
miRNA和siRNA在hERG基因相关疾病中的研究进展
2016-12-26 01:57:49庄凯丽廉姜芳
浙江医学 2016年7期
庄凯丽 廉姜芳
●综 述
miRNA和siRNA在hERG基因相关疾病中的研究进展
庄凯丽 廉姜芳
1991年Warmke等首先从果蝇中克隆到EAG(ether-a-go-go)基因,其编码一种电压门控性钾通道,即EAG K+通道。EAG K+通道有3个亚家族,其中EAG相关基因(hERG)K+通道在人海马中被发现,其编码的K+通道称为hERG K+通道[1]。hERG主要在心脏中表达,编码快速激活延迟整流钾通道的a亚基,在动作电位复极化中发挥重要作用。至今其相关研究从未中断,并取得了一定的进展,尤其是在长QT综合征(KQTS)和肿瘤方面。1993年,Kee等[2]发现控制秀丽新小杆线虫发育的时序性的基因lin-4不编码蛋白质,其相应的mRNA水平没有发生变化。这一矛盾现象没有引起足够的重视。2000年,Reinhart等[3]在秀丽新小杆线虫中发现与lin-4类似的基因。自此,科学界开始了miRNA的时代。1999年,实验发现21~23ntRNA引发植物基因表达沉默[4]。进一步研究揭示这种RNAi由双链RNA转换而来。笔者就miRNA和siRNA在hERG基因相关疾病中的研究进展作一综述。
1 hERG
hERG编码快速激活延迟整流钾通道Ikr的α亚基,在心肌动作电位的复极化,消化、分泌、生殖系统的耦合收缩,神经系统的信号传导和人体的发育等方面起着重要作用[5]。hERG基因因突变、药物等因素可影响通道的转运,基因过表达或低表达以及异位过表达可出现相应的功能异常。
2 miRNA与siRNA
小RNA主要包括miRNA和siRNA两大类。此外,还有核糖体开关、反义核酸等非编码RNA。但以siRNA和miRNA研究最多见。miRNA是长度在19~25nt的调控蛋白质生物合成的一类重要的内源性非编码RNA。双链miRNA分子被解链,单链的miRNA主要与AGO结合形成RNA诱导的沉默复合体,然后可通过与靶mRNA的3′UTR近乎完全互补结合在转录后水平使其降解,或者与之不完全互补结合在翻译水平抑制蛋白合成,从而在基因表达中发挥重要的调节作用。人类基因编码的miRNA多达1 800多种,调控约60%的基因。miRNA在人类的生理、病理生理过程中通过其自身的复杂调控网络起着重要的作用。真核细胞没有特定的基因编码内源性siRNA,siRNA具有特定长度(21~25nt)和特定序列的小片段RNA。siRNA与miRNA相似,也主要与AGO2结合形成RNA诱导的沉默复合体,与靶序列mRNA完全互补,通过降解靶序列导致基因表达沉默。shRNA呈发卡状,是siRNA的前体。
3 miRNA、siRNA在hERG基因相关疾病的研究
hERG基因相关性疾病中,以KQTS和肿瘤研究最多,其它相关性疾病少有研究。在肿瘤的研究进展中,衍生出心脏-肿瘤学科[6-8]。
3.1 KQTS hERG基因突变或其参与编码的Ikr通道受到药物等因素影响后,心室复极延迟而引发Q-T间期延长,常并发室性心律失常,尤其是尖端扭转型室性心动过速(TdP)、晕厥,严重者可致心脏性猝死。
心肌miRNA的相关研究起步较晚且多是建立在骨骼肌研究基础之上。2002年,Kagos-Quintana等[9]发现miR-1和miR-133在心肌组织表达最丰富,且有一定的组织特异性,说明这两种miRNA对心肌细胞发育具有重要意义。Chen等[10]对此进行了验证。之后的研究揭示了miRNA与心肌肥大、心肌缺血、糖尿病性心肌病等不同病理发生、发展有着密切关系,且发现缺血性心肌病和糖尿病性心肌病中miR-133表达明显增高,抑制ERG和KCNQ1基因的表达,引发Q-T间期延长,但miR-133对糖尿病性心肌的KCNQ1抑制作用不明显[11]。Hedley等[12]发现miR-1-1和miR-133a-1基因序列没有发生变化。虽然miR-1-2有一个位点(n.100A>G)和miR-133a-2有两个位点(n.19G>A andn.98C>T)的改变,但不影响miRNA的成熟。可以看出,miR-1-1,miR-1-2,miR-133a-1和miR-133a-2序列变化与先天性KQTS的发病无明显关系。miR-133靶向小鼠的ERG基因,使Ikr通道功能丧失,Q-T间期延长,但是在hERG上还未发现其结合位点[13]。miR-133家族与miR-1的亚族共同调控hERG基因表达[14]。经As2O3诱导后,豚鼠心肌细胞的miR-1和miR-133及其反式激活血清应答因子表达水平均明显上调,导致ERG蛋白水平下降,Q-T间期延长[15]。慢性氧化应激过程中,miR-17-5p作用于多个内质网应激相关的伴侣Hsp70、Hsc70、CANX和Golga2,从而减少hERGK+通道蛋白的转运[16]。如图1所示。
