
非洲菊单倍体植株的生长特性初报
2016-12-07 09:58单芹丽阮继伟吴丽芳汪国鲜李绅崇杨春梅
江西农业学报 2016年12期
单芹丽,阮继伟,吴丽芳,张 松,汪国鲜,李绅崇*,杨春梅*
(1.云南省农业科学院 花卉研究所/国家观赏园艺工程技术研究中心,云南 昆明 650205;2.玉溪云星生物科技有限公司,云南 玉溪 652604)
非洲菊单倍体植株的生长特性初报
单芹丽1,2,阮继伟1,吴丽芳2,张 松2,汪国鲜1,李绅崇1*,杨春梅2*
(1.云南省农业科学院 花卉研究所/国家观赏园艺工程技术研究中心,云南 昆明 650205;2.玉溪云星生物科技有限公司,云南 玉溪 652604)
以4个非洲菊品种胚珠诱导的单倍体植株为材料,相应母本杂合二倍体植株为对照,研究了非洲菊单倍体植株的生长繁殖特性。结果表明:单倍体的繁殖系数、开花植株最大叶片长、花序直径、炼苗至开花所需时间在不同基因型间存在显著差异;非洲菊繁殖系数大小在苗期和移栽期均与倍性大小无关,与杂合二倍体相比,单倍体株型较小;鲜重增加速度相对较慢;炼苗至开花所需天数相对较长,单倍体需261.3~354.8 d,而杂合二倍体只需要117.0~132.5 d。
非洲菊;二倍体;单倍体;生长特性
非洲菊(GerberajamesoniiBolus)为菊科大丁草属多年生宿根草本植物,是世界著名的五大鲜切花之一,属于典型的异花授粉植物,基因型高度杂合[1],致使后代纯合体选育极度困难。单倍体植株经染色体加倍后,可快速成为纯合二倍体,进而提高育种效率[2]。国外通过非洲菊胚珠培养获得单倍体植株的研究相对较早[3-4]。国内自2007年起,云南省农业科学院花卉研究所首先开展了相关研究[5-7],通过多年的研究积累,获得并保存了一批非洲菊单倍体株系,但单倍体植株的长势较差,制约了对其进一步的利用和深入研究。因此,有必要对非洲菊单倍体植株的生长特性开展相关研究。
目前大部分非洲菊的商业品种几乎都为杂合二倍体[8],其生长特性已有相关研究[9-11],其中用花托进行组培繁殖的报道相对较多[12-13],但非洲菊单倍体植株的生长繁殖特性方面的研究还未见相关的系统报道。为此,本试验在前期研究的基础上,以母本杂合二倍体植株为对照,研究不同基因型非洲菊单倍体植株的鲜重增加速度、繁殖系数、株高、炼苗至开花所需时间、最大叶片的长和宽、花序直径和花梗长,以便对单倍体植株的生长特性有较清晰的了解,为推进非洲菊单倍体植株在育种中的研究利用奠定基础。
1 材料与方法
1.1 材料
材料来自云南省农科院花卉研究所的非洲菊育种资源圃,为同一时间采摘的4个非洲菊品种白马王子、阳光露、鸿运当头和红极星的花蕾和胚珠诱导培养的杂合二倍体和单倍体株系,继代次数均为4次。
