
大兴安岭北坡多年冻土区植物群落分类及其物种多样性对冻土融深变化的响应
2016-12-28 07:23郭金停韩风林布仁仓胡远满聂志文朱菊兰任百慧
生态学报 2016年21期
郭金停,韩风林,布仁仓,胡远满,马 俊,聂志文,朱菊兰,任百慧
1 中国科学院沈阳应用生态研究所,森林与土壤生态国家重点实验室,沈阳 110016 2 中国科学院大学,北京 100049 3 沈阳师范大学,沈阳 110034
大兴安岭北坡多年冻土区植物群落分类及其物种多样性对冻土融深变化的响应
郭金停1,2,韩风林1,2,布仁仓1,*,胡远满1,马 俊1,2,聂志文1,2,朱菊兰3,任百慧1,2
1 中国科学院沈阳应用生态研究所,森林与土壤生态国家重点实验室,沈阳 110016 2 中国科学院大学,北京 100049 3 沈阳师范大学,沈阳 110034
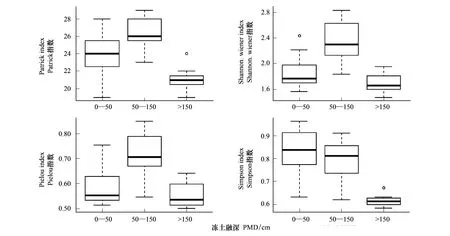
气候变化背景下冻土环境对地上植物群落的影响备受关注。鉴于此,选择了大兴安岭北坡作为研究区,应用双向指示种分析(TWINSPAN)和典范对应分析法(CCA)对大兴安岭北坡不同冻土融深的30个样地进行了群落分类,分析了物种多样性对冻土融深的响应。结果表明,1)研究区30个沟谷冻土样地植物群落可划分为3个群丛组,TWINSPAN的分类结果很好的反映了群丛组的分布与冻土融深的关系,即随着冻土融深由浅变深,群落由柴桦(Betulafruticosa)+狭叶杜香(Ledumpalustrevar.angustum)-苔草(Carexsubpediformis)群丛组逐渐过渡到柴桦(Betulafruticosa)-苔草(Carexsubpediformis)群丛组和柴桦(Betulafruticosa)+细叶沼柳(Salixrosmarinifolia)-苔草(Carexsubpediformis)群丛组,并在CCA二维排序图上得到了验证;2)地上植物群落的物种多样性指数随着冻土融深的增加表现出先上升后下降的单峰变化趋势;在50cm 大兴安岭北坡;冻土融深;物种多样性;TWINSPAN ;CCA 高寒生态系统是高寒环境下多年冻土的冻融交替作用发展起来的一种特殊生态系统[1],多年冻土的发育程度是维持其生态平衡的物质基础。大兴安岭位于欧亚大陆多年冻土带南缘,也是我国多年冻土的发育地之一,具有地温高、冻土厚度浅、热稳定性差、对气候变暖的反应十分敏感等特点[2- 3]。由于气候变暖,大兴安岭多年冻土正在由南向北逐步消退[4- 5],主要表现为最大季节融化深度增大、冻土厚度减薄、地温升高、融区扩大以及多年岛状冻土消失等[6]。这种变化将对区域生态系统产生巨大影响,主要表现为植物群落结构、物种多样性以及物种分布等诸多结构和生态功能发生不同程度的变化[7-9],并由此导致高寒生态系统消退加速[10- 12],继而对整个生态系统产生影响[13- 14]。因此认清植物群落对冻土融深变化的响应特征是研究大兴安岭冻土区植被恢复和物种多样性保护的首要工作。 迄今为止,对大兴安岭冻土的研究多局限于冻土分布、冻土的退化模拟、资源保护等[15- 18],而关于物种多样性对冻土融深变化响应的研究尚见报道。为此,本文应用双向指示种分析法(TWINSPAN)和典范对应分析法(CCA),对大兴安岭北坡冻土区的30个植被群落进行了排序和分类,并分析了群落变化的主要影响因子以及物种多样性对冻土融深的变化规律,旨在为冻土退化及其对全球变化的响应研究提供理论基础。 1.1 研究区的自然概况 研究区地处大兴安岭北坡(50°01′01″—53°26′25″N,119°07′02″—121°49′17″E),海拔620—960m。该地区属寒温带大陆性季风气候,冬季严寒而漫长,年均气温-4—-1℃,年均降水量350—550mm,无霜期80—100d。研究样地位于草地生态系统与森林生态系统的过渡带,物种组成较为丰富,多数物种属于寒温带物种,少数属于温带物种;植物群落的垂直结构比较简单,成层现象明显,分别由乔木层、灌木层、草本层和地被层四个层次组成。其中乔木层由兴安落叶松(Larixgmelinii)老头林和少量白桦(Betulaplatyphylla)组成;灌木层物种较多,其中柴桦(Betulafruticosa)、狭叶杜香(Ledumpalustrevar.angustum)、笃斯越桔(Vacciniumuliginosum)等为冻土植被群落的特征种;草本层物种最多,其中苔草(Carexsubpediformis)等为草本层的优势种;地被层主要由中位泥炭藓和白齿泥炭藓等各种泥炭藓所覆盖[19];土壤类型主要是草甸土和沼泽土,有机质含量较高,水分、养分丰富,土质较疏松,地表有积水[20]。 1.2 样方调查 2014年7月,采用系统取样方法,在研究区内选择基本相似生境条件下的30块样地进行调查(表1)。每个样地随机设置2m×2m灌木样方6个,1m×1m草本样方9—12个,以确保每个样地90%的物种被调查到。对于高度小于2m或胸径小于2.5cm的乔木,作为灌木调查。在每个样方中记录植物种的名称、多度、盖度、高度等数量指标,以及样地所在的经度、纬度、海拔高度、坡度、坡向、冻土融深等环境因子,并对每个样地活动层进行了土壤采样。 表1 调查样地基本情况 “—”代表在0—150cm范围内没有挖到冻土 1.3 研究方法 1.3.1 地理数据的获取 通过GPS获取每个样点的经度、纬度、海拔高度、坡度(表1)。 1.3.2 土壤数据的获取 将取土样放置烘箱,烘至恒重,土壤水分中自由态水以蒸汽形式全部散失掉,再称重量从而获得土壤含水量。采用直接挖深法获得每个样点的冻土融深(表1)。 1.3.3 群落分类 采用双向指示种分析法(TWINSPAN)和典范对应分析法(CCA)[21],对大兴安岭北坡30个沟谷冻土湿地的植物群落进行数量分类和排序分析,将冻土植被群落分成3个群丛组,两种方法分别用国际通用软件WinTWINS 2.3和CANOCO for Windows 4.5完成,并对群落变化的影响因子进行分析,所用软件为R 3.0.2vegan程序包。 