
激活核受体PXR促进小鼠GM-CSF来源的树突状细胞的分化①
2016-12-26 01:46邱怀娜查贺飞曲佳乐齐艳伟
中国免疫学杂志 2016年11期
邱怀娜 查贺飞 曲佳乐 王 梅 李 璐 齐艳伟 黄 俊 杨 权
(广州医科大学基础学院病原生物学与免疫学教研室,广州医科大学中法霍夫曼免疫研究所,广州511436)
激活核受体PXR促进小鼠GM-CSF来源的树突状细胞的分化①
邱怀娜 查贺飞 曲佳乐 王 梅 李 璐 齐艳伟 黄 俊 杨 权
(广州医科大学基础学院病原生物学与免疫学教研室,广州医科大学中法霍夫曼免疫研究所,广州511436)
目的:研究孕烷受体(PXR)激动剂pregnane-16α-carbonitrile(PCN)对小鼠GM-CSF来源树突状细胞(DC)分化的影响。方法:建立小鼠DC体外GM-CSF和IL-4诱导分化体系,并用PCN对体系进行处理。流式细胞术检测CD11c+DC的比例;ELISA和流式细胞术检测DC产生细胞因子IFN-γ、IL-1、IL-2和IL-12的量;实时荧光定量聚合酶链反应(Real-time RT-PCR)检测调控DC分化相关信号通路基因的表达。结果:成功建立小鼠DC体外分化体系;PCN处理能显著促进DC分化,且DC产生的细胞因子IFN-γ和IL-2的量显著增多;Real-time RT-PCR检测结果表明,与对照组相比,PCN处理组中调控DC分化的Notch和Wnt信号通路相关基因HES1、WISP1和WISP2的表达量显著上升。结论:PXR激动剂PCN能显著促进小鼠GM-CSF来源的DC的分化及其IFN-γ和IL-2的产量,并且这种促进作用可能是通过Notch和Wnt信号通路进行调控。
树突状细胞;分化;Pregnane X Receptor;Notch信号通路;Wnt信号通路
树突状细胞(Dendritic cell,DC)是由Steinman 于1973年首次在小鼠脾脏中发现,因其具有特殊的星形形态学结构而得名[1,2]。DC是机体内功能最强大的专职抗原提呈细胞,能通过模式识别受体识别外来抗原,并迁移到外周淋巴器官将经其加工后的抗原提呈给T细胞,从而激活初始T细胞,启动抗原特异性的获得性免疫反应[3,4]。可以说DC是免疫系统的多能调控者,是连接固有免疫和获得性免疫反应的重要因子[5]。研究已证实,DC的分化受复杂的信号通路网络调控,其中包括细胞因子和转录因子等[6,7]。近期的研究表明,脂质信号通路在DC分化过程中起到重要的作用,核受体(Nuc-lear receptor)调控的脂类物质与DC分化调控基因表达的变化相关[8]。孕烷受体(Pregnane X receptor,PXR)是核受体超家族中的一员,高表达于肝脏和肠道,低表达于许多其他组织,调节葡萄糖、脂质和胆固醇众多物质的体内代谢平衡[9]。PXR的激活能诱导药物代谢酶和相关转运蛋白的表达,在药物代谢、肝脏解毒、肠道炎症以及肿瘤耐药性方面具有重要作用[10]。PXR同时表达于免疫细胞中[11],并具有免疫调节和抗炎症作用,调节免疫细胞的增殖和功能[12-14]。但是目前PXR在DC分化过程中的作用尚无报道。为此,本研究通过使用PXR激动剂PCN处理DC[15],检测诱导体系中CD11c+DC的比例、细胞因子分泌量以及调控DC分化的信号通路相关基因的表达,阐述PXR对小鼠DC分化的影响。
1 材料与方法