siRNA序列靶向干预已转染WT-hERG(野生型)、WT/E637K-hERG(杂合型)和E637K-hERG(突变型)的HEK-293细胞,发现其能有效地沉默E637K突变体的表达,并且不影响干预前细胞的生长和凋亡。在通道电流方面,siRNA不仅增加了杂合型电流幅度,而且纠正了通道电流的特性,但对野生型通道的电流幅度和通道特性没有明显的影响[17]。基于siRNA此作用特点,其在KQTS未来基因治疗方面是很有前景的。Zarzoso等[18]就此项研究作出评论:小干扰RNA对于hERG K+通道转运的恢复是很有希望的一种基因治疗方案。RNA干扰伴侣分子FKBp38而降低其表达水平,随之hERG转运出现了下降[19]。诱导多功能干细胞来源的心肌细胞在心脏疾病方面的研究已较为成熟。经特异性RNA干扰携带KCNH2 c.G1681A突变基因的表达,心肌细胞的APD和K+电流可恢复正常[20]。如图1所示。

图1 miRNA和siRNA作用于心肌细胞ERG基因
3.2 肿瘤 由于肿瘤本质上是一种多基因异常疾病,通过激活一种或多种原癌基因的表达及抑癌基因的突变缺失从而使肿瘤细胞逃避了正常生长调控机制,自主进行增殖和侵袭,出现恶性表型。hERG的癌症组织中过表达,如肾癌[21]、神经细胞瘤[22]、白血病[23]和乳腺癌[24],与癌细胞的增殖、凋亡、分化、发病和转移有密切的关系[25]。阻碍通道的生成或抑制其转运成为癌症治疗的一个新途径。
Calin等[26]于2002年首次对miRNA与肿瘤的关系作出报道,此后人们发现miRNA在多种肿瘤中特异表达,并且在肿瘤的增殖、分化、侵袭转移、治疗效果中扮演了重要角色,在肿瘤的早期诊断及治疗中可能具有重要意义。随着对miRNA研究的深入,发现它们具有调节细胞增殖分化和凋亡的功能。这些效应通过调控信号分子,如细胞因子、转录因子、生长因子、促凋亡和抗凋亡基因的表达而实现[27]。这预示着miRNA与肿瘤的发生、发展和预后存在着密切的联系。多项研究证实,某些miRNA在部分肿瘤中特异性高表达。如果它们的被抑靶基因是抑癌基因,其升高可以刺激肿瘤增殖、血管生成、侵袭转移等,则这些miRNA扮演了癌基因角色[28]。另有证据提示miRNA尚可扮演抑癌基因的角色,不仅多个miRNA被发现在肿瘤中表达降低,而且通过抑制Drosha而降低miRNA生成时发现肿瘤细胞的恶性程度降低[29]。hERG1控性钾离子通道的EAG家族成员之一,其编码的钾通道是一种特殊类型的钾通道,具有内向整流特性。研究发现hERG1基因在胰腺癌组织及细胞中高表达,并与胰腺癌细胞的增殖、转移等生物活性密切相关,但其在肿瘤中的调控机制方面的作用尚不明确。miR-96可下调hERG1的表达,同时胰腺癌细胞增殖、转移和侵袭方面也显著受到了抑制。针对miR-96/herg1的基因靶向治疗在胰腺癌的治疗中有潜在的应用前景[30]。多数急性髓细胞白血病异常表达miR-9/9,后者通过下调ERG基因表达来干扰正常的中性粒细胞分化[31]。TMPRSS2-ERG癌基因存在于50%以上前列腺癌患者,其表达预示预后不良。TMPRSS2-ERG基因融合诱发前列腺肿瘤,研究发现miR-200C参与此病理生理过程[32]。此外,miR-145可以调控前列腺癌原癌基因ERG的转录后水平来抑制疾病的发展进程[33]。如图2所示。