1.3.4 不同群丛组物种组成分析 以重要值(IV)表示物种在3个群丛组的综合型数量指标,其计算公式如下: IV=(相对高度+相对盖度+相对频度)/3 1.3.5 物种多样性对冻土融深的响应分析 数据分析时将每个群丛组的若干样方的调查结果进行平均,选择能反应群丛组状况的多样性指数,采用Patrick丰富度指数、Simpson多样性指数、Shannon-Wiener多样性指数、Pielou均匀度指数[22],指数计算公式如下: Patrick丰富度指数 R=S Simpson多样性指数 Shannon-Wiener多样性指数 Pielou均匀度指数: 2.1 大兴安岭北坡冻土植被群落的TWINSPAN分类和CCA排序 应用TWINSPAN分类和CCA排序对大兴安岭北坡冻土植被群落30个样地进行了群落分类,结果显示,根据冻土融深由浅到深,研究区群落可分为3个群丛组,群从组的定义以及群落的分类原则和系统,这3个组分别代表3个植物群丛组,即植物群从组Ⅰ、Ⅱ、Ⅲ(Ⅰ柴桦+狭叶杜香-苔草群丛组、Ⅱ柴桦-苔草群丛组、Ⅲ柴桦+细叶沼柳-苔草群丛组)(图1)。 由分类结果显示,冻土融深(PMD)和海拔高度(Alt)、坡度(Slo)有线性正相关关系,与土壤含水量(SWC)有线性负相关关系(图1)。4个排序轴的特征值分别为0.487、0.376、0322和-0.433,CCA排序结果图能很好地反应3个群丛组与环境的关系,在冻土融深由浅到深的连续变化上,群落组也由群丛组Ⅰ逐渐变化到群丛组Ⅱ和群丛组Ⅲ(图1)。 研究结果显示,土壤含水量随冻土融深的增加呈现出明显的降低趋势,土壤含水量(SWC)与冻土融深(PMD)之间存在明显的负相关关系(R2=0.74,P<0.01)(图1)。 研究结果显示,冻土融深(PMD)、地形和土壤含水量3个因子对群落变化的解释率为38%,其中地形因子、冻土融深(PMD)和土壤含水量三者的综合效应为群落变化的最大解释因子(图2),除冻土融深、地形和土壤含水量以外还有其它因素也影响群落的变化,还有待于进一步研究。 图1 研究区冻土植物群落的CCA二维排序图Fig.1 Two-dimensional CCA ordination diagram of plant communities in the study area 图2 不同环境因素对群落变化的解释率Fig.2 Different environmental factors on the interpretation of the community change rate 2.2 不同群丛组物种组成分析 植物群丛组Ⅰ为柴桦+狭叶杜香-苔草群丛组。该群丛组主要分布于冻土融深为0—50cm范围内,属于冻土融深较浅的群落类型。灌木层以柴桦和狭叶杜香为优势种,其盖度分别为30%—45%和15%—60%,并伴有笃斯越桔和兴安落叶松等伴生种;草本层以白毛羊胡子草和大叶樟等作为优势种,并伴有三叶鹿药、北悬钩子、草问荆等伴生种(表2)。 表2 研究区各群丛组中优势种和主要伴生种重要值 *P<0.05;#:优势种或共优种;同行数据后不同字母表示差异显著(P<0.05) 植物群丛组Ⅱ为柴桦-苔草群丛组。该群丛组主要分布于冻土融深为50—150cm范围内,属于冻土融深较深的群落类型。灌木层以柴桦、笃斯越桔和柳叶绣线菊为优势种,且狭叶杜香为主要伴生种,除此以外还有细叶沼柳和小叶杜鹃等;草本丛除群丛组Ⅰ的伴生种外还有野古草、五脉山黧豆和北方拉拉藤等伴生种。 植物群丛组Ⅲ为柴桦+细叶沼柳-苔草群丛组。该群丛组主要分布于冻土融深大于150cm范围内,属于岛状冻土的群落类型。灌木层以柴桦、细叶沼柳为优势种,且笃斯越桔和金老梅等为主要伴生种,除此以外还有越桔柳和五蕊柳等;草本丛以大穗苔草大叶樟、苔草等为优势种并伴有五脉山黧豆、草问荆、小白花地榆、北方拉拉藤等伴生种。 2.3 物种多样性对冻土融深的响应 Patrick指数与冻土融深具有显著的相关性(R2=0.58,P<0.01)。PMD≤50cm时,物种种类较为丰富,物种数为23.83±2.44;50cm 图3 不同活动层埋深下物种多样性的变化Fig.3 Variations of plant species diversities with different depths of active layer of permafrost 3.1 大兴安岭北坡多年冻土区植物群落的TWINSPAN分类和CCA排序 本文运用了TWINSPAN分类和CCA排序将群落分为3个群丛组并得到了相互验证。由图1、图2和图3可以看出:在景观尺度范围内,海拔高度及坡度均是冻土发育的地形条件,地势较低的平缓地带是冻土发育的客观条件,冻土融深的大小也会直接影响到土壤含水量,融深较浅的土壤含水量较高[23- 25],冻土融深由浅到深群丛组也由Ⅰ柴桦+狭叶杜香-苔草群丛组逐渐过渡到Ⅱ柴桦-苔草群丛组和Ⅲ柴桦+细叶沼柳-苔草群丛组,该分类结果较客观的反映了植物群落类型的分布与环境因子的关系。3个群丛组间的明显过渡表明群落类型对生境的变化规律有显著地响应,由以上的4个因素差异造成了冻土区不同群丛组的群落组成存在一定的间断性。 3.2 气候变化情境下冻土群落变化 群丛组Ⅰ、群丛组Ⅱ和群丛组Ⅲ分别代表3个不同的群丛组,不同的群丛组所对应的冻土融深范围也不相同,随着冻土融深由浅变深,部分物种的重要值明显减小,如狭叶杜香、白毛羊胡子草、大叶樟等;另外一些物种的重要值明显增加,如柴桦、细叶沼柳等;随着冻土融深的增加,一些在冻土融深较浅的环境下不存在的物种也相继出现,如野古草、五脉山黧豆、北方拉拉藤、地榆、东北羊角芹、马蔺、二歧银莲花、和裂叶蒿等。气候变化情境下,冻土融深由浅变深甚至部分岛状冻土消失,由空间序列代替时间序列,群落也由群丛组Ⅰ逐渐变化到群丛组Ⅱ和群丛组Ⅲ。 