1.1 材料 6~8周SPF级雌性C57BL/6小鼠,由广东省实验动物中心提供。PBS缓冲液、RPMI1640基础培养基、胎牛血清(FBS)、β-巯基乙醇和TRIzol购自Invitrogen公司;红细胞裂解液、青霉素、链霉素、氯仿、异丙醇和乙醇购自生工生物工程(上海)股份有限公司;小鼠 GM-CSF和IL-4购自Peprotech公司;PCN和LPS购自Sigma公司;PrimeScript® RT reagent Kit和SYBR® Premix Ex TaqTM购自TaKaRa公司;Real-time RT-RCR引物由Invitrogen公司合成;PE-Cy5标记的抗小鼠CD11c、PE标记的抗小鼠IFN-γ、FITC标记的抗小鼠IL-2流式抗体购自eBioscience公司;ELISA试剂盒购自BD公司;倒置生物显微镜购自Leica Microsystems公司;CO2培养箱和Nanodrop2000c仪购自Thermofisher scientific公司;FACS Calibur流式细胞检测仪购自Beckman Coulter公司;S1000 PCR仪和CFX96 Real-time PCR仪购自Bio-Rad公司;酶标仪购自Promega公司。
1.2 方法
1.2.1 小鼠DC体外分化诱导 将小鼠快速颈椎脱臼处死,于75%的消毒酒精中浸泡5 min,取胫骨和股骨。将胫骨和股骨两端轻微剪开,用5 ml注射器吸取适量RPMI1640 培养基,插入骨髓腔,将骨髓细胞完全冲出,得到骨髓细胞悬液;3 500 r/min离心5 min,弃上清;加入3 ml红细胞裂解液重悬细胞,室温裂解5 min,加入27 ml PBS缓冲液终止裂解;3 500 r/min离心5 min,弃上清;加入适量RPMI1640 培养基重悬细胞,计数;按细胞数量计算接种孔数,每孔2×106细胞,2 ml培养基;按比例加入各种培养因子,使终浓度为:10%胎牛血清(FBS)、1%双抗(链霉素和青霉素)、50 μmol/L 2-巯基乙醇、20 ng/ml GM-CSF和10 ng/ml IL-4;用RPMI1640 培养基将细胞悬液浓度调整至1×106cells/ml,混匀,接种培养;每隔3 d更换新鲜培养液:吸弃1.8 ml旧培养液,加入2 ml含各种培养因子的新的完全培养液。
1.2.2 药物处理DC 分为对照组、PCN 20 μmol/L处理组、PCN 40 μmol/L处理组。细胞接种24 h内加药处理细胞,处理组:加入2 μl溶剂(DMSO),轻轻混匀;PCN 20 μmol/L处理组:加入2μl的PCN 20 mmol/L储存液,轻轻混匀;PCN 40 μmol/L处理组:加入2 μl的PCN 40 mmol/L储存液,轻轻混匀;更换培养液的同时重新加药处理细胞。对于细胞因子检测实验,增加LPS刺激细胞:DC诱导第5天,各实验组均加入100 ng/ml的LPS处理24 h。
1.2.3 Real-time RT-RCR检测基因表达 根据实验需要,收集相应时间点的2×106个待检测细胞,提取总的RNA并测定浓度和纯度,每个实验组各取100 ng RNA进行逆转录;使用PrimeScript® RT reagent Kit进行逆转录,操作步骤严格按照试剂盒说明书进行;使用SYBR® Premix Ex TaqTM进行Real-time PCR,操作步骤严格按照试剂盒说明书进行。各检测基因引物序列如表1所示,选用β-actin作为内参基因。
