
柑橘绿霉病菌木聚糖酶基因(PdXY 2)功能初步研究
2016-12-06 03:01由书妍刘红霞于瑞君郑丽君范诗言李红叶
植物保护 2016年6期
由书妍, 刘红霞, 于瑞君, 郑丽君, 范诗言, 李红叶
(1.大连市农业科学研究院, 大连 116036; 2. 浙江大学生物技术研究所, 杭州 310058)
柑橘绿霉病菌木聚糖酶基因(PdXY2)功能初步研究
由书妍1, 刘红霞1, 于瑞君1, 郑丽君1, 范诗言1, 李红叶2*
(1.大连市农业科学研究院, 大连 116036; 2. 浙江大学生物技术研究所, 杭州 310058)
柑橘绿霉病菌(Penicilliumdigitatum)是引起柑橘储藏期腐烂最主要的病原之一,严重影响了柑橘产业发展。本试验克隆了柑橘绿霉病菌的木聚糖酶基因(PdXY2),对其表达进行研究,在柑橘发病过程中,PdXY2的表达显著升高,至48 h达到最高,其表达量为对照的4倍。为进一步明确PdXY2基因功能,构建了PdXY2基因缺失突变株(ΔPdXY2),ΔPdXY2突变株的致病性与野生型相比没有明显差异。由此我们推断,在柑橘绿霉病菌侵染柑橘的过程中PdXY2有一定的作用,但是单一缺失木聚糖酶PdXY2基因不能影响其致病性。
柑橘绿霉病菌; 木聚糖酶; 基因表达; 基因突变
植物病原菌侵入寄主时必须先破坏寄主的细胞壁,这个过程主要有两方面作用:一方面破坏寄主的天然屏障,使病原物能侵入寄主,建立侵染关系;另一方面降解的产物能够为病原物提供营养物质。在细胞壁的降解过程中,细胞壁水解酶起到了重要的作用[1-2]。已有研究表明细胞壁水解酶对致病性至关重要,如在黄曲霉(Aspergillusflavus)中,内切多聚半乳糖醛酸酶突变株对棉铃的致病性降低[3];在灰葡萄孢(Botrytiscinerea)中,内切多聚半乳糖醛酸酶突变株对番茄的致病性降低[4],果胶甲酯酶突变株对多种作物的致病性均有所下降[5];在稻瘟病菌(Magnaportheoryzae)中,角质酶突变株对水稻和大麦的致病性下降[6];在胶孢炭疽菌(Colletotrichumgloeosporioides)中,果胶裂解酶突变株对牛油果的致病性下降[7-8];紫麦角菌(Clavicepspurpurea)中,多聚半乳糖醛酸酶基因突变株对黑麦的致病性下降[9]。因此,研究细胞壁水解酶对探索病原菌的致病性有重要意义。
