从标本摆放到至少一只亲鸟返回巢附近25 m的时间[31];返回数量:从标本摆放到返回巢附近25 m处的亲鸟数量[31].统计繁殖巢某行为是否出现时,用“1”代表“是”,用“0”代表“否”.
1.3 数据处理
对三种风险处理下栗斑腹鹀是否进巢、报警和快速靠近标本的质量性状数据用非参数Cochran’s检验,组间差异用McNemar检验;非参数Friedman’s检验检验快速靠近标本的次数、从标本摆放到返回巢附近25 m处的栗斑腹鹀数量、报警级别,组间差异分析用Wilcoxon signed-ranks检验[32].从标本摆放到至少一只亲鸟返回巢附近25 m处的返回时间用单因素方差分析(one-way ANOVA),多重比较用最小显著差数法(LSD);独立样本t检验检验回巢时间.显著性水平设置为0.05,所有检验都是双尾检验,数据用平均值±标准差表示.以上分析均在SPSS22.0统计分析软件上完成.
2 结果
捕食风险、寄生风险和无风险下栗斑腹鹀的8种行为指标的显著性差异和多重比较分析结果见表1.由表1可见,栗斑腹鹀的进巢行为与风险类型相关(Cochran’s检验:Q=14.923,P=0.001),捕食风险下无进巢继续孵化行为,寄生风险和无风险下分别有8巢和11巢亲鸟进巢继续孵化,捕食风险和无风险的进巢行为差异极显著(McNemar检验:PH-S=0.001),捕食风险和寄生风险的进巢行为差异极显著(McNemar检验:PH-D=0.008),寄生风险和无风险的进巢行为无显著差异.t检验结果显示,回巢时间也无显著差异(t=0.929,df=17,P=0.366),放置大杜鹃时回巢时间为(290.8±144.3)s(n=8),放置山斑鸠时回巢时间为(227.9±146.4)s(n=11).
报警行为的发生与风险类型无关(Cochran’s检验:Q=4.308,P=0.116),但是报警级别与风险类型有关(Friedman检验:χ2=10.131,P=0.006).捕食风险的报警级别最高,为1.6±0.9(n=20),寄生风险的报警级别为0.85±0.7(n=20),无风险的报警级别为0.8±0.9(n=20);捕食风险和寄生风险的报警级别差异极显著(Wilcoxon检验:Z=-2.697,PH-D=0.007),捕食风险和无风险的报警级别差异极显著(Wilcoxon检验:Z=-2.949,PH-S=0.003).