1.2.4 流式细胞术检测DC表型 细胞接种12 h,加入PCN对细胞进行处理。DC诱导第3天,更换新鲜的完全培养基,同时加入相应浓度的PCN处理细胞;DC诱导第6天,收集1×106个待检测细胞,3 500 r/min离心5 min,弃上清;按说明书用量将适量抗体加入100 μl PBS中,混匀,加入细胞沉淀中,涡旋震荡15 s;4℃避光孵育30 min;加入3 ml PBS缓冲液终止染色 ,3 500 r/min离心5 min,弃上清;重复洗涤细胞1次;细胞沉淀中加入500 μl含2%多聚甲醛的PBS缓冲液重悬细胞,涡旋混匀;流式细胞仪检测,读取50 000个细胞,结果在CellQuest (Becton Dickinson,Mountain View,CA)软件上进行分析。
表1 Real-time RT-PCR 引物序列
Tab.1 Primers for Real-time RT-PCR detection

GeneForwardprimer(5′⁃3′)Reverseprimer(5′⁃3′)Productsize(bp)CYP3A11CACACTTTCCTTCACCCTGCTATCATACGTGGGAGGTGCC110HES1GAGTGCATGAACGAGGTGACCGTTGATCTGGGTCATGCAG109WISP1AGCCTACACTAGTCCCTGGATCTTCCCTGCCTTGATGTGT164WISP2CCAGGAGAATACAGGTGCCAAGGAGTGACAAGGGCAGAAA122β⁃actinGTGGGAATGGGTCAGAAGGACTTCTCCATGTCGTCCCAGT120
1.2.5 培养上清细胞因子含量检测 细胞接种12 h,加入PCN对细胞进行处理。DC诱导第3天,更换新鲜的完全培养基,同时加入相应浓度的PCN处理细胞;DC诱导第5天,加入LPS处理细胞。LPS处理24 h后,收集待检测细胞培养上清,采用ELISA试剂盒检测相应细胞因子的含量,操作步骤严格按照说明书进行,根据标准曲线用间接法计算各实验组中相应细胞因子的含量(pg/ml)。
1.2.6 流式胞内因子染色 细胞接种12 h,加入PCN对细胞进行处理。DC诱导第3天,更换新鲜的完全培养基,同时加入相应浓度的PCN处理细胞;DC诱导第5天,加入LPS处理细胞。LPS处理24 h后,收集待检测细胞。将细胞终浓度调至2×106ml,加入佛波醇-乙酸酯(PMA) 20 ng/ml,离子霉素1 μg/ml,涡旋振荡混匀。37℃,5%CO2培养箱中孵育1 h后,加布雷菲德菌素A(BFA)10 μg/ml继续孵育4 h。加入2 ml PBS, 4℃、1 860 r/min离心5 min。重复2次。PBS重悬细胞,每管加入2 ml 4%的多聚甲醛,充分混合,避光静置8 min。PBS洗涤,4℃、2 500 r/min离心5 min。每管加入细胞因子染色液(含Saponin)100 μl,4℃静置过夜。加入相应荧光标记抗体各1 μl/每管,充分混匀,4℃避光静置30 min。细胞因子染色液(不含Saponin)2 ml,2 500 r/min离心5 min,重复2次。用细胞因子染色液重悬细胞,流式细胞仪检测,读取50 000个细胞。

2 结果
2.1 DC分化过程中PXR靶基因的表达 接种后,收集第0、3、6天的细胞, Real-time RT-RCR检测PXR靶基因CYP3A11的表达。与第0天相比,诱导培养3 d后的细胞中,CYP3A11的表达量显著上调(1.2±0.23)与(5.96±0.88),差异具有统计学意义(P<0.05,图1);与第3天相比,诱导培养6 d后的细胞中,CYP3A11的表达量显著上调(5.96±0.88)与(14.65±1.4),差异具有统计学意义(P<0.01,图1)。说明在小鼠骨髓细胞向DC分化的过程中,PXR处于激活状态,其靶基因表达在分化过程中表达显著上调。