柑橘是我国最主要的水果之一,而储藏期柑橘腐烂病对柑橘的经济效益造成了严重的影响,通常造成的损失达到20%~30%,而其中90%以上是由柑橘绿霉病菌(P.digitatum)引起的[10-11]。在柑橘绿霉病菌中,内切多聚半乳糖醛酸酶对致病性有非常重要的作用[12]。为明确柑橘绿霉病菌中其他水解酶的作用,本试验克隆柑橘绿霉病菌的木聚糖酶基因,并对其表达及功能进行分析,以阐明其在柑橘绿霉病菌致病过程中的作用,增加对柑橘绿霉病菌致病机制的了解。
1 材料与方法
1.1 材料
柑橘绿霉病菌野生型菌株Pd01(CBS 130525)分离于浙江[13],保存于本实验室;根癌农杆菌AGL-1、大肠杆菌DH5α保存于本实验室;质粒pTFCM含有T-DNA插入臂序列,其潮霉素基因由构巢曲霉强启动子PtrpC启动,由强终止子TtrpC终止,保存于本实验室;Real-time RT-PCR采用7300 real-time PCR系统;本试验使用到的引物见表1,由上海桑尼公司合成。

表1 试验中使用的引物
1.2 方法
1.2.1 柑橘绿霉病菌木聚糖酶PdXY2基因克隆
根据基因组信息设计扩增木聚糖酶基因的引物PdXY2-F和PdXY2-R(表1),以柑橘绿霉病菌野生型Pd01基因组DNA为模板扩增PdXY2基因的片段,PCR产物经1%琼脂糖凝胶电泳后,利用AxyPrepTMDNA Gel Extration Kit (Axygen,杭州)回收目的条带,将目的片段连接到克隆载体pMD18-T(TaKaRa,大连)上并送到Invitrogen公司进行测序。
1.2.2 柑橘绿霉病菌PdXY2在侵染过程中的表达分析
从市场上购买成熟温州蜜柑(Citrusnobilis),经0.1%次氯酸浸泡5~10 min后吹干待用;用无菌水洗脱PDA培养基上培养7 d的野生型菌株Pd01孢子,并将孢子稀释到1.0×106个/mL,取3 μL孢子悬浮液接种于柑橘果实的伤口上(用针簇在果皮上刺1~2 mm深的伤口),25℃保湿培养,分别在接种后间隔12 h提取发病组织的总RNA(Xygen),并反转录成cDNA(TaKaRa),使用SYBR Premix Ex TaqTM试剂盒(TaKaRa)进行real-time RT-PCR,用PdXY2-qF与PdXY2-qR引物扩增木聚糖酶基因,以γ-Actin基因(GenBank登录号:AB030227)作为内参,Actin-qF和Actin-qR引物扩增内参基因,相对表达量的计算方法参考文献[14]。
1.2.3 柑橘绿霉病菌PdXY2突变体构建