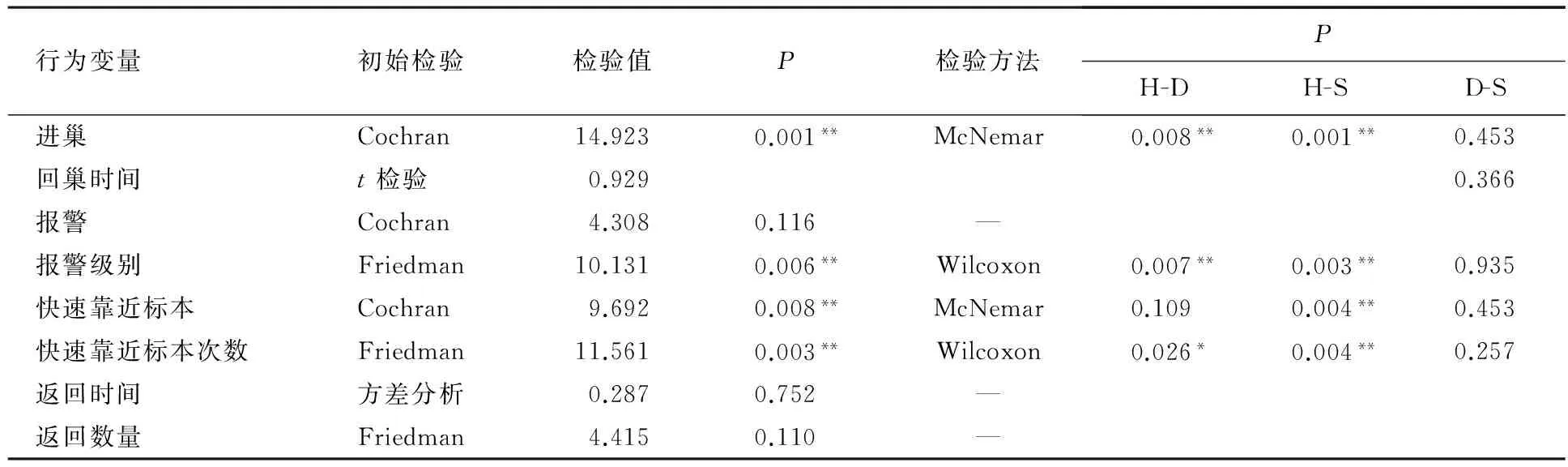
表1 栗斑腹鹀对三种风险行为反应
注:H-D表示红隼和大杜鹃的组间差异,H-S表示红隼和山斑鸠的组间差异,D-S表示大杜鹃和山斑鸠的组间差异.**差异极显著,*差异显著.
快速靠近标本行为与风险类型相关(Cochran’s检验:Q=9.692,P=0.008),捕食风险、寄生风险和无风险下分别有11,5,2巢亲鸟快速靠近标本,快速靠近标本行为在捕食风险和无风险间差异极显著(McNemar检验:PH-S=0.004).快速靠近标本的次数与风险类型有关(Friedman检验:χ2=11.561,P=0.003),捕食风险下,栗斑腹鹀靠近标本(0.9±1.1)次(n=20),无风险下快速靠近标本(0.1±0.3)次(n=20),捕食风险和无风险下快速靠近标本次数差异极显著(Wilcoxon检验:Z=-2.877,PH-S=0.004);寄生风险下,快速靠近标本(0.3±0.4)次(n=20),捕食风险和寄生风险下快速靠近标本的次数差异显著(Wilcoxon检验:Z=-2.228,PH-D=0.026),寄生风险和无风险下快速靠近标本次数差异不显著(Wilcoxon检验:Z=-1.134,PD-S=0.257).
捕食风险下,除繁殖巢亲鸟外,同种其他栗斑腹鹀个体也会在捕食者附近做出行为反应,存在共同御敌的现象,每巢有(2.1±1.2)只(n=20)个体返回;寄生风险和无风险下仅繁殖亲鸟返回,分别有(1.6±0.5)只(n=20)和(1.5±0.5)只(n=20)返回,栗斑腹鹀返回数量与风险类型无关(Friedman检验:χ2=4.415,P=0.110).捕食风险下,(328.5±277.8)s(n=20)后,第一只亲鸟飞回巢附近;寄生风险下,(336.6±325.8)s(n=20)后返回;无风险下,(398.5±356.8)s(n=20)后返回.三种风险下,栗斑腹鹀的返回时间差异不显著(F=0.287,df=2,P=0.752).