2.2 PXR激动剂PCN促进DC的分化 细胞接种12 h,加入PCN处理细胞。倒置显微镜下观察,细胞生长状态良好,说明各实验组浓度药物未产生细胞毒作用。收集药物处理第0天、第3天和第6天的细胞进行Real-time RT-RCR检测,结果表明,与对照组相比,PCN 20 μmol/L处理组和PCN 40 μmol/L处理组的细胞中CYP3A11的表达在DC分化过程中均显著上调(P<0.01,P<0.05,图2),说明PCN成功激活了PXR。流式细胞术检测培养体系中CD11c+DC的比例:对照组CD11c+DC细胞的比例为(49.48±4.1)%;PCN 20 μmol/L处理组CD11c+DC细胞的比例为(59.2±7.86)%;PCN40 μmol/L处理组CD11c+DC细胞的比例为(71.25±4.2)%(图2)。结果表明,PXR激动剂PCN处理能显著促进培养体系中DC的分化,提高CD11c+DC细胞的比例,差异具有统计学意义(P<0.05),并且这种促进作用具有浓度依赖性。
2.3 PXR激动剂PCN促进DC产生细胞因子IFN-γ和IL-2 ELISA检测结果表明,与对照组相比,PCN 40 μmol/L处理组DC培养上清中细胞因子IFN-γ和IL-2的含量显著上升,差异具有统计学意义(P<0.01、P<0.05,图3A);而细胞因子IL-1和IL-12的含量在对照组和PCN 40 μmol/L处理组中无明显变化,差异不具有统计学意义(P>0.05,图3A)。流式胞内因子染色检测结果表明,与对照组相比,PCN 40 μmol/L处理组的CD11c+DC细胞内,细胞因子IFN-γ和IL-2的比例显著上升,差异具有统计学意义(P<0.01、P<0.05,图3B)。上述结果表明,PXR激动剂PCN处理能显著促进培养体系中的CD11c+DC细胞产生细胞因子IFN-γ和IL-2。

图1 Real-time RT-RCR检测DC分化过程中CYP3A11的表达Fig.1 Real-time RT-RCR detecting expression of CYP3A11 gene during DC differentiationNote: * .P<0.05 vs 0 d;**.P<0.01 vs 3 d.

图2 PCN处理后培养体系中CYP3A11的表达和CD11c+ DC细胞的比例Fig.2 Expression of CYP3A11 gene and proportion of CD11c+ DC were measured after PCN treatmentNote: *.P<0.05 vs control group;**.P<0.01 vs control group.

图3 ELISA和流式细胞术检测细胞因子IFN-γ、IL-1、IL-2和IL-12的含量Fig.3 Protein levels of IFN-γ,IL-1,IL-2 and IL-12 were measured by ELISA and flow cytometryNote: *.P<0.05 vs control group;**.P<0.01 vs control group;NS.No significance.
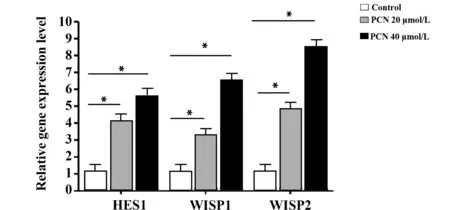
图4 Real-time RT-RCR检测PCN处理的DC中HES1、WISP1和WISP2的表达Fig.4 Real-time RT-RCR detecting expression of HES1,WISP1 and WISP2 in DC treated with PCNNote: *.P<0.05 vs control group.