采用同源重组原理构建PdXY2基因敲除质粒,基本过程如图1:以柑橘绿霉病菌野生型菌株Pd01的基因组DNA为模板,用引物PdXY2D(含XhoI酶切位点)和PdXY2C(含SpeI酶切位点)扩增PdXY2基因3′端侧翼0.75 kb的序列,产物用限制性内切酶XhoI和SpeI消化后连接到质粒pTFCM的hph的3′端;用引物PdXY2B(含SacI酶切位点)和PdXY2A(含KpnI酶切位点)扩增PdXY2基因5′端侧翼0.82 kb的序列,产物用限制性内切酶SacI和KpnI消化回收后连接到质粒pTFCM中hph基因的5′端。PCR鉴定选择成功连接的单菌体,碱法小量抽提质粒,得到新的质粒pTFCM-XY2,将质粒电击到农杆菌AGL-1中。
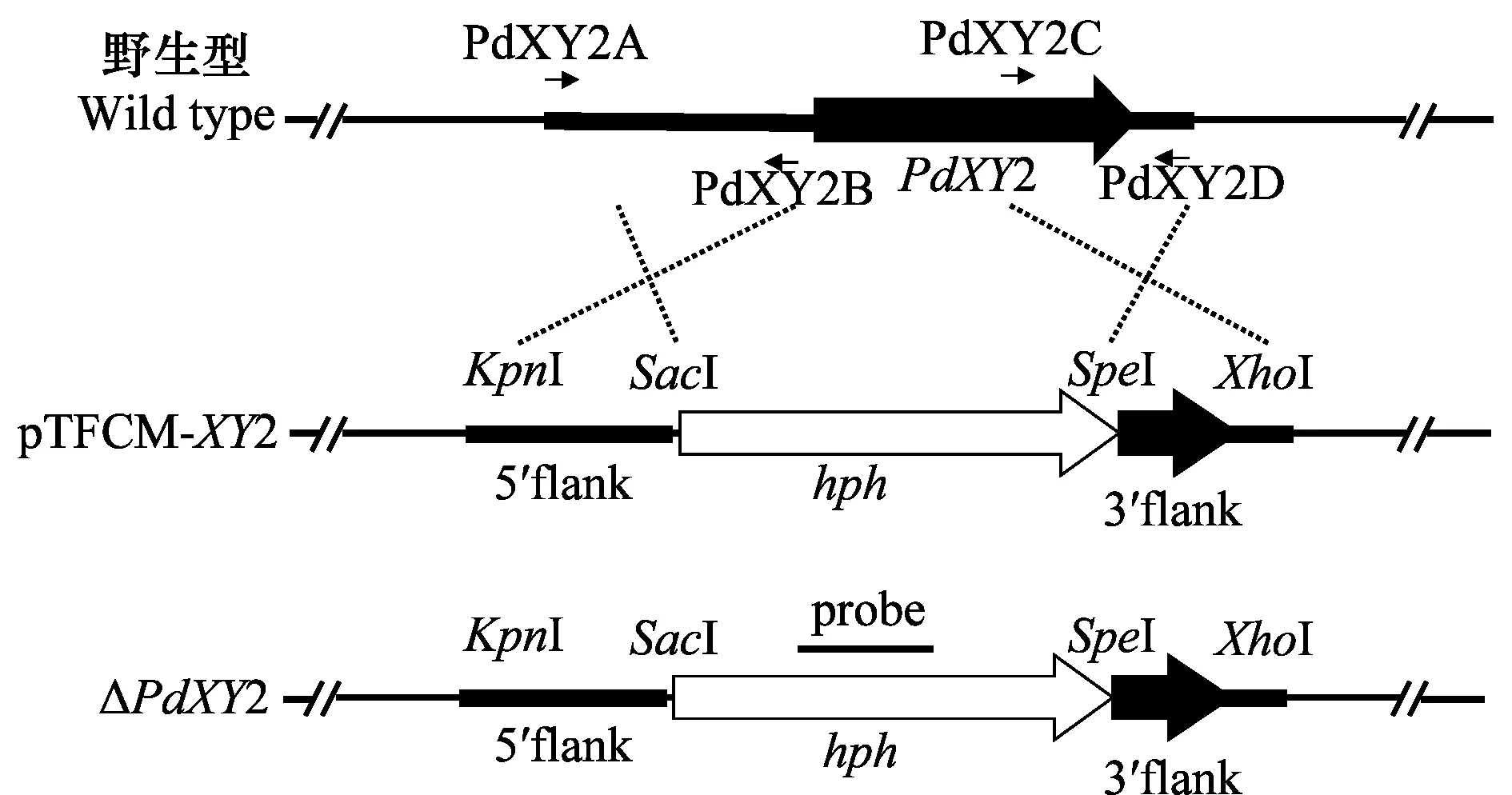
图1 PdXY2功能缺失突变株的构建Fig.1 Construction of PdXY2 knock-out mutant
1.2.4 柑橘绿霉病菌转化及转化子的分析
农杆菌转化的方法参考文献[15]。挑取含有潮霉素抗性的单菌落转入含有潮霉素的PDB培养基中,28℃培养5 d后取部分菌丝提取基因组DNA,用引物PdXY2jd-F和PdXY2jd-R进行PCR鉴定。利用Southern杂交分析PdXY2基因缺失突变株中外源插入潮霉素抗性基因的拷贝数,其步骤是用限制性内切酶HindⅢ消化野生型Pd01和ΔPdXY2的基因组DNA 10 μg,在37℃的水浴条件下酶切24 h,将经过酶切处理后的基因组DNA在1%琼脂糖凝胶中电泳12 h(电压20 V),将电泳后的基因组DNA经毛细管法转移到尼龙膜(Millipore)上,再以地高辛标记的潮霉素基因片段为探针,用DIG High Primer DNA Labeling and Detection Starter Kit II(Roche)试剂盒进行杂交,具体的操作方法详见使用说明书。
1.2.5 ΔPdXY2突变株菌丝生长及产孢能力
野生型Pd01和ΔPdXY2的菌碟制作方法见文献[16],将菌碟放在PDA中培养7 d后测量菌落直径,并用无菌水或者5%吐温洗脱孢子,将孢子稀释后用血球计数板计数孢子量,每次试验3个重复,整个试验重复3次。
1.2.6 ΔPdXY2突变株致病性分析
柑橘准备方法同上,在PDA培养基上培养野生型菌株Pd01和ΔPdXY2突变株7 d,再用无菌水洗脱孢子并稀释到1.0×106个/mL,接种方法同上,25℃保湿培养,观察发病情况,每个菌株接种约20~30个果实,整个试验重复3次以上。
2 结果与分析
2.1PdXY2基因的克隆
利用引物PdXY2-F和PdXY2-R扩增柑橘绿霉病菌的基因组,得到一条2 611 bp的核苷酸序列,将该序列在NCBI官方网站上进行比对,发现与其他物种的木聚糖酶基因具有较高的一致性;根据转录组数据测序该基因cDNA全长651 bp,含有1个大小为58 bp的内含子,能够编码出含有216个氨基酸的多肽,在NCBI中登录PdXY2基因序列,登录号为JX495171。