siRNA在肿瘤基因治疗方面已有很多的研究[34],但是siRNA的细胞毒性和低转染率阻碍了其临床治疗方面的研究价值。Whitehead等[35]研究发现PAMAM-PEG-cRGD作为siRNA靶向hERG基因的载体,不仅在很大一定程度上提高了转染效率,同时所产生细胞的毒性也是极低的,几乎可以忽略。这对于其在hERG基因的小干扰RNA方面研究和未来的临床应用都具有一定的意义。在一些研究中,小干扰RNA作为沉默靶基因的实验方法。小干扰RNA转染至胰腺癌细胞株PANC-1与CFPAC-1中,稳定沉默hERG1基因[30]。hERG1基因在胃癌组织过表达,但在正常的黏膜上皮组织不表达,而且还发现其表达与癌细胞的分化和临床分期相关联[36-37]。顺铂是临床常用的化疗药物,对于其化疗机制是否与hERG基因编码的钾通道有关,Zhang等[38]利用siRNA干扰作用靶向抑制hERG基因的表达与联合顺铂进行研究。结果显示hERG与顺铂诱导的细胞凋亡有着密切的相关性,对于推测hERG可能是顺铂化疗的靶目标提供了有力的实验依据。在胶质母细胞瘤的细胞周期停滞和凋亡与hERG基因相关性研究中,同样用小干扰RNA可抑制hERG基因表达[39]。小细胞型肺癌4个细胞系(SW2、OH1、OH3、H82)均有herg1的表达,在研究其在肿瘤细胞增殖、转移等机制时发现siRNA与ERG通道特异性阻滞剂E-4031效应不完全不同,抑制hERG1蛋白的表达后癌细胞的增殖减少近50%,通道电流几乎消失。而E-4031虽然改变通道电流,但并不影响癌细胞的增殖[40]。Urbinati等[41]发现siRNA能够有效地抑制TMPRSS2-ERG癌基因的表达,有明显的抗肿瘤效应。shRNA与成神经细胞瘤hERG基因结合,使其停滞在G0/G1期,延缓细胞增长速度,降低细胞活性,抑制集落形成。在裸鼠的肿瘤细胞中导入shRNA,肿瘤受到了抑制[42]。shRNA沉默herg基因表达对于延缓肿瘤的增长具有一定的临床治疗意义[43]。如图2所示。
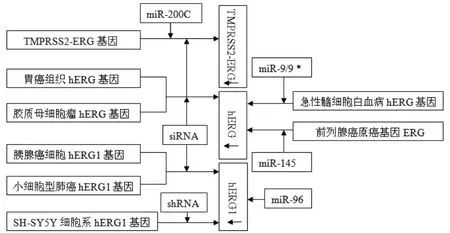
图2 miRNA和siRNA作用于肿瘤hERG/hERG1/TMPRSS2-ERG基因
3.3 其它疾病 hERG基因除心脏和肿瘤组织外也有表达,但在如消化、内分泌等系统相关的miRNA和siRNA研究甚少,故不作叙述。
4 展望
hERG基因表达异常引发室性心律失常,严重者可致患者死亡,其临床诊断困难,治疗效果有限。基因诊断和治疗或许可以为临床开辟一条新出路。miRNA在 hERG基因相关疾病主要扮演病理性的角色,过度上调或下调均可引起离子通道的紊乱。miRNA作为诊疗标志物应用于临床具有一定的前景;shRNA和siRNA其作用机制为降解靶基因hERG基因片段。与miRNA相比,其基因治疗效果更明显。但是机体有着复杂的网络信号通路,miRNA和干扰RNA基因治疗不可避免地干扰其它通道的正常功能。而其如何安全有效地导入靶器官成为基因治疗的又一大挑战。
[1] Warmke J W,Ganetzky B.A family of potassium channel genes related to eag in Drosophila and mammals[J].Proc Natl Acad Sci U S A,1994,91(8):3438-3442.
[2] Lee R C,Feinbaum R L,Ambros V.The C.elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14[J].Cell,1993,75(5):843-854.
[3] Reinhart B J,Slack F J,Basson M,et al.The 21-nucleotide let-7 RNA regulates developmental timing in Caenorhabditis elegans [J].Nature,2000,403(6772):901-906.
[4] Tomari Y,Zamore P D.Perspective:machines for RNAi[J].Genes Dev,2005,19(5):517-529.
[5] Babcock J J,Li M.hERG channel function:beyond long QT[J].Acta Pharmacologica Sinica,2013,34(3):329-335.