3.3 物种多样性对冻土融深的响应 目前就多样性与生境之间的关系已有大量报道,研究表明物种多样性与生境条件密不可分[26-29]。更有学者对生境条件对物种多样性影响进行了总结和综述[30- 33],指出了土壤水热条件对物种多样性有重要的作用,本研究发现,冻土的不同发育程度对土壤含水量有直接的影响,冻土融深小于50cm时土壤含水量较大,由于植物根系无法汲取土壤水分导致物种多样性指数较小,随着冻土融深的增加50cm [1] Wang G X, Li S N, Hu H C, Li Y S. Water regime shifts in the active soil layer of the Qinghai-Tibet Plateau permafrost region, under different levels of vegetation. Geoderma, 2009, 149(3/4): 280- 289. [2] 金会军, 于少鹏, 吕兰芝, 郭东信, 李英武. 大小兴安岭多年冻土退化及其趋势初步评估. 冰川冻土, 2006, 28(4): 467- 475. [3] Torre Jorgenson M, Racine C H, Walters J C, Osterkamp T E. Permafrost degradation and ecological changes associated with a warming climate in central Alaska. Climatic Change, 2001, 48(4): 551- 579. [4] 鲁国威, 翁炳林, 郭东信. 中国东北部多年冻土的地理南界. 冰川冻土, 1993, 15(2): 214- 218. [5] 顾钟炜, 周幼吾. 气候变暖和人为扰动对大兴安岭北坡多年冻土的影响-以阿木尔地区为例. 地理学报, 1994, 49(2): 182- 187. [6] 常晓丽, 金会军, 何瑞霞, 杨思忠, 于少鹏, 吕兰芝, 郭东信, 王绍令, 康兴成. 中国东北大兴安岭多年冻土与寒区环境考察和研究进展. 冰川冻土, 2008, 30(1): 176- 182. [7] Bhiry N, Payette S, Robert É C. Peatland development at the arctic tree line (Québec, Canada) influenced by flooding and permafrost. Quaternary Research, 2007, 67(3): 426- 437. [8] Lloyd A H, Yoshikawa K, Fastie C L, Hinzman L, Fraver M. Effects of permafrost degradation on woody vegetation at arctic treeline on the Seward Peninsula, Alaska. Permafrost and Periglacial Processes, 2003, 14(2): 93- 101. [9] Schuur E A, Crummer K G, Vogel J G, Mack M C. Plant species composition and productivity following permafrost thaw and thermokarst in Alaskan tundra. Ecosystems, 2007, 10(2): 280- 292. [10] Bakalin V A, Vetrova V P. Vegetation-Permafrost relationships in the zone of sporadic permafrost distribution in the Kamchatka Peninsula. Russian Journal of Ecology, 2008, 39(5): 318- 326. [11] Nguyen T N, Burn C R, King D J, Smith S L. Estimating the extent of near-surface permafrost using remote sensing, Mackenzie Delta, Northwest Territories. Permafrost and Periglacial Processes, 2009, 20(2): 141- 153. [12] Wania R, Ross I, Prentice I C. Integrating peatlands and permafrost into a dynamic global vegetation model: 1. Evaluation and sensitivity of physical land surface processes. Global Biogeochemical Cycles, 2009, 23(3): GB3014. [13] Kokfelt U, Rosén P, Schoning K, Christensen T R, Förster J, Karlsson J, Reuss N, Rundgren M, Callaghan T V, Jonasson C, Hammarlund D. Ecosystem responses to increased precipitation and permafrost decay in subarctic Sweden inferred from peat and lake sediments. Global Change Biology, 2009, 15(7): 1652- 1663. [14] Sannel A B K, Kuhry P. Long-term stability of permafrost in subarctic peat plateaus, west-central Canada. The Holocene, 2008, 18(4): 589- 601. [15] 张武文, 马秀珍, 丁国栋, 崔向新. 