1.2 试验设计
试验以不同杂合二倍体非洲菊品种(花托诱导培养的再生植株)为对照,将相应不同品种来源诱导的单倍体植株(胚珠诱导培养的再生植株)设为不同处理(表1),每个处理设4次重复。将组培苗的丛生芽切成单株,株高保留1 cm长左右,接种在MS+BA 0.6 mg/L+NAA 0.1 mg/L、附加食用白糖30 g/L、琼脂7 g/L、pH值5.5~5.8的非洲菊常规增殖培养基中,置于温度(22±2) ℃、光照强度3200~4000 lx、光照时间为10 h/d的培养室培养。接种前称量各瓶的苗鲜重,每个处理接种10瓶,每瓶接种8株。瓶苗培养30 d后,每个处理中随机抽取未污染的4瓶(4次重复),称量每瓶苗的鲜重,并调查苗期各重复的株高、最大叶片的长和宽、单株新株萌发后高0.5 cm以上的植株数量。随后将各处理调查后剩余的瓶苗切成单株,生根炼苗后移栽花盆内,每个处理盆栽15株,置于大棚离地苗床内,按完全随机排列,株行距30 cm×35 cm,移栽苗要求健康、叶片4~5个、新根生长数5条以上,栽培基质为红土∶腐殖土=2∶1,移栽前称量各重复植株的鲜重,采用常规方法进行肥水管理直至开花,期间观察并记录各植株第一朵花的开花时间,栽培365 d时,各处理随机抽取10株,调查栽培期各重复炼苗至开花所需时间、称量鲜重、测量株高、花序直径、花梗长、最大叶片的长和宽,单株新株萌发后高0.5 cm以上的植株数量。

表1 试验处理及对照设计
1.3 统计与分析
统计各处理组培苗和栽培植株的繁殖系数、株高、最大叶片的长和宽、鲜重增加速度、炼苗至开花所需时间、花序直径和花梗长,计算各测量参数的平均值,具体计算方法如下。
繁殖系数=单株新株萌发后高0.5 cm以上的植株数量/接种或移栽前的株数。
鲜重增加速度=每株苗的鲜重增加量(调查时每株苗的鲜重-接种或炼苗前每株苗的鲜重)/室内培养或室外栽培的天数。
试验数据采用Excel 2003和DPS软件进行统计分析,利用Duncan’s新复极差法对各处理进行差异显著性检验。
2 结果与分析
2.1 单倍体和杂合二倍体组培苗及栽培植株的繁殖系数比较
不同非洲菊品种诱导的单倍体植株繁殖系数在苗期和移栽期与对照杂合二倍体相比发生了一些变化,不同品种的变化情况见图1。白马王子诱导的单倍体组培苗继代培养的繁殖系数与杂合二倍体相比没有变化,均为3.4,栽培植株的繁殖系数则小于杂合二倍体的繁殖系数;鸿运当头诱导的单倍体植株组培苗的繁殖系数与杂合二倍体相比也没有变化,均为2.9,但栽培植株的繁殖系数则大于杂合二倍体的繁殖系数;阳光露诱导的单倍体组培苗和栽培植株的繁殖系数都小于杂合二倍体的繁殖系数;而红极星诱导的单倍体组培苗和栽培植株的繁殖系数都大于杂合二倍体的繁殖系数。结果表明:非洲菊的繁殖系数大小在苗期和移栽期均与倍性大小无相关性,并且单倍体组培苗和栽培植株的繁殖系数在不同基因型间均存在显著差异。