2.4 PXR激动剂PCN促进培养体系中HES1、WISP1和WISP2的表达 PCN处理24 h后,Real-time RT-RCR检测调控DC分化的Notch 和Wnt信号通路相关基因HES1、WISP1和WISP2的表达。与对照组相比,PCN 20 μmol/L处理组和PCN 40 μmol/L处理组细胞中HES1、WISP1和WISP2的表达显著上调,差异具有统计学意义(P<0.05,图4)。结果表明, PCN处理能显著促进细胞中Notch 和Wnt信号通路相关基因的表达,说明PCN对DC分化的调控作用可能与Notch 和Wnt信号通路的激活有关。
3 讨论
DC是介导固有免疫反应和启动获得性免疫反应的重要免疫细胞,能识别、加工和提呈外来抗原,同时维持免疫耐受,保证在正常生理情况下效应T细胞不会攻击自身正常细胞或组织产生的抗原[5,16]。研究已证实多种信号通路在DC分化过程中发挥重要的调控作用,其中包括Wnt、Notch、NF-κB以及JAK/STAT等[17-19]。PXR是核受体超家族中的一员,能被多种激动剂激活,从而对其功能进行研究。目前对PXR的研究主要集中在药物代谢、肝脏脂肪病变、肠道炎症、肥胖、心血管疾病以及肿瘤药物反应等领域[10,20,21],其在免疫系统中的调节作用研究甚少,对DC分化的影响尚未见相关报道。
Zhong等[22]报道核受体LXR(Liver X receptor)激动剂GW3965能通过抑制STAT3信号通路的磷酸化从而促进DC的分化[23]。更有研究报道PCN能诱导抗原提呈细胞中MHCⅡ基因家族成员RT1.B的表达。说明核受体家族在树突状细胞的分化过程中可能具有很重要的调控作用。本研究首先成功建立了小鼠骨髓细胞向DC分化的体外诱导体系,并用PXR激动剂PCN处理DC。药物处理后细胞生长状态良好,说明本研究使用的药物浓度并未产生细胞毒作用。本研究发现PXR低表达于小鼠骨髓细胞中(结果省略),并且在DC分化过程中PXR靶基因CYP3A11表达显著上调,说明在小鼠骨髓细胞向DC分化的过程中,PXR处于激活状态,其靶基因CYP3A11对于DC分化可能是一个必需的有利因子。随后的实验结果表明,诱导体系经PCN处理后,PXR靶基因CYP3A11表达上调,体系中CD11c+DC的比例显著上升,并且这种上调作用具有药物浓度依赖性。本研究结果首次证实了核受体家族中的另一成员PXR能调控小鼠GM-CSF来源的DC的分化。
本研究结果发现,PCN处理能显著促进DC功能相关细胞因子IFN-γ和IL-2的产量。IFN-γ和IL-2是DC分泌的重要细胞因子,可上调DC 中MHC Ⅰ类和Ⅱ类分子的表达,促进DC将抗原肽提呈给初始T细胞,促进T、B细胞增殖和分化,启动获得性免疫应答反应[24,25],同时诱导CTL和NK细胞的杀伤活性,在抗病毒和抗肿瘤免疫反应中具有重要作用[26]。有研究表明,PXR可调节免疫反应[13]。从本研究结果推测,PXR激动剂PCN通过促进诱导体系中DC产生IFN-γ和IL-2,而分泌的IFN-γ和IL-2可帮助DC激活多种免疫细胞,从而调节免疫系统反应活性。
本研究结果显示,PCN处理DC后,调控DC分化的Notch和Wnt信号通路相关基因HES1、WISP1和WISP2表达显著上调,说明Notch和Wnt处于激活状态。研究表明,Notch和Wnt信号通路在对DC的分化具有重要的调控作用[27];Zhou等[17]研究表明在造血祖细胞中激活Notch信号通路可促进DC的分化,而这种促进作用是由Wnt信号通路介导的。Becker等[28]的研究结果发现激活Wnt信号通路能促进DC的产生。由此推测,PXR激动剂PCN可能是通过激活Notch和Wnt信号通路从而促进DC的分化。
综上所述,本研究结果显示核受体超家族成员PXR的激动剂PCN能显著促进小鼠GM-CSF来源DC的分化,促进其产生细胞因子IFN-γ和IL-2,并且这种促进作用可能是通过激活Notch和Wnt信号通路的活性进行调控的。本研究结果为阐明DC分化机制,完善调控DC分化信号通路网络提供了新的线索;为PXR在免疫系统中的调控作用提供了新的理论依据;为疾病免疫治疗方案制定及新药研发提供了新的思路。
[1] Schraml BU,Reis ESC.Defining dendritic cells[J].Curr Opin Immunol,2015,32:13-20.
[2] Poltorak MP,Schraml BU.Fate mapping of dendritic cells[J].Front Immunol,2015,6:199.