3 讨论
3.1 栗斑腹鹀对风险的识别
栗斑腹鹀对捕食风险的反应最强烈,出现共同御敌的现象,返回的栗斑腹鹀数量最多;发生快速靠近标本行为的巢数最多;快速靠近标本的次数也最多.寄生风险和无风险下行为反应依次减弱,说明栗斑腹鹀能够识别三种风险并且有合适的行为对策,结果与中国家燕(Hirundorustica)对雀鹰和大杜鹃的识别研究一致.中国家燕不是大杜鹃的常见宿主,大杜鹃虽在形态特征上高度模仿雀鹰,但中国家燕仍可区分雀鹰(Accipiternisus)和大杜鹃[30].实验从侧面支持了一般巢的防御假说[18],栗斑腹鹀并未将杜鹃标本当作特殊的敌人,而表现出与山斑鸠标本相似的反应,这与黑顶林莺(Sylviaatricapilla)对大杜鹃和原鸽(Columbalivia)的识别的研究结果一致[31];黑顶林莺对大杜鹃和原鸽标本表现出相同级别的反应,也与田雀鹀(Spizellapusilla)对棕头牛鹂(Molothrusater)和冠蓝鸦(Cyanocittacristata)的研究结果一致[33].1999—2001年的栗斑腹鹀调查研究显示,161个损失卵中,5.6%为大杜鹃的巢寄生,11.8%被捕食,捕食和寄生是影响栗斑腹鹀繁殖成功的主要因素[29].巢寄生者一般在产卵初期对宿主构成威胁,将宿主的卵推出巢中[18],而我们的实验是在孵化中期开展的,寄生风险已经不是影响繁殖的主要因素了.
3.2 栗斑腹鹀风险对策
鸟类对风险的评估是动态的[34],需要权衡繁殖能量和存活能量的分配[35-36].研究发现,栗斑腹鹀在高捕食风险下,将更多的时间用于观察环境和防御捕食者,将能量分配给自身存活,减少进巢的继续孵化行为;低风险下,亲鸟继续进巢孵化,将能量分配给当前繁殖.因此可以认为,栗斑腹鹀在高捕食风险下会减少繁殖能量投资,增加存活能量投资.与赵亮(2005)[37]的研究结果相似,在捕食风险下,繁殖期角百灵(Eremophilaalpestris)和小云雀(Alaudagulgula)亲鸟通过缩短坐巢时间、延长消失时间、减少递食率等行为,降低当前繁殖投入,提高雏鸟的存活率.
3.3 栗斑腹鹀报警行为
鸟类的报警声也是一种对捕食者的行为应答,传递已经发现捕食者的信息,或利用鸣声技巧给捕食者带来错误的信息[38].报警声的内容与捕食者密切相关,其他个体可通过听觉感受器获得报警声的内容.鸟类有时会对任何入侵者发出报警声,也会发出高频错误的报警声[39].本研究发现,栗斑腹鹀在非捕食风险下也会发出报警声,对未构成任何威胁的山斑鸠标本仍有45%(9/20)的个体发出报警声,而对威胁不大的大杜鹃有75%(15/20)的个体报警.栗斑腹鹀这种防患于未然的错误报警行为是有益的,可以预防真正的捕食攻击,更有利于应对潜在的威胁.
[1] 牛翠娟,娄安如,孙儒泳,等.基础生态学[M].高等教育出版社,2002:141-142.
[2] CARO T.Antipredator defenses in birds and mammals [M].Chicago:University of Chicago Press,2005:7-8.
[3] 孟凡明,梁醒财.昆虫假死行为研究历史及现状[J].昆虫知识,2009(6):985-991.
[4] 尚玉昌.动物行为学[M].北京:北京大学出版社,2005:174-180.
[5] COURTER J R,RITCHISON G.Alarm calls of tufted titmice convey information about predator size and threat[J].Behavioral Ecology,2010,21(5):936-942.
[6] SUZUKI T N.Referential mobbing calls elicit different predator-searching behaviours in Japanese great tits [J].Animal Behaviour,2012,84(1):53-57.
[7] LIMA S L.Stress and decision-making under the risk of predation:recent developments from behavioral,reproductive,and ecological perspectives [J].Advances in the Study of Behaviour,1998,27(8):215-290.
[8] KATS L B,DILL L M.The scent of death:chemosensory assessment of predation risk by prey animals [J].Ecoscience,1998,5(3):361-394.
[9] STANKOWICH T,BLUMSTEIN D T.Fear in animals:a meta-analysis and review of risk assessment [J].Proceedings of the Royal Society of London B:Biological Sciences,2005,272(1581):2627-2634.
[10] CURIO E,KLUMP G,REGELMANN K.An anti-predator response in the great tit(Parusmajor):is it tuned to predator risk [J].Oecologia,1983,60(1):83-88.