大兴安岭水湿地改造对土壤和植被的影响. 东北林业大学学报, 1996, 24(6): 9- 13. [16] 牟长城, 孙晓新, 倪志英, 杨明, 张娜. 大兴安岭林区沼泽_森林人工造林效果综合评估. 林业科学, 2007, 43(10): 51- 58. [17] 庄凯勋, 侯武才. 大兴安岭东部国有林区的湿地资源现状及保护对策. 东北林业大学学报, 2006, 34(1): 83- 86. [18] 吕久俊, 李秀珍, 胡远满, 王宪伟, 孙菊. 冻结数模型在中国东北多年冻土分区中的应用. 应用生态学报, 2008, 19(10): 2271- 2276. [19] 周梅, 余新晓, 冯林, 王林和, 那平山. 大兴安岭林区冻土及湿地对生态环境的作用. 北京林业大学学报, 2003, 25(6): 91- 93. [20] Tan Y Y, Wang X, Yang Z F, Wang Y L. Research progress in cold region wetlands, China. Sciences in Cold and Arid Regions, 2011, 3(5): 441- 447. [21] Hill M O. DECORANA: A FORTRAN Program for Detrended Correspondence Analysis and Reciprocal Averaging[D]. New York: Cornell University, 1979. [22] 马克平, 刘玉明. 生物群落多样性的测度方法Iα多样性的测度方法(下). 生物多样性, 1994, 2(4): 231- 239. [23] 刘庆仁, 孙振昆, 崔永生, 刘俊德, 程东方. 大兴安岭林区多年冻土与植被分布规律研究. 冰川冻土, 1993,15(2): 246- 251. [24] 孙菊, 李秀珍, 胡远满, 王宪伟, 吕久俊, 李宗梅, 陈宏伟. 大兴安岭沟谷冻土湿地植物群落分类、 物种多样性和物种分布梯度. 应用生态学报, 2009, 20(9): 2049- 2056. [25] 孙菊, 李秀珍, 王宪伟, 吕久俊, 李宗梅, 胡远满. 大兴安岭冻土湿地植物群落结构的环境梯度分析. 植物生态学报, 2010, 34(10): 1165- 1173. [26] Davies R G, Orme C D L, Webster A J, Jones K E, Blackburn T M, Gaston K J. Environmental predictors of global parrot (Aves: Psittaciformes) species richness and phylogenetic diversity. Global Ecology and Biogeography, 2007, 16(2): 220- 233. [27] Humphries C J, Densham P J, Lampinen R, Hagemeijer W J M, Mitchell-Jones A J, Gasc J P. Would environmental diversity be a good surrogate for species diversity? Ecography, 2001, 24(1): 103- 110. [28] Keller A, Rödel M O, Eduard Linsenmair K, Ulmar Grafe T. The importance of environmental heterogeneity for species diversity and assemblage structure in Bornean stream frogs. Journal of Animal Ecology, 2009, 78(2): 305- 314. [29] Trakhtenbrot A, Kadmon R. Effectiveness of environmental cluster analysis in representing regional species diversity. Conservation Biology, 2006, 20(4): 1087- 1098. [30] Dai Y C, Yang Z L, Cui B K, Yu C J, Zhou L W. Species diversity and utilization of medicinal mushrooms and fungi in China (Review). International Journal of Medicinal Mushrooms, 2009, 11(3): 287- 302. [31] Levin L A, Etter R J, Rex M A, Gooday A J, Smith C R, Pineda J, Stuart C T, Hessler R R, Pawson D. Environmental influences on regional deep-sea species diversity. Annual Review of Ecology and Systematics, 2001, 32(1): 51- 93. [32] 李禄军, 曾德慧. 物种多样性与生态系统功能的关系研究进展. 