[3] 强叶涛,吴水芸,韩慕天,等.丁酸对小鼠骨髓源树突状细胞的免疫调节作用[J].中国免疫学杂志,2015,31(10):1315-1319.
[4] Ganguly D,Haak S,Sisirak V,etal.The role of dendritic cells in autoimmunity[J].Nat Rev Immunol,2013,13(8):566-577.
[5] Mellman I.Dendritic cells:master regulators of the immune response[J].Cancer Immunol Res,2013,1(3):145-149.
[6] Gabrilovich D.Mechanisms and functional significance of tumour-induced dendritic-cell defects[J].Nat Rev Immunol,2004,4(12):941-952.
[7] Moretto MM,Lawlor EM,Khan IA.Aging mice exhibit a functional defect in mucosal dendritic cell response against an intracellular pathogen[J].J Immunol,2008,181(11):7977-7984.
[8] Nagy L,Szanto A,Szatmari I,etal.Nuclear hormone receptors enable macrophages and dendritic cells to sense their lipid environment and shape their immune response[J].Physiol Rev,2012,92(2):739-789.
[9] Ihunnah CA,Jiang M,Xie W.Nuclear receptor PXR,transcriptional circuits and metabolic relevance[J].Biochim Biophys Acta,2011,1812(8):956-963.
[10] Koutsounas I,Patsouris E,Theocharis S.Pregnane X receptor and human malignancy[J].Histol Histopathol,2013,28(4):405-420.
[11] Schote AB,Turner JD,Schiltz J,etal.Nuclear receptors in human immune cells:expression and correlations[J].Mol Immunol,2007,44(6):1436-1445.
[12] Hu G,Xu C,Staudinger JL.Pregnane X receptor is SUMOylated to repress the inflammatory response[J].J Pharmacol Exp Ther,2010,335(2):342-350.
[13] Mencarelli A,D′Amore C,Renga B,etal.Solomonsterol A,a marine pregnane-X-receptor agonist,attenuates inflammation and immune dysfunction in a mouse model of arthritis[J].Mar Drugs,2014,12(1):36-53.
[14] Dubrac S,Elentner A,Ebner S,etal.Modulation of T lymphocyte function by the pregnane X receptor[J].J Immunol,2010,184(6):2949-2957.
[15] Gahir SS,Piquette-Miller M.Gestational and pregnane X receptor-mediated regulation of placental ATP-binding cassette drug transporters in mice[J].Drug Metab Dispos,2011,39(3):465-471.
[16] 贾 原,李文丽,李 芳,等.维生素C对DC细胞功能调控影响研究[J].中国免疫学杂志,2015,31(10):1324-1328.
[17] Zhou J,Cheng P,Youn JI,etal.Notch and wingless signaling cooperate in regulation of dendritic cell differentiation[J].Immunity,2009,30(6):845-859.
[18] Platzer B,Jorgl A,Taschner S,etal.RelB regulates human dendritic cell subset development by promoting monocyte intermediates[J].Blood,2004,104(12):3655-3663.
[19] Onai N,Manz MG.The STATs on dendritic cell development[J].Immunity,2008,28(4):490-492.
[20] Zhou C.Novel functions of PXR in cardiometabolic disease[J].Biochim Biophys Acta,2016,1859(9):1112-1120.
[21] Habano W,Gamo T,Terashima J,etal.Involvement of promoter methylation in the regulation of Pregnane X receptor in colon cancer cells[J].Bmc Cancer,2011,11:81.
[22] Zhong L,Yang Q,Xie W,etal.Liver X receptor regulates mouse GM-CSF-derived dendritic cell differentiation in vitro[J].Mol Immunol,2014,60(1):32-43.
[23] Jimenez BD,Maldonado L,Dahl RH,etal.Ectopic expression of MHC class Ⅱ genes (RT1.B(I) beta/alpha) in rat hepatocytes in vivo and in culture can be elicited by treatment with the pregnane X receptor agonists pregnenolone 16 alpha-carbonitrile and dexamethasone[J].Life Sci,2002,71(3):311-323.