2.2 单倍体和杂合二倍体组培苗及栽培植株的鲜重增加速度比较
由图2可知,单倍体栽培植株比组培苗的鲜重增加速度快,并且与对照杂合二倍体相比,单倍体植株鲜重增加速度相对较慢。单倍体栽培植株增加量为76.6~423.8 mg/(株·d),而组培苗的增加量只有6.3~9.2 mg/(株·d),其中阳光露诱导的单倍体栽培植株和二倍体栽培植株相比差异明显,二倍体的增加量是单倍体的5倍多,单倍体只有76.6 mg/(株·d),而二倍体高达438.1 mg/(株·d)。
2.3 单倍体与杂合二倍体植株的株型对比
非洲菊单倍体和对照杂合二倍体植株相比,株型较矮小,叶片短窄,花朵较小。A、B、C、D的左边均为杂合二倍体,右边均为单倍体,A:红极星诱导培养的杂合二倍体和单倍体组培增殖苗;B:阳光露诱导培养的杂合二倍体和单倍体组培苗叶片;C:红极星诱导培养的杂合二倍体和单倍体开花植株;D:阳光露诱导培养的杂合二倍体和单倍体开花植株的叶片和花朵(图3)。
2.3.1 株高差异 从图4可知,同一培养和栽培环境条件下,在非洲菊的整个生长发育阶段,单倍体的株高均低于杂合二倍体的株高。组培苗增殖培养基上培养30 d后,单倍体株高为1.7~2.3 cm,杂合二倍体株高为2.8~4.4 cm;移栽苗在室外栽培365 d后,单倍体的株高为14.1~19.6 cm,杂合二倍体株高为25.1~36.1 cm。
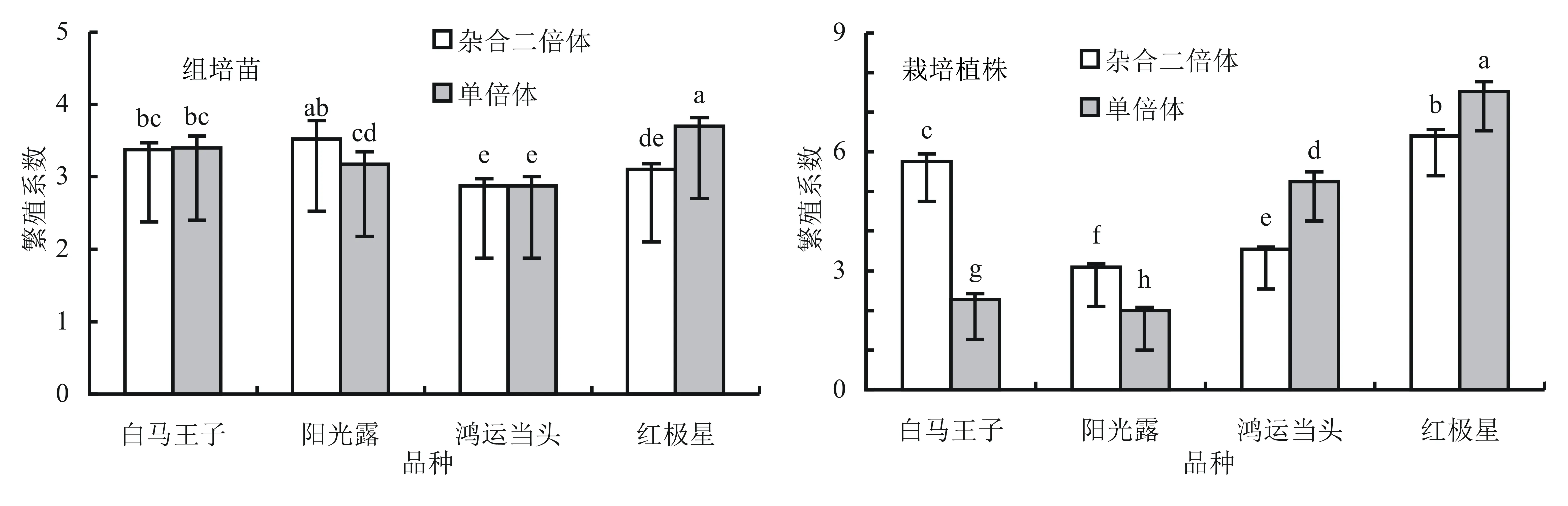
图1 非洲菊单倍体和杂合二倍体组培苗及栽培植株的繁殖系数

图2 非洲菊单倍体和杂合二倍体组培苗及栽培植株的鲜重增加速度

图3 单倍体与杂合二倍体代表株的株型对比图

图4 非洲菊单倍体和杂合二倍体组培苗及开花植株的株高比较
2.3.2 叶片长宽对比 从表2可知,单倍体与对照杂合二倍体相比,叶片长和叶片宽都较小,并且各处理开花植株的最大叶片长在0.05水平上差异显著,单倍体组培苗培养30 d后,单倍体的最大叶片长、叶宽与杂合二倍体相比差异不明显,但移栽苗栽培365 d后,单倍体的最大叶片长、叶宽与杂合二倍体相比差异较大,单倍体开花植株的叶片长在10.0~12.9 cm之间,叶片宽度在1.9~4.5 cm之间,杂合二倍体叶片长在20.1~22.4 cm之间,叶片宽在6.8~11.1 cm之间。
