
谷子MYB转录因子家族与抗旱关系的研究
2016-12-20 03:38程璐张彬张耀元苏彦冰魏东郭展李红英
山西农业大学学报(自然科学版) 2016年12期
程璐,张彬,张耀元,苏彦冰,魏东,郭展,李红英
(山西农业大学 农学院,山西 太谷 030801)
谷子MYB转录因子家族与抗旱关系的研究
程璐,张彬,张耀元,苏彦冰,魏东,郭展,李红英*
(山西农业大学 农学院,山西 太谷 030801)
[目的]MYB蛋白家族是一类含有MYB保守结构域的转录因子,广泛参与调控植物生长发育及抗逆等多种生理反应。为探究抗旱作物谷子的抗旱性与MYB转录因子家族基因关系,从而为谷子特有抗旱基因资源的发掘及谷子抗旱分子机制的研究提供参考。[方法]本研究以抗旱品种勾勾母鸡咀(GG)和干旱敏感品种晋汾16(JF)为研究对象,在苗期PEG处理条件下,取叶片进行转录组测序,进而对有差异表达的MYB转录因子家族基因进行生物信息学分析。[结果]研究表明,26个MYB转录因子基因在GG和JF两个谷子品种中表现出较为明显的表达差异;且对应启动子区域的抗旱相关调控元件也呈现出差异的组成与分布;抗旱品种GG中明显上调的3个MYB调控基因(Si034363m、Si003539m、Si030711m)与拟南芥中主要参与抗逆的MYB转录因子存在较近亲缘关系。[结论]基于对谷子MYB转录因子的研究,推测26个表达差异的MYB类转录因子基因可作为谷子抗旱的重要候选基因,可为后续谷子抗旱分子机制的研究提供理论依据。
谷子;MYB转录因子;干旱胁迫;生物信息学分析
谷子(Setariaitalica),在中国有7 000多年的栽培历史,是中国北方地区种植较广的小杂粮[1],主要分布于华北、西北和东北等干旱半干旱地区,具有极强的耐旱性。谷子虽然是耐旱作物,但干旱仍然是影响谷子产量的最大制约因素。谷子在萌发期如遇干旱种子萌发率会明显降低,植株延缓或停止生长;孕穗期遭遇干旱会出现秕谷,严重影响谷子产量[2]。因此,了解谷子的抗旱机制,对提高谷子产量尤为重要。
MYB类转录因子作为植物体内最大的转录因子家族之一,在植物的抗逆胁迫中发挥着重要的作用。已经有研究证实MYB 转录因子广泛参与植物的抗逆反应[3,4]。如厚叶旋蒴苣苔中的BcMYB1受干旱诱导表达,同时可以对PEG、高盐等胁迫产生应答[5];拟南芥的AT-MYB41可以在ABA、干旱胁迫下高水平表达,参与植物的抗旱反应[6]; AtMYB44和AtMYB60可以调控有关抗旱基因的表达,通过减小气孔开度达到减少水分散失的目的。AtMYB2在逆境下表达增强,增加下游干旱应答基因的表达[7]。水稻中的Osmyb4 基因的过量表达可以提高转基因植物对干旱、高盐等的耐受性[8]。
本研究以抗旱品种勾勾母鸡咀(GG)和干旱敏感品种晋汾16(JF)为材料,分析了对照和PEG胁迫下谷子MYB类转录因子的表达量差异,并且利用生物信息学手段比较了谷子与拟南芥中抗旱相关MYB基因的亲缘关系,旨在为谷子抗旱节水机制提供理论依据。
1 材料与方法
1.1 试验材料
抗旱品种勾勾母鸡咀(GG),干旱敏感品种晋汾16(JF),均由山西农业大学农业生物工程研究所提供。
1.2 试验方法
1.2.1 谷子种植及表达谱测序