[6] Conti E,Romiti A,Musumeci M B,et al.Arterial thrombotic events and acute coronary syndromes with cancer drugs:are growth factors the missed link?:what both cardiologist and oncologist should know about novel angiogenesis inhibitors[J].Int J Cardiol, 2013,167(6):2421-2429.
[7] Rateesh S,Luis S A,Luis C R,et al.Myocardial infarction secondary to 5-fluorouracil:not an absolute contraindication to rechallenge?[J].Int J Cardiol,2014,172(2):e331-333.
[8] Kurisu S,Iwasaki T,Ishibashi K,et al.Comparison of treatment and outcome of acute myocardial infarction between cancer patientsandnon-cancerpatients[J].IntJCardiol,2013,167(5):2335-2337.
[9] Lagos-Quintana M,Rauhut R,Yalcin A,et al.Identification of tissue-specific microRNAs from mouse[J].Curr Biol,2002,12(9): 735-739.
[10] Chen J F,Mandel E M,Thomson J M,et al.The role of microRNA-1 and microRNA-133 in skeletal muscle proliferation and differentiation[J].Nat Genet,2006,38(2):228-233.
[11] Zhang Y,Xiao J,Lin H,et al.Ionic mechanisms underlying abnormal QT prolongation and the associated arrhythmias in diabetic rabbits:a role of rapid delayed rectifier K+current[J].Cell Physiol Biochem,2007,19(5-6):225-238.
[12] Hedley P L,Carlsen A L,Christiansen K M,et al.MicroRNAs in car diac arrhythmia:DNA sequence variation of MiR-1 and MiR-133A in long QT syndrome[J].Scand J Clin Lab Invest, 2014,74(6):485-491.
[13] Van Slyke A C,Rezazadeh S,Snopkowski M,et al.Mutations within the S4-S5 linker alter voltage sensor constraints in hERG K+channels[J].Biophys J,2010,99(9):2841-2852.
[14] Latronico M V,Condorelli G.MicroRNAs and cardiac conduction [J].Curr Drug Targets,2010,11(8):907-912.
[15] Shan H,Zhang Y,Cai B,et al.Upregulation of microRNA-1 and microRNA-133 contributes to arsenic-induced cardiac electrical remodeling[J].Int J Cardiol,2013,167(6):2798-2805.
[16] Wang Q,Hu W,Lei M,et al.MiR-17-5p impairs trafficking of H-ERG K+channel protein by targeting multiple er stress-related chaperones during chronic oxidative stress[J].PLoS One, 2013,8(12):e84984.
[17] Lu X,Yang X,Huang X,et al.RNA interference targeting E637K mutation rescues hERG channel currents and restores its kinetic properties[J].Heart Rhythm,2013,10(1):128-136.
[18] Zarzoso M,Noujaim S F.Mission possible:RNA interference rescues the hERG current[J].Heart Rhythm,2013,10(1):137-138.
[19] Walker V E,Atanasiu R,Lam H,et al.Co-chaperone FKBP38 promotesHERGtrafficking[J].JBiolChem,2007,282(32):23509-23516.
[20] Matsa E,Dixon J E,Medway C,et al.Allele-specific RNA interference rescues the long-QT syndrome phenotype in humaninduced pluripotency stem cell cardiomyocytes[J].Eur Heart J, 2014,35(16):1078-1087.
[21] Wadhwa S,Wadhwa P,Dinda A K,et al.Differential expression of potassium ion channels in human renal cell carcinoma[J].Int Urol Nephrol,2009,41(2):251-257.
[22] Masi A,Becchetti A,Restano-Cassulini R,et al.hERG1 channels are overexpressed in glioblastoma multiforme and modulate VEGF secretion in glioblastoma cell lines[J].Br J Cancer,2005, 93(7):781-792.
[23] Pillozzi S,Brizzi M F,Balzi M,et al.HERG potassium channels are constitutively expressed in primary human acute myeloid leukemias and regulate cell proliferation of normal and leukemic hemopoietic progenitors[J].Leukemia,2002,16(9):1791-1798.
[24] Wang Y,Zhang Y,Yang L,et al.Arsenic trioxide induces the apoptosis of human breast cancer MCF-7 cells through activation of caspase-3 and inhibition of HERG channels[J].Exp Ther Med,2011,2(3):481-486.
[25] Pardo L A,Contreras-Jurado C,Zientkowska M,et al.Role of voltage-gated potassium channels in cancer[J].J Membr Biol, 2005,205(3):115-124.