2.2 PdXY2蛋白分析
将柑橘绿霉病菌PdXY2编码的蛋白在NCBI进行BLAST,结果表明该蛋白与其他丝状真菌中水解酶/内切木聚糖酶有较高的一致性(图2),与Ophiostomapiliferum中内切木聚糖酶(AFR33048)的一致性为91%,与Penicilliumcamemberti中糖苷水解酶(CRL24571)的一致性为90%,与Penicilliumcanescens内切木聚糖酶(ACP27610)的一致性为81%。

图2 PdXY2蛋白与其他同源蛋白比较Fig.2 Comparison of PdXY2 with other homologous proteins
2.3PdXY2重组转化子的分析
将获得的转化子进行PCR鉴定,结果(图3a)表明,在野生型Pd01中扩增出了一条1.4 kb的条带,而在重组突变株中扩增出一条2.4 kb的条带(2 kb潮霉素抗性基因取代了PdXY2基因及其启动子区1 kb片段)。取其中一个转化子(ΔPdXY2A)进行Southern杂交鉴定,结果如图3b所示,在野生型中潮霉素标记的探针没有杂交出条带,而在ΔPdXY2突变株中则会出现一条约4.5 kb的条带,结果表明获得的重组突变株只有一条潮霉素基因序列,不存在异位插入,可以进行后续研究。
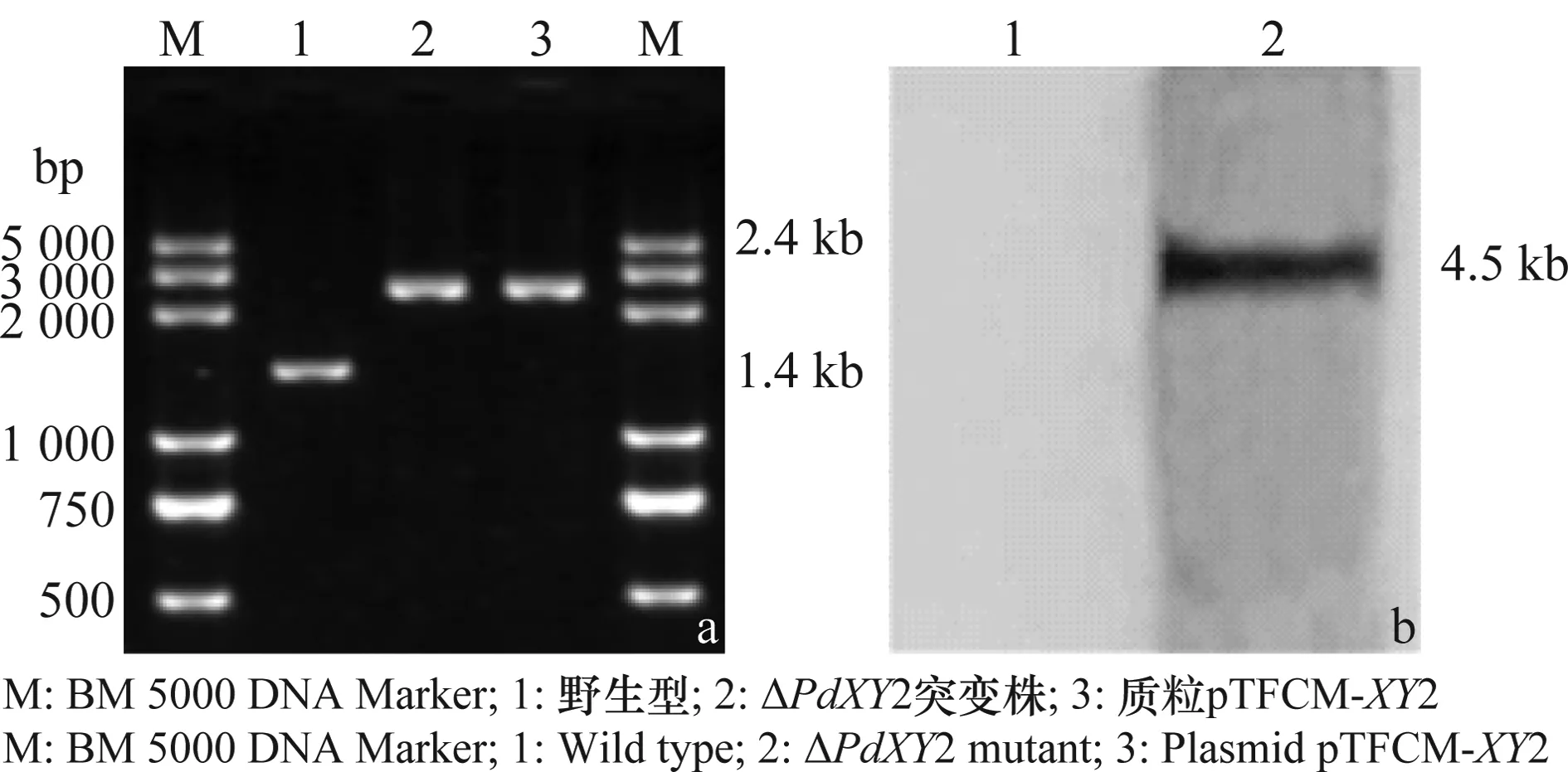
图3 ΔPdXY2突变株的PCR鉴定结果(a)和Southern鉴定结果(b)Fig.3 Identification of PdXY2-disrupted mutant by PCR (a)and Southern blot (b)
2.4PdXY2基因的表达分析
在接种柑橘的发病过程中,PdXY2基因的表达情况如图4所示,侵染初期,PdXY2基因的相对表达量呈现出一个升高趋势,在接种后48 h达到高峰,此时的相对表达量为对照的4倍,随后其表达量逐渐下降,至84 h与对照差异不大,可见在侵染过程中,PdXY2基因表现出了先升高后下降的趋势。
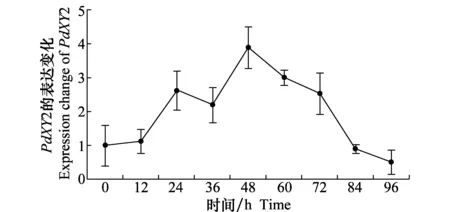
图4 在侵染柑橘过程中PdXY2的表达Fig.4 Expression of PdXY2 during infection
2.5 ΔPdXY2突变株生长及产孢分析