[11] KLEINDORFER S,FESSL B,HOI H.Avian nest defence behaviour:assessment in relation to predator distance and type,and nest height [J].Animal Behaviour,2005,69(2):307-313.
[12] ROTHSTEIN S I.A model system for coevolution:avian brood parasitism [J].Annual Review of Ecology and Systematics,1990,21(1):481-508.
[13] LANGMORE N E,HUNT S,KILNER R M.Escalation of a coevolutionary arms race through host rejection of brood parasitic young [J].Nature,2003,422(6928):157-160.
[14] DAVIES N B,BROOKE M L.Cuckoos versus reed warblers:adaptations and counter adaptations [J].Animal Behaviour,1988,36(1):262-284.
[15] HOSOI S A,ROTHSTEIN S I.Nest desertion and cowbird parasitism:evidence for evolved responses and evolutionary lag [J].Animal Behaviour,2000,59(4):823-840.
[16] CLARKE A L,ØIEN I J,HONZA M,et al.Factors affecting reed warbler risk of brood parasitism by the common cuckoo [J].The Auk,2001,118(2):534-538.
[18] SEALY S G,NEUDORF D L,HOBSON K A,et al.Nest defense by potential hosts of the Brown-headed Cowbird:methodological approaches,benefits of defense,and coevolution [J].Oxford Ornithology Series,1998(9):194-211.
[19] GRIM T,HONZA M.Differences in behaviour of closely related thrushes(TurdusphilomelosandT.merula) to experimental parasitism by the common cuckoo (Cuculuscanorus) [J].BIOLOGIA-BRATISLAVA,2001,56(5):549-556.
[20] R∅SKAFT E,MOKSNES A,STOKKE B G,et al.Aggression to dummy cuckoos by potential European cuckoo hosts [J].Behaviour,2002,139(5):613-628.
[21] KLEINDORFER S,EVANS C,COLOMBELLI-NÉGREL D,et al.Host response to cuckoo song is predicted by the future risk of brood parasitism [J].Frontiers in Zoology,2013,10(1):1-10.
[22] GRIM T,KLEVEN O,MIKULICA O.Nestling discrimination without recognition:a possible defence mechanism for hosts towards cuckoo parasitism [J].Proceedings of the Royal Society of London B:Biological Sciences,2003,270:S73-S75.
[23] BIRD LIFE INTERNATIONAL.Species factsheet:Emberizajankowskii[DB/OL].http://www.bird life.Org,2013-12-03.
[24] 佟富春,高玮,肖以华,等.吉林白城地区草原栗斑腹鹀窝卵数、营巢成功率和繁殖成功率的研究[J].应用生态报,2002,13(3):281-284.
[25] 程瑾瑞,高玮,王海涛,等.栗斑腹鹀种群数量变化的分析[J].东北师大学报(自然科学版),2002,34(1):49-53.
[26] 高玮,王海涛,孙丹婷.栗斑腹鹀的栖息地和巢址选择[J].生态学报,2003,23(4):665-672.
[27] 陶慧娟,姜云垒,秦博,等.栗斑腹鹀双亲递食率比较研究[J].东北师大学报(自然科学版),2014,46(3):105-108.
[28] WANG H T,JIANG Y L,GAO W.Jankowski’s Bunting(Emberizajankowskii):current status and conservation [J].Chinese Bird,2011,1(1):251-258.
[29] JIANG Y L,GAO W,LEI F M,et al.Nesting biology and population dynamics of Jankowski’s Bunting(Emberizajankowskii) in Western Jilin,China[J].Bird Conservation International,2008,18(2):153-163.
[30] YU J P,WANG L W,XING X Y,et al.Barn swallows(Hirundorustica) differentiate between common cuckoo and sparrowhawk in China:alarm calls convey information on threat[J].Behavioral Ecology and Sociobiology,2016,70(1):171-178.