[26] Calin G A,Dumitru C D,Shimizu M,et al.Frequent deletions and down-regulation of micro-RNA genes miR15 and miR16 at 13q14 in chronic lymphocytic leukemia[J].Proc Natl Acad Sci U S A,2002,99(24):15524-15529.
[27] Garzon R,Fabbri M,Cimmino A,et al.MicroRNA expression and function in cancer[J].Trends Mol Med,2006,12(12):580-587.
[28] Miska E A.How microRNAs control cell division,differentiation and death[J].Curr Opin Genet Dev,2005,15(5):563-568.
[29] Thomson J M,Newman M,Parker J S,et al.Extensive posttranscriptional regulation of microRNAs and its implications for cancer[J].Genes Dev,2006,20(16):2202-2207.
[30] Feng J,Yu J,Pan X,et al.HERG1 functions as an oncogene in pancreatic cancer and is downregulated by miR-96[J].Oncotarget,2014,5(14):5832-5844.
[31] Nowek K,Sun S M,Bullinger L,et al.Aberrant expression of miR-9/9*in myeloid progenitors inhibits neutrophil differentiation by post-transcriptional regulation of ERG[J].Leukemia,2016,30 (1):229-237.
[32] Kim J,Wu L,Zhao J C,et al.TMPRSS2-ERG gene fusions induce prostate tumorigenesis by modulating microRNA miR-200c[J].Oncogene,2014,33(44):5183-5192.
[33] Hart M,Wach S,Nolte E,et al.The proto-oncogene ERG is a target of microRNA miR-145 in prostate cancer[J].FEBS J, 2013,280(9):2105-2016.
[34] Burnett J C,Rossi J J,Tiemann K.Current progress of siRNA/shRNA therapeutics in clinical trials[J].Biotechnol J,2011,6 (9):1130-1146.
[35] Whitehead K A,Langer R,Anderson D G.Knocking down barriers:advances in siRNA delivery[J].Nat Rev Drug Discov,2009, 8(2):129-138.
[36] Shao X D,Wu K C,Guo X Z,et al.Expression and significance of HERG protein in gastric cancer[J].Cancer Biol Ther,2008,7(1): 45-50.
[37] Cherubini A,Taddei G L,Crociani O,et al.HERG potassium channels are more frequently expressed in human endometrial cancer as compared to non-cancerous endometrium[J].Br J Cancer,2000,83(12):1722-1729.
[38] Zhang R,Tian P,Chi Q,et al.Human ether-a-go-go-related gene expression is essential for cisplatin to induce apoptosis in human gastric cancer[J].Oncol Rep,2012,27(2):433-440.
[39] Staudacher I,Jehle J,Staudacher K,et al.HERG K+channel-dependent apoptosis and cell cycle arrest in human glioblastoma cells[J].PLoS One,2014,9(2):e88164.
[40] Glassmeier G,Hempel K,Wulfsen I,et al.Inhibition of HERG1 K+channel protein expression decreases cell proliferation of human small cell lung cancer cells[J].Pflugers Arch,2012,463(2): 365-376.
[41] Urbinati G,Ali H M,Rousseau Q,et al.Antineoplastic Effects of siRNA against TMPRSS2-ERG Junction Oncogene in Prostate Cancer[J].PLoS One,2015,10(5):e0125277.
[42] Zhao J,Wei X L,Jia Y S,et al.Silencing of herg gene by shRNA inhibits SH-SY5Y cell growth in vitro and in vivo[J].Eur J Pharmacol,2008,579(1-3):50-57.
[43] Asher V,Sowter H,Shaw R,et al.Eag and HERG potassium channels as novel therapeutic targets in cancer[J].World J Surg Oncol,2010,8:113.
2015-10-08)
(本文编辑:马雯娜)
国家自然科学基金(81370207);浙江省自然科学基金(LY13H020009)
315211 宁波大学医学院(庄凯丽);宁波市医疗中心李惠利医院心内科(廉姜芳)
猜你喜欢
保健医苑(2022年5期)2022-06-10 07:46:38
中国临床医学影像杂志(2021年6期)2021-08-14 02:21:56
肝博士(2020年5期)2021-01-18 02:50:18
华人时刊(2017年21期)2018-01-31 02:24:04
中国比较医学杂志(2018年5期)2018-01-22 13:21:04
海南医学(2016年8期)2016-06-08 05:43:00
健康管理(2016年2期)2016-05-30 21:36:03
中国医疗美容(2015年1期)2015-07-12 10:06:52
医学研究杂志(2015年9期)2015-07-01 17:27:46
医学研究杂志(2015年7期)2015-06-22 11:01:01