[24] Lee CC,Wang CC,Huang HM,etal.Ferulic acid induces Th1 responses by modulating the function of dendritic cells and ameliorates Th2-mediated allergic airway inflammation in mice[J].Evid Based Complement Alternat Med,2015,2015:678487.
[25] Herr F,Lemoine R,Gouilleux F,etal.IL-2 phosphorylates STAT5 to drive IFN-gamma production and activation of human dendritic cells[J].J Immunol,2014,192(12):5660-5670.
[26] 金伯泉.医学免疫学[M].第6版,北京:人民卫生出版社,2015:49-54.
[27] Cheng P,Zhou J,Gabrilovich D.Regulation of dendritic cell differentiation and function by Notch and Wnt pathways[J].Immunol Rev,2010,234(1):105-119.
[28] Becker MR,Choi YS,Millar SE,etal.Wnt signaling influences the development of murine epidermal Langerhans cells[J].J Invest Dermatol,2011,131(9):1861-1868.
[收稿2016-04-01 修回2016-05-16]
(编辑 倪 鹏)
Activation of PXR promotes differentiation of mouse GM-CSF derived dendritic cells
QIU Huai-Na,ZHA He-Fei,QU Jia-Le,WANG Mei,LI Lu,QI Yan-Wei,HUANG Jun,YANG Quan.
Department of Pathogen Biology & Immunology,School of Basic Sciences,Guangzhou Medical University;Sino-French Hoffmann Institute,Guangzhou Medical University,Guangzhou 511436,China
Objective:To investigate the influence of Pregnane X Receptor (PXR) agonist pregnane-16α-carbonitrile (PCN) on mouse GM-CSF derived dendritic cell (DC) differentiation.Methods: Mouse DC differentiation was induced by GM-CSF and IL-4 in vitro,and treated with PCN.The proportion of CD11c+DC was measured by flow cytometry;the production of Interferon-γ (IFN-γ),Interleukin-1 (IL-1),Interleukin-2 (IL-2) and Interleukin-12 (IL-12) of DC were analyzed by ELISA and flow cytometry;Real-time RT-RCR was used to detect the expression of genes involved in the signaling pathway which regulated DC differentiation.Results: Mouse DC was successfully induced by GM-CSF and IL-4 in vitro.Treatment of PCN significantly increased the percentage of CD11c+DC and the protein levels of IFN-γ and IL-2 produced by DC were notably enhanced.Real-time RT-RCR results indicated that the expression of Notch and Wnt signaling pathway related genes HES1,WISP1 and WISP2 were up-regulated in PCN treatment group when compared to control group.Conclusion: The PXR agonist PCN can promote the differentiation of mouse GM-CSF derived DC and increased the production of IFN-γ and IL-2 by DC,and Notch and Wnt signaling pathway may play a pivotal role in this process.
Dendritic cells;Differentiation;Pregnane X Receptor;Notch signaling pathway;Wnt signaling pathway
10.3969/j.issn.1000-484X.2016.11.006
①本文受广州医科大学科学科研项目资助(2015C01)。
邱怀娜(1983年-),女,博士,讲师,主要从事肿瘤分子免疫方面的研究,E-mail:autumn611@163.com。
及指导教师:杨 权(1985年-),男,博士,讲师,主要从事肿瘤分子免疫方面的研究,E-mail:yquangy20 15@163.com。
R392.12 R392.11
A
1000-484X(2016)11-1593-05
猜你喜欢
清华金融评论(2022年4期)2022-04-13
现代临床医学(2021年4期)2021-07-31
国际放射医学核医学杂志(2021年10期)2021-02-28
房地产导刊(2020年7期)2020-08-24
中成药(2018年10期)2018-10-26
天然产物研究与开发(2018年6期)2018-07-09
中西医结合心脑血管病杂志(2016年20期)2016-03-01
中国卫生标准管理(2015年18期)2016-01-20
医学研究杂志(2015年12期)2015-06-10
中国药理学通报(2014年2期)2014-05-09