[31] GRIM T.Host recognition of brood parasites:implications for methodology in studies of enemy recognition[J].The Auk,2005,122(2):530-543.
[32] MUNDRY R,FISCHER J.Use of statistical programs for nonparametric tests of small samples often leads to incorrectPvalues:examples from animal behaviour [J].Animal Behaviour,1998,56(1):256-259.
[33] BURHANS D E.Enemy recognition by field sparrows[J].The Wilson Bulletin,2001,113(2):189-193.
[34] MONTGOMERIE R D,WEATHERHEAD P J.Risks and rewards of nest defence by parent birds [J].Quarterly Review of Biology,1988,63(2):167-187.
[35] COLOMBELLI-NÉGREL D,ROBERTSON J,KLEINDORFER S.Nestling presence affects the anti-predator response of adult superb fairy-wrens(Maluruscyaneus)[J].Acta Ethologica,2010,13(2):69-74.
[36] GHALAMBOR C K,MARTIN T E.Fecundity-survival trade-offs and parental risk-taking in birds[J].Science,2001,292(5516):494-497.
[37] 赵亮.繁殖期两种百灵科鸟类对捕食风险的行为响应[J].动物学研究,2005,26(2):113-117.
[38] PERRINS C.The purpose of the high-intensity alarm call in small passerines [J].Ibis,1968,110(2):200-201.
[39] HAFTORN S.Contexts and possible functions of alarm calling in the willow tit(Parusmontanus) the principle of ‘better safe than sorry’ [J].Behaviour,2000,137(4):437-449.
(责任编辑:方 林)
Behavioral response of Jankowski’s Bunting(Emberizajankowskii)when faced with predator and brood parasitism risk
CHEN Yan-dong1,JIANG Yun-lei1,2,QIN bo3,ZHANG Li-shi1,LIU Li-qiu1
(1.College of Animal Science and Technology,Jilin Agricultural University,Changchun 130118,China;2.The Key Laboratory of Animal Production and Product Quality and Security,Ministry of Education,Jilin Agricultural University,Changchun 130118,China;3.Jilin District Bureau of Momoge Nature Reserve,Baicheng 137316,China)
Predation and brood parasitism can induce reproductive success of Jankowski’s Bunting(Emberizajankowskii).When faced with different risk,their behavioral strategies may be different.To investigate different behavior to predator and brood parasites,dummy experiment was conducted at Ar Horqin Banner in Chifeng City and Jarud Banner in Tongliao City of Inner Mongolia.Used Kestrel dummy(referred to predator),common cuckoo(referred to brood parasites),Oriental turtle dove(neutral control),and found that response intensity of Jankowski’s Bunting declined from Kestrel through common cuckoo to Oriental turtle dove.The result show that Jankowski’s Bunting can identify the type of dummy or risk and adopt adaptive behavioral strategies.
Jankowski’s Bunting(Emberizajankowskii);dummy;behavioral strstegies;alarm call
1000-1832(2016)04-0106-05
10.16163/j.cnki.22-1123/n.2016.04.023
2016-02-09
国家自然科学基金资助项目(31172109);吉林省自然科学基金资助项目(20150101067JC).
陈艳东(1990—),女,硕士研究生;通讯作者:姜云垒(1962—),男,博士,教授,主要从事鸟类生态学研究.
Q 959.7+39 [学科代码] 180.5744
A
猜你喜欢
数学物理学报(2021年6期)2021-12-21
心声歌刊(2021年3期)2021-08-05
百科探秘·海底世界(2020年11期)2020-12-31
小聪仔(幼儿版)(2020年12期)2020-02-01
民族音乐(2018年5期)2018-11-17
乡村地理(2018年3期)2018-11-06
太原师范学院学报(自然科学版)(2018年2期)2018-08-17
铁道通信信号(2018年5期)2018-06-28
中外文摘(2016年13期)2016-08-29
汽车维护与修理(2016年10期)2016-07-10