供试材料在人工气候室中采用营养土∶蛭石= 3∶1种植,在14 h光照28 ℃/10 h黑暗23 ℃的条件下培养,培养21 d后,釆用20% PEG-6000模拟干旱,处理材料0.5 h,对照用蒸馏水处理相同时间,对照和处理各取3株,立即放入液氮速冻,提取RNA,进行转录组测序。
1.2.2 谷子MYB基因家族抗旱相关基因的生物信息学分析
利用R语言pheatmap程序绘制基因表达热图。利用Phytozome 11.0谷子基因组数据库查找MYB基因序列信息。利用PlantCARE分析启动子元件。利用NCBI查找在拟南芥MYB基因家族中的抗旱相关基因。利用MEGA6.0软件构建MYB抗旱基因家族系统发育树。
2 结果与分析
2.1 谷子MYB抗旱相关基因表达量分析
根据转录组数据,选取了干旱胁迫后GG和JF中表达有差异的基因,绘制了2个品种在干旱和非干旱条件下的表达热图。在聚类图中,用颜色的深浅表示基因表达量的变化。紫色代表基因表达量较高,绿色代表基因表达水平比较低,颜色越深,代表基因表达量越大。图1中从左到右依次代表GG、JF在非干旱条件下和干旱条件下的表达量变化情况。由图1可见,2个谷子品种GG和JF在干旱胁迫处理后26个基因与对照相比都有不同程度的变化,有部分基因表达变化较明显,如Si009576m、Si002426m、Si034363m。根据热图聚类分析,可以将这些基因分为两大类,Si014513m、Si009576m、Si002426m、Si034363m、Si006824m为一大类,其余基因为一大类。第一大类在两个品种中的表达差异较大,例如Si014513m、Si009576m、Si002426m、Si034363m在干旱胁迫处理之前GG的表达量明显高于JF,干旱胁迫后Si014513m、Si002426m两个基因在GG中的表达量高于JF。但是,Si009576m在干旱处理之后表达量较对照组高;Si034363m在两个品种中的表达差异不明显。

图1 GG和JF中MYB转录因子PEG处理前后表达谱分析Fig.1 The expression profiling analysis of MYB transcription factors before and after PEG treatment in GG and JF
第二大类又分为两个亚类,第一亚类在两个品种中干旱和非干旱条件下的表达差异不明显,而且表达水平比较低,第二亚类基因的表达量比第一大类基因表达量低,比第一亚类基因表达水平高。Si026734m、Si003538m、Si030441m表达水平相似,即在干旱处理之前,在JF中表达量较高,在干旱处理之后,在GG中表达量较高。从整体来看,各个基因在GG和JF的不同处理之间表达关系复杂。
2.2 谷子MYB抗旱相关基因基本信息分析
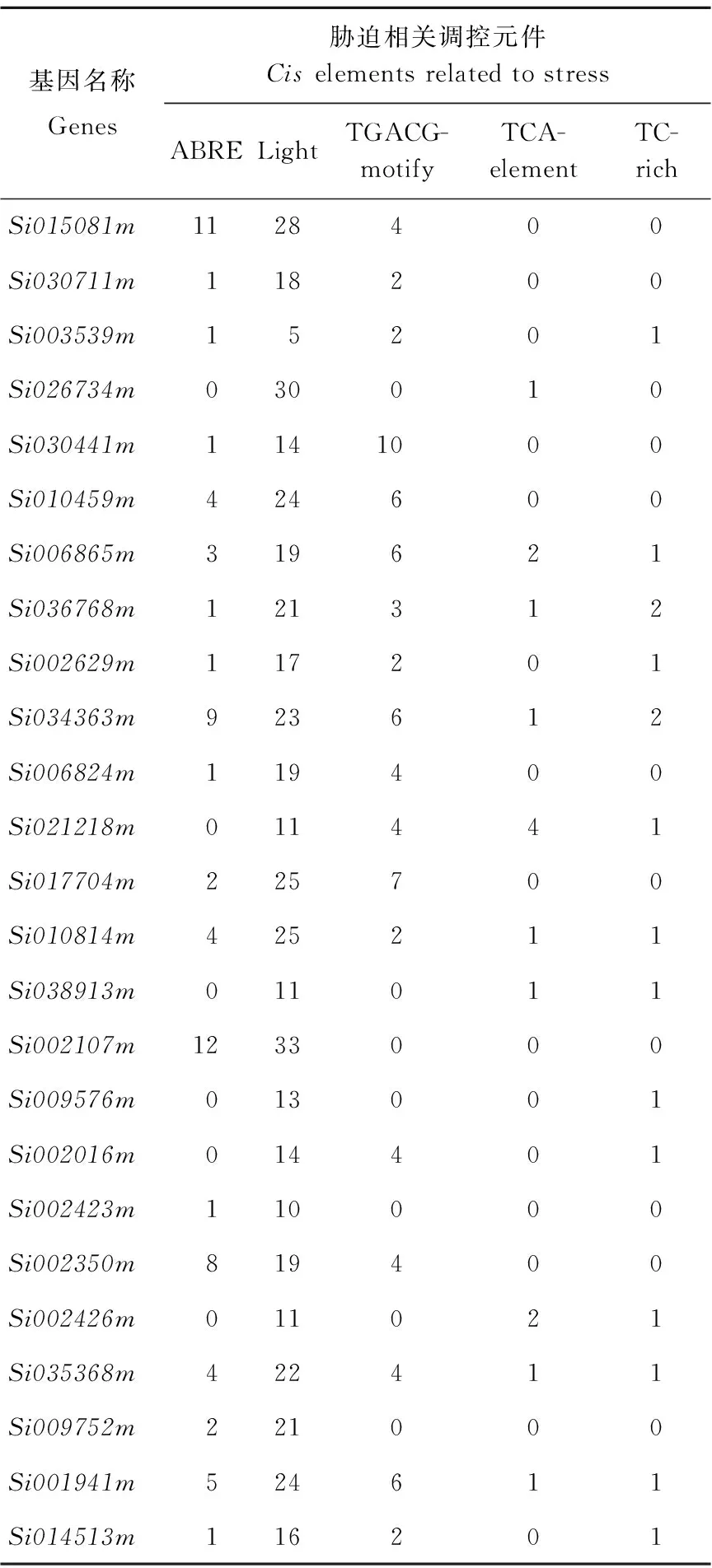
在谷子基因组数据库Phytozome中查找谷子MYB基因家族与抗旱有关基因的相关信息(基因组序列的长度,氨基酸序列长度,染色体位置,以及内含子和外显子数目),结果见表1。以品种豫谷1号为参照,在Phytozome中查找基因上游1 500 bp启动子序列(表2),从表2可以看出这些基因中与光响应有关的元件最多,Si026734m与此功能相关的元件高达30个。这些基因至少含有一个抗旱相关元件。例如Si010459m不含有TCA,TC-rich,MBS调控元件,但是它还含有TFACG-motify元件 6个,ABRE元件4个。Si002107m所含的ABRE元件最多,高达12个。茉莉酸甲酯作为与损伤相关的植物激素和信号分子,广泛地存在于植物体中。谷子MYB基因家族与抗旱相关的基因中,含有0~10个TFACG-motify调控元件,其中Si030441m基因含量最多(10个)。此外,这些基因还含有TCA、TC-rich和MBS调控元件,它们均与抗旱相关。
2.3 GG中表达上调基因与拟南芥MYB转录因子抗旱相关基因系统发育树分析
通过GG和JF抗旱基因表达谱可以看出,抗旱品种GG中的Si034363m、Si003539m、Si030441m、Si030711m、Si026734m在干旱胁迫处理后基因表达量都有不同程度的增长。利用MEGA6.0对这5个基因与拟南芥中和抗旱相关的21个基因进行进化树分析(图2)。从图2可以看出,Si030711m与AtMYB102、AtMYB41亲缘关系比较近,Si003539m与AtMYB104,Si034363m与AtMYB44亲缘关系最近。
表1 谷子MYB转录因子基因特征
Table 1 The characteristics of MYB transcription factor in foxtail millet

基因名称Genesname基因长度/bpGeneslength氨基酸个数Aminoacidsnumber染色体位置Chromosomelocation外显子数目ExonsnumberSi034207m7938860916Si015081m75024961Si030711m142927322Si003539m14578852Si026734m155728082Si030441m477635352Si010459m364035574Si006865m288333246Si036768m80726891Si002629m165125653Si034363m5479761914Si006824m562034249Si021218m8311968312Si017704m196834313Si010814m284127473Si038913m293954394Si002107m450834053Si009576m455565472Si002016m272423952Si002423m186929452Si002350m173830852Si002426m270029452Si035368m285749493Si009752m6871554710Si001941m453136653Si014513m979426265
表2 谷子MYB转录因子基因启动子元件分析
Table 2 The promoter element analysis of MYB transcription factors in foxtail millet
基因名称Genes胁迫相关调控元件CiselementsrelatedtostressABRELightTGACG-motifyTCA-elementTC-richMBSSi015081m11284001Si030711m1182003Si003539m152012Si026734m0300103Si030441m11410002Si010459m4246000Si006865m3196210Si036768m1213121Si002629m1172011Si034363m9236121Si006824m1194003Si021218m0114411Si017704m2257003Si010814m4252111Si038913m0110112Si002107m12330001Si009576m0130016Si002016m0144015Si002423m1100002Si002350m8194001Si002426m0110212Si035368m4224116Si009752m2210004Si001941m5246111Si014513m1162011
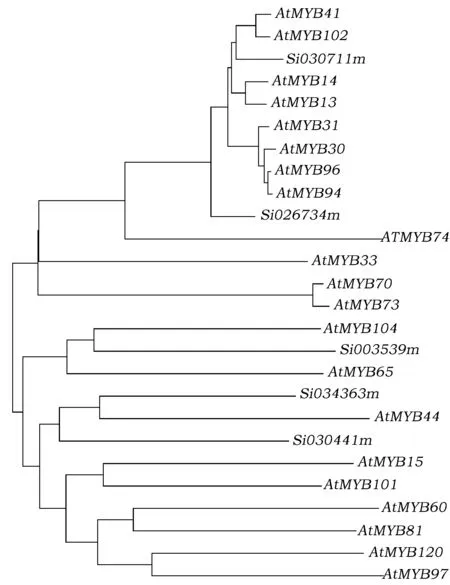
图2 GG中表达上调基因与拟南芥MYB转录因子抗旱相关基因系统发育树分析Fig.2 Phylogenetic tree analysis with five up-regulated genes in GG and some MYB genes in Arabidopsis which is related with drought tolerance
3 结论与讨论
干旱影响作物生长和产量。谷子作为耐旱性极强的作物,研究其抗旱机理对于小麦、水稻等作物的抗旱改良具有极其重要的意义。本文通过对谷子中MYB转录因子进行转录组数据分析,探讨这些转录因子与抗旱的关系。从MYB转录因子在干旱胁迫处理之后的表达量变化情况来看,这些转录因子调节干旱胁迫的机理比较复杂,而且在不同的品种中存在差异。
Si002107m含有的ABRE元件最多,ABRE是脱落酸合成相关元件。抗旱品种GG在干旱胁迫后Si002107m表达量增大,可能是因为该基因在接收到干旱胁迫信号后,ABRE元件被激活,促进脱落酸合成,使谷子适应干旱环境。但是干旱敏感品种JF在处理和对照之间表达量差异不明显,说明这个转录因子在调节干旱胁迫时存在品种差异。经过比对,发现该转录因子与玉米转录因子MYB-IF35同源性最高,而MYB-IF35在调节植物次级代谢、抵抗各种压力胁迫中起重要作用[9],本研究结果与前人研究一致。
拟南芥MYB44在遭受逆境胁迫(如病害,盐胁迫,冷害)后,表达量提高,植物抗逆性增强[10]。从系统进化树结果来看,Si034363m与MYB44亲缘关系最近,而且这个基因干旱胁迫后在GG和JF中表达都增强了,与以往研究结果[10]一致。
MYB转录因子与抗旱相关基因的转录调控在干旱胁迫的反应中起着承上启下的作用。但是这些基因启动干旱信号通路并参与抗旱反应的机制目前还不清楚,尚需进一步深入研究。
鉴定和阐明MYB类转录因子的功能以及其应对逆境胁迫的调控网络对植物的分子抗性育种有着重要意义。通过改良或者增加一个MYB关键转录因子的调控能力,可以激活下游多个功能基因的表达,进而提高植物的抗旱能力,这在植物的分子抗旱育种中有着广阔的应用前景。
[1]Doust AN, Kellogg EA, Devos KM, et al. Foxtail millet: a sequence-driven grass model system [J]. Plant Phy, 2009, 149(1):137-141.
[2]张雁明,刘晓东,马建萍,等.谷子抗旱研究进展[J].山西农业科学,2013,41(3):282-285.
[3]刘蕾,杜海,唐晓凤.MYB转录因子在植物抗逆胁迫中的作用及其分子机理[J].遗传,2008,30(10): 1265-71.
[4]乔孟,于延冲,向凤宁.拟南芥R2R3-MYB类转录因子在环境胁迫中的作用[J]. 生命科学, 2009, 21(001): 145-50.
[5]Chen BJ, Wang Y, Hu YL, et al. Cloning and characterization of a drought-inducible MYB gene from Boea crassifolia [J]. Plant Sci, 2005, 168(2): 493-500.
[6]Cominelli E, Sala T, Calvi D, et al. Overexpressions of the Arabidopsis AT-MYB41 gene alter cell expanision and leaf surface permeability [J]. Plant J, 2008, 53(1): 53-64.
[7]Hoeren FU, Dolferus R, Wu Y, et al. Evidence for a role for AtMYB2 in the induction of the Arabidopsis alcohol dehydrogenase gene (ADH1) by low oxygen [J]. Genetics, 1998, 149(2): 479?490.
[8]Valliyodan B, Nguyen HT. Understanding regulatory networks and engineering for enhanced drought tolerance in plant [J]. Curr Opin Plant Biol, 2006, 9(2): 189-95.
[9]Heine GF, Malik V, Dias AP,et al. Expression and molecular characterization of ZmMYB-IF35 and related R2R3-MYB transcription factors [J]. Mol Biotech, 2007, 37(2): 155-164.
[10]时浩杰.转录因子AtMYB44和AtMYB15在拟南芥防卫反应中的功能研究[D],南京农业大学,2010.
(编辑:马荣博)
MYB transcription factors and their relationship with drought tolerance in foxtail millet
Cheng Lu, Zhang Bin, Zhang Yaoyuan, Su Yanbing, Wei Dong, Guo Zhan, Li Hongying*
(CollegeofAgriculture,ShanxiAgriculturalUniversity,Taigu030801,China)
[Objective]MYB transcription factors are a class of protein with conserved MYB domain. They are involved in the regulation of plant growth, stress resistance and other physiological responses. Foxtail millet is an important drought-tolerance crop in northern China. It is important to explore the unique genetic resources and the molecular mechanism of drought-tolerance in foxtail millet to understand the relationship between drought resistance and MYB transcription factor family genes. [Methods]In this study, the drought-tolerant variety Go Go Mu Ji Ju (GG) and the drought sensitive variety Jin Fen 16 (JF) were used as experimental materials, they were treated by PEG6000 in seedling stage, and transcriptome sequencing was conducted with mRNA from GG and JF leaves, bioinformatics analysis were used to analyze the differentially expressed MYB transcription factors. [Results]The results showed that: there were 26 MYB transcription factor genes that showed apparent differential expression in GG and JF, in addition, the drought-tolerant regulatory elements in promoters of the corresponding genes showed different composition and distribution.Si034363m,Si003539mandSi030711m, which were up-regulated upon drought stress in GG, showed close homology with some MYB transcription factors that mainly involved in drought tolerance inArabidopsis. [Conclusion]Based on the research of MYB transcription factors in foxtail millet, we speculated that 26 differentially expressed MYB transcription factor genes could be used as important drought tolerant candidate genes in foxtail millet. This study could provide a theoretical basis for the study of the molecular mechanism of drought tolerance in foxtail millet.
Setariaitalica, MYB transcription factors, Drought stress, Bioinformatics analyses
2016-08-28
2016-10-20
程璐(1992-),女(汉),山西吕梁人,硕士,研究方向:植物基因工程
*通讯作者:李红英,教授,博士生导师。Tel:0354-6287239; E-mail:hongying1964@163.com
国家自然科学基金(31471556)
S515
A
1671-8151(2016)12-0846-04
猜你喜欢
当代陕西(2021年21期)2022-01-19
今日农业(2021年14期)2021-11-25
今日农业(2020年13期)2020-12-15
国际放射医学核医学杂志(2020年4期)2020-07-27
建材发展导向(2019年11期)2019-08-24
生命科学研究(2018年1期)2018-05-29
河南水利年鉴(2017年0期)2017-05-19
上海农业学报(2017年3期)2017-04-10
现代农业(2015年5期)2015-02-28
植物营养与肥料学报(2014年1期)2014-03-11