生态学杂志, 2008, 27(11): 2010- 2017. [33] Orlov N L, Nguyen S N, Ho C T. Description of a new species and new records of Rhacophorus genus (Amphibia: Anura: Rhacophoridae) with the review of amphibians and reptiles diversity of Ghu Yang Sin National Park (Dac Lac Province, Vietnam). Russian Journal of Herpetology, 2008, 15(1): 67- 84. [34] Trombulak S C, Frissell C A. Review of ecological effects of roads on terrestrial and aquatic communities. Conservation Biology, 2000, 14(1): 18- 30. [35] Kemper J T, Macdonald S E. Effects of contemporary winter seismic exploration on low Arctic plant communities and permafrost. Arctic, Antarctic, and Alpine Research, 2009, 41(2): 228- 237. [36] Kokelj S V, Zajdlik B, Thompson M S. The impacts of thawing permafrost on the chemistry of lakes across the subarctic boreal-tundra transition, Mackenzie Delta region, Canada. Permafrost and Periglacial Processes, 2009, 20(2): 185- 199. Classification of plant communities and species diversity responses to changes in the permafrost depths of the north slope of the Great Khingan Mountain valley of Northeast China GUO Jinting1,2,HAN Fenglin1,2,BU Rencang1,*,HU Yuanman1, MA Jun1,2,NIE Zhiwen1,2, ZHU Julan3,REN Baihui1,2 1StateKeyLaboratoryofForestandSoilEcology,InstituteofAppliedEcology,ChineseAcademyofSciences,Shenyang110016,China2UniversityofChineseAcademyofSciences,Beijing100049,China3Shenyangnormaluniversity,Shenyang110034,China The Great Khingan Mountains, located in northeastern China, feature zonal permafrost. Lying on the southern edge of the Eurasian permafrost boundary, the permafrost region of the northeast portion of the Great Khingan Mountains is one of the plant communities most sensitive to the effects of global warming. Permafrost is gradually degrading from south to north, because of the effects of human activity and environmental changes such as global warming. The degradation of permafrost leads to changes in the characteristics of permafrost plant communities, such as species composition, species diversity, and biomass. The impacts of permafrost on vegetation have become a key research topic in the field of climate change research. However, previous studies of permafrost have mostly focused on the distribution of permafrost, and so the mechanistic connection between permafrost and environmental factors, and the impacts of permafrost degradation on community composition and structure have not been the focus of much quantitative research. Thus, the plant communities of 30 plots on the north slope of the Great Khingan Mountains were classified according