2.3.3 花朵大小对比 由图5可知,单倍体的花梗长及花序直径都较对照杂合二倍体的短小,其中处理1和处理2、处理3和处理4的单倍体花梗长差异不显著,但不同品种来源的单倍体花序直径在不同基因型间存在显著差异,并且变化幅度较大,在2.1~7.0 cm之间,而杂合二倍体的花序直径在不同基因型间的变化幅度较小,在9.2~10.1 cm之间。
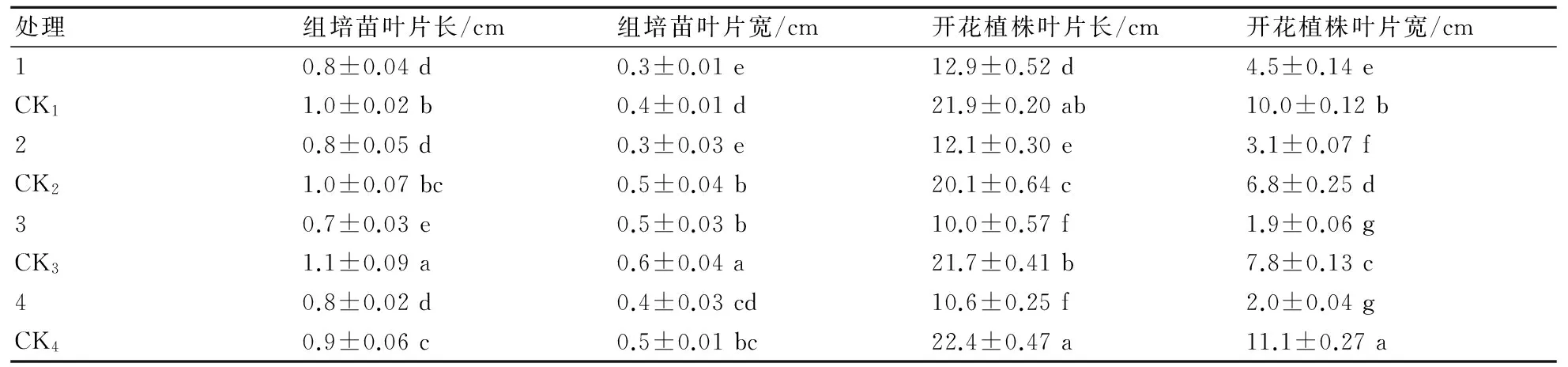
表2 非洲菊单倍体和杂合二倍体组培苗及开花植株长宽对比
注:同列数值后小写字母表示在0.05水平上的差异显著性。

图5 单倍体与杂合二倍体植株的花朵大小
2.4 单倍体和杂合二倍体从炼苗至开花所需天数对比
杂合二倍体和单倍体植株从炼苗至开花所需天数的观察记录结果见图6。单倍体从炼苗至开花所需天数在不同基因型间存在显著差异,不同处理间相差达93.5 d,而对照杂合二倍体在4个品种间相差不到16 d,并且与杂合二倍体相比,单倍体植株从炼苗至开花所需天数较长,是杂合二倍体植株的2倍多,杂合二倍体需要117.0~132.5 d,而单倍体长达261.3~354.8 d。

图6 单倍体和杂合二倍体从炼苗至开花所需天数
3 讨论
谭志勇等研究了7个甘蔗基因型的组织培养特性,认为不同甘蔗基因型之间的生长速度有明显的差异[14]。本试验的研究结果也表明:4个非洲菊品种诱导的单倍体栽培植株的鲜重增加速度、炼苗至开花所需时间在不同基因型间均差异显著,证明非洲菊单倍体的生长速度在不同基因型间存在差异。
吴红英等研究表明:不同品种红掌组织培养的增殖倍率不同[15],张永兵等以单倍体甜瓜无菌苗的顶芽和腋芽为外植体进行离体繁殖研究,认为与二倍体甜瓜相比,单倍体甜瓜的增殖能力较弱[16]。在本试验中,单倍体组培苗和栽培植株的繁殖系数在不同基因型间均存在显著差异,并且发现非洲菊的繁殖系数大小在苗期和移栽期均与倍性大小无相关性,如:来源于白马王子和鸿运当头诱导培养的单倍体和杂合二倍体在组培苗的繁殖系数差异不显著,但由白马王子诱导培养的杂合二倍体栽培植株繁殖系数高于单倍体栽培植株的繁殖系数,而由鸿运当头诱导的杂合二倍体栽培植株繁殖系数则低于单倍体栽培植株。
许多研究表明:植株的形态特征在不同倍性间的存在差异[16]。玉米穗轴长、株高、穗位高和雄穗主轴长从单倍体水平到二倍体呈增大的趋势,而雄穗分支数呈减小的趋势[18],灯盏花四倍体与二倍体相比,植株具有巨大性[19],苎麻随着倍性的增加,茎的平均粗度呈增大的趋势[20],西瓜叶面积同源四倍体与二倍体的相比显著增大[21]。本研究结果表明:非洲菊单倍体和对照杂合二倍体植株相比,株型较矮小,叶片短窄,花朵较小。
4 结论