菌丝生长试验结果表明,ΔPdXY2突变株与野生型菌株生长速度相似,生长速率均为1.0~1.2 cm/d。产孢试验结果显示,ΔPdXY2突变株与野生型菌株的产孢量相似,在PDA平板上培养7 d后其孢子量均可达到1×108,因此,ΔPdXY2突变株与野生型菌株在菌落生长和产孢方面没有明显差异。
2.6 ΔPdXY2突变株致病性分析
致病性试验结果(图5)显示,ΔPdXY2突变株能够侵染柑橘果实,接种后培养24 h,ΔPdXY2和野生型Pd01均出现了腐烂,且病斑扩展速度差异不大,由此可见,ΔPdXY2突变株对柑橘绿霉病菌的致病性与野生型相比没有明显差异。

图5 野生型(WT)与ΔPdXY2突变株接种84 h后发病情况Fig.5 Virulence of the wild type (WT) and ΔPdXY2 mutant after 84 h of incubation
3 讨论
柑橘绿霉病菌是造成柑橘采后腐烂的最主要病原,对柑橘产业造成了严重的损失,为了增加对柑橘绿霉病菌致病分子机制的了解,本研究对柑橘绿霉病菌中的木聚糖酶基因(PdXY2)进行研究,表达分析表明PdXY2在侵染柑橘的过程中明显升高,可能起到一定的作用。ΔPdXY2突变株的致病性与野生型相比没有明显的差异,可见单一突变PdXY2基因对柑橘绿霉病菌的致病性影响不大,可能与其他同源基因相关。细胞壁水解酶含有很多同源基因,很多单一水解酶功能缺失不会引起致病性的下降[17-18]。如在赤球丛赤壳(Nectriahaematococca)中,双突变果胶裂解酶A基因和果胶裂解酶D基因能够降低致病性,然而单一突变其中任何一个基因都对致病性没有影响[19];内切多聚半乳糖醛酸酶对柑橘链格孢(Alternariacitri)引起的柑橘黑腐病致病性有影响,但是对链格孢(A.alternata)引起的褐斑病没有影响[20];在稻瘟病菌(M.oryzae)中,突变内切木聚糖酶对致病性没有影响[21];在尖孢镰刀菌(Fusariumoxysporum)中,突变外切多聚半乳糖醛酸酶对致病性没有影响[22];在玉米圆斑病菌(Cochlioboluscarbonum)中,双突变胞外多聚半乳糖醛酸酶可导致酶活下降,但是致病性与野生型差异不大[23]。此外还有一些报道也同样表明单一的突变对致病性没有影响[24-26]。
[1] Annis S L, Goodwin P H.Recent advances in the molecular genetics of plant cell wall-degrading enzymes produced by plant pathogenic fungi[J]. European Journal of Plant Pathology, 1997, 103(1): 1-14.