to permafrost depth, using two-way indicator species analysis (TWINSPAN) and canonical correspondence analysis (CCA). Accordingly, the effects of permafrost depth on plant species diversity were analyzed using these plots. The results indicate the following. (1) Plant communities were categorized into three plant associations, and as the permafrost depths changed along a gradient from shallow to deep, plant associations changed fromBetulafruticose+Ledumpalustrevar.angustum-CarexsubpediformistoBetulafruticosa-CarexsubpediformisandBetulafruticosa+Salixrosmarinifolia-Carexsubpediformis. These relationships were validated by CCA ordination. In addition, permafrost melting depth (PMD), topography, and soil moisture could together explain 38% of the community change. Their combined effect was the biggest explanatory factor for community change. (2) The species diversity index increased trend initially (in shallow permafrost) and decreased as the permafrost melting depth increased. It reached its maximum value at about 50—150 cm deep. When PMD ≤ 50 cm and PMD > 150 cm, the species richness and diversity index are low, and the significant difference. Patrick index and melting permafrost depth has significant correlation (R2= 0.58, P < 0.58). When PMD ≤ 50 cm, the species richness of the communities was higher, at 23.83±2.44; when 50 < PMD ≥ 150 cm, it was 26.36±2.01; and when PMD > 150 cm, it fell sharply to 21.14±1.57. The Pielou index and Shannon-Wiener index with the increase of the permafrost melts deep appeared a trend of reducing the rise, and when 50 The Great Khingan Mountains;permafrost deeps;species diversity;TWINSPAN;CCA 国家重点基础研究发展计划(2013CBA01807);中国科学院先导专项(XDA05050200);国家自然科学基金项目(41371198) 2015- 04- 30; 日期:2016- 03- 02 10.5846/stxb201504300895 *通讯作者Corresponding author.E-mail: burc@iae.ac.cn 郭金停,韩风林,布仁仓,胡远满,马俊,聂志文,朱菊兰,任百慧.大兴安岭北坡多年冻土区植物群落分类及其物种多样性对冻土融深变化的响应.生态学报,2016,36(21):6834- 6841. Guo J T,Han F L,Bu R C,Hu Y M, Ma J,Nie Z W, Zhu J L,Ren B H.Classification of plant communities and species diversity responses to changes in the permafrost depths of the north slope of the Great Khingan Mountain valley of Northeast China.Acta Ecologica Sinica,2016,36(21):6834- 6841.1 研究地区与研究方法


2 结果与分析



3 讨论
猜你喜欢
东北林业大学学报(2023年2期)2023-02-22
水土保持研究(2022年1期)2022-12-27
内蒙古师范大学学报(自然科学汉文版)(2022年1期)2022-01-10
草地学报(2021年6期)2021-07-05
铁道学报(2018年5期)2018-06-21
水利科技与经济(2016年8期)2016-04-22
中国铁道科学(2015年6期)2015-06-21
山西大学学报(自然科学版)(2014年2期)2014-05-10
浙江农林大学学报(2014年2期)2014-04-26
草业科学(2012年5期)2012-03-13