与对照杂合二倍体相比,非洲菊单倍体株型较小,表现为株高较矮、叶片较短窄、花枝较短、花序直径较小;单倍体的鲜重增加速度相对较慢;炼苗至开花所需天数相对较长,单倍体需要261.3~354.8 d,而杂合二倍体只需要117.0~132.5 d。非洲菊繁殖系数大小在苗期和移栽期均与倍性大小无关,无论是单倍体还是杂合二倍体,非洲菊的繁殖系数都与基因型相关,同时,非洲菊单倍体的开花植株最大叶片长、花序直径、炼苗至开花所需时间在不同基因型间也存在显著差异。
[1] 李绅崇,李淑斌,蒋亚莲,等.非洲菊品种间杂交主要观赏性状在F1代的遗传表现[J].云南农业大学学报,2007,22(2):197-201.
[2] 张献龙,唐克轩.植物生物技术[M].北京:科学出版社,2004:174-177.
[3] Miyoshi K, Asakura N. Callus induction, regeneration of haploid plants and chromosome doubling in ovule cultures of pot gerbera (Gerberajamesonii)[J]. Plant Cell Reports, 1996, 16(1): 1-5.
[4] Tosca A, Arcara L, Frangi P. Effect of genotype and season on gynogenesis efficiency in Gerbera[J]. Plant Cell, Tissue and Organ Culture, 1999, 59(1): 77-80.
[5] 单芹丽,王继华,吴丽芳,等.不同供体材料对非洲菊胚珠离体培养的影响[J].植物生理学报,2015,51(7):1151-1156.
[6] 单芹丽,李绅崇,王继华,等.基因型和预处理对非洲菊胚珠离体培养的影响[J].江西农业学报,2015,27(7):44-47.
[7] 王丽花,瞿素萍,杨秀梅,等.非洲菊未授粉胚珠的离体诱导和植株再生[J].植物生理学通讯,2007,43(6):1089-1092.
[8] 李涵,鄢波,张婷,等.切花非洲菊多倍体诱变初报[J].园艺学报,2009,36(4):605-610.
[9] 李绅崇,蒋亚莲,吴丽芳,等.非洲菊良种保持及繁育技术[J].北方园艺,2008(7):192-193.
[10] 牛佳,程智慧.现蕾期营养液浓度对基质栽培非洲菊生长和养分吸收的影响[J].西北农林科技大学学报,2013,41(1):130-136.
[11] 王宝钦.无土栽培基质特性与非洲菊生长关系的研究[J].科技园地,2015(10):8-9.
[12] 高艳明,李建设,李晓娟.非洲菊花托组织培养的研究[J].西北农业学报,2006,15(4):200-202.
[13] 李高燕,王海云,牛佳佳,等.非洲菊组织培养研究进展[J].中国农学通报,2009,25(10):72-76.
[14] 谭志勇,谭中文,张志胜.不同基因型甘蔗组织培养特性的研究[J].广东农业科学,2005(1):34-36.
[15] 吴红英,蔡林,何贵整,等.不同红掌品种在组培生产上的差异表现[J].北方园艺,2012(16):86-87.
[16] 胡婷婷,王健康,丁成伟,等.粳稻品种间杂交F1代花药培养及后代鉴定评价[J].南方农业学报,2014,45(11):1910-1915.
[17] 张永兵,伊鸿平,贾媛媛,等.单倍体甜瓜离体繁殖的方法[J].园艺学报,2007,34(2):497-500.
[18] 张如养,段民孝,赵久然,等.玉米单倍体与其加倍后代性状的相关性研究[J].种子,2014,33(11):72-74.
[19] 梅琳,郑思乡,杨声超,等.不同倍性灯盏花形态学与细胞学研究[J].贵州农业科学,2009,37(7):134-135.
[20] 晏春耕,曹瑞芳,李宗道,等.不同倍性苎麻形态与产量、品质性状的研究[J].中国农业科学,2003,36(6):628-632.
[21] 王镇,党选民,詹园凤.不同倍性小型西瓜主要农艺性状的比较分析[J].中国蔬菜,2010(2):51-55.
(责任编辑:曾小军)
Preliminary Study on Growth Characteristics of Haploid Plants ofGerberajamesonii
SHAN Qin-li1,2,RUAN Ji-wei1, WU Li-fang2, ZHANG Song2,WANG Guo-xian1, LI Shen-chong1*, YANG Chun-mei2*
(1. Flower Research Institute, Yunnan Academy of Agricultural Sciences / National Research Center for Ornamental Horticulture Engineering and Technology, Kunming 650205, China; 2. Yuxi Yunxing Biotechnology Company Limited in Yunnan Province, Yuxi 652604, China)
The haploid plants induced from the ovules of fourGerberajamesoniicultivars were used as the experimental materials, their female-parent heterozygous diploid plants were selected as the control materials, and the growth and reproduction characteristics of haploid plants ofGerberajamesoniiwere studied. The results showed that the propagation coefficient, the maximum blade length of flowering plants, the inflorescence diameter, and the duration from hardening-seedling to flowering were significantly different among various genotypes of haploids. The propagation coefficient ofGerberajamesoniiat both seedling stage and transplanting stage was nothing to do with its ploidy. In comparison with the heterozygous diploid plants, the haploid plants had relatively small plant type, relatively slow fresh-weight gain speed, and relatively long duration from hardening-seedling to flowering (261.3~354.8 d for haploid plants, whereas 117.0~132.5 d for heterozygous diploid plants).
Gerberajamesonii; Diploid; Haploid; Growth characteristics
2016-09-05
云南省科技厅技术创新暨产业发展专项资金项目(2015XB015);国家科技支撑计划项目(2015BAD10B00);云南省科技厅科技创新强省计划项目(2014AB014)。
单芹丽(1973─),女,云南祥云人,研究员,硕士,主要从事花卉育种与组培繁殖方面的研究。*通讯作者:李绅崇、杨春梅。
S682.11
A
1001-8581(2016)12-0013-05
猜你喜欢
现代农业科技(2022年1期)2022-01-17
上海农业科技(2020年4期)2020-08-19
山西农业科学(2020年8期)2020-08-13
河南农业科学(2018年5期)2018-01-19
江苏农业科学(2017年16期)2017-10-27
现代农业科技(2017年8期)2017-06-10
浙江农业科学(2016年11期)2016-05-04
中国学术期刊文摘(2015年8期)2015-10-29
西北农林科技大学学报(自然科学版)(2015年5期)2015-02-21
桉树科技(2015年1期)2015-02-14