[2] Collmer A, Keen N T.The role of pectic enzymes in plant pathogenesis [J]. Annual Review of Phytopathology, 1986, 24: 383-409.
[3] Shieh M T, Brown R L, Whitehead M P, et al.Molecular genetic evidence for the involvement of a specific polygalacturonase, P2c, in the invasion and spread ofAspergillusflavusin cotton bolls[J]. Applied and Environmental Microbiology, 1997, 63(9): 3548-3552.
[4] ten Have A, Mulder W, Visser J, et al. The endopolygalacturonase gene Bcpg1 is required for full virulence ofBotrytiscinerea[J]. Molecular Plant-Microbe Interactions, 1998, 11(10): 1009-1016.
[5] Valette-Collet O, Cimerman A, Reignault P, et al. Disruption ofBotrytiscinereapectin methylesterase gene Bcpme 1 reduces virulence on several host plants [J]. Molecular Plant-Microbe Interactions, 2003, 16(4): 360-367.
[6] Skamnioti P, Gurr S J.Magnaporthegriseacutinase2 mediates appressorium differentiation and host penetration and is required for full virulence [J]. The Plant Cell, 2007, 19(8): 2674-2689.
[7] Yakoby N, Beno-Moualem D, Keen N T, et al.ColletotrichumgloeosporioidespelB is an important virulence factor in avocado fruit-fungus interaction [J]. Molecular Plant-Microbe Interactions, 2001, 14(8): 988-995.
[8] Yakoby N, Freeman S, Dinoor A, et al. Expression of pectate lyase fromColletotrichumgloeosporioides inC.magnapromotes pathogenicity [J]. Molecular Plant-Microbe Interactions, 2000, 13(8): 887-891.
[9] Oeser B, Heidrich P M, Muller U, et al. Polygalacturonase is a pathogenicity factor in theClavicepspurpurea/ryeinteraction [J]. Fungal Genetics and Biology, 2002, 36(3): 176-186.
[10]Kanetis L, Förster H, Adaskaveg J E.Comparative efficacy of the new postharvest fungicides azoxystrobin, fludioxonil, and pyrimethanil for managing citrus green mold[J]. Plant Disease, 2007, 91(11): 1502-1511.
[11]Macarisin D, Cohen L, Eick A, et al.Penicilliumdigitatumsuppresses production of hydrogen peroxide in host tissue during infection of citrus fruit[J]. Phytopathology, 2007, 97(11): 1491-1500.
[12]Zhang Tianyuan, Sun Xuepeng, Xu Qian, et al. The pH signaling transcription factor PacC is required for full virulence inPenicilliumdigitatum[J]. Applied Microbiology and Biotechnology, 2013, 97(20): 9087-9098.
[13]Zhu Jinwen, Xie Qingyun, Li Hongye. Occurrence of imazalil-resistant biotype ofPenicilliumdigitatumin China and the resistant molecular mechanism [J]. Journal of Zhejiang University Science A, 2006,7(S2): 362-365.
[14]Pfaffl M W, Horgan G W, Dempfle L.Relative expression software tool (REST (c)) for group-wise comparison and statistical analysis of relative expression results in real-time PCR [J]. Nucleic Acids Research, 2002, 30(9): e36.
[15]Wang Jiye, Li Hongye. Agrobacterium tumefaciens-mediated genetic transformation of the phytopathogenic fungusPenicilliumdigitatum[J]. Journal of Zhejiang University Science B, 2008, 9(10): 823-828.
[16]张志芳, 吴可嘉, 李红叶. 浙江省柑橘绿霉病菌对嘧菌酯的敏感性研究[J]. 农药学学报, 2008, 10(3): 375-378.
[17]Walton J D.Deconstructing the Cell-Wall [J]. Plant Physiology, 1994, 104(4): 1113-1118.
[18]Hamer J E, Holden D W. Linking approaches in the study of fungal pathogenesis: A commentary [J]. Fungal Genetics and Biology, 1997, 21(1): 11-16.
[19]Rogers L M, Kim Y K, Guo Wenjin, et al. Requirement for either a host-or pectin-induced pectate lyase for infection ofPisumsativumbyNectriahaematococca[J]. Proceedings of the National Academy of Sciences of the United States of America, 2000, 97(17): 9813-9818.
[20]Isshiki A, Akimitsu K, Yamamoto M, et al. Endopolygalacturonase is essential for citrus black rot caused byAlternariacitribut not brown spot caused byAlternariaalternata[J]. Molecular Plant-Microbe Interactions, 2001, 14(6): 749-757.
[21]Wu Shengcheng, Ham K S, Darvill A G, et al. Deletion of two endo-β-1,4-xylanase genes reveals additional isozymes secreted by the rice blast fungus[J]. Molecular Plant-Microbe Interactions, 1997, 10(6): 700-708.
[22]García-Maceira F I, Di Pietro A, Roncero M I. Cloning and disruption of pgx4 encoding an in planta expressed exopolygalacturonase fromFusariumoxysporum[J]. Molecular plant-Microbe Interactions, 2000, 13(4): 359-365.
[23]Scott-Craig J S, Cheng Yiqiang, Cervone F, et al. Targeted mutants ofCochlioboluscarbonumlacking the two major extracellular polygalacturonases [J]. Applied and Environmental Microbiology, 1998, 64(4): 1497-1503.
[24]Wu Shengcheng, Kauffmann S, Darvill A G, et al. Purification, cloning and characterization of two xylanases fromMagnaporthegrisea, the rice blast fungus [J]. Molecular Plant-Microbe Interactions, 1995, 8(4): 506-514.
[25]Wu Shengcheng, Halley J E, Luttig C, et al. Identification of anendo-β-1,4-D-xylanase fromMagnaporthegriseaby gene knockout analysis, purification, and heterologous expression [J]. Applied and environmental microbiology, 2006, 72(2): 986-993.
[26]Mori T, Jung H Y, Maejima K, et al.Magnaportheoryzaeendopolygalacturonase homolog correlates with density-dependent conidial germination [J]. FEMS Microbiology Letters, 2008, 280(2): 182-188.
(责任编辑:田 喆)
Preliminary study on the function of PdXY2 in Penicillium digitatum
You Shuyan1, Liu Hongxia1, Yu Ruijun1, Zheng Lijun1, Fan Shiyan1, Li Hongye2
(1. Dalian Academy of Agricultural Sciences, Dalian 116036, China;2. Institute of Biotechnology, Zhejiang University, Hangzhou 310058, China)
Penicilliumdigitatumis one of the most important pathogen causing green mold disease which impedes the development of citrus industry. In this study, we cloned a xylanase genePdXY2 and characterized its functions via knock-out strategy. The expression ofPdXY2 was up-regulated during the primary stage of infection, and reached a peak at 48 h. However,PdXY2-disrupted mutant (ΔPdXY2) showed no reduction in virulence. Taken together, our results demonstrated thatPdXY2 may play an important role in infection and single knock-out ofPdXY2 showed no reduction in virulence.
Penicilliumdigitatum; Xylanase; gene expression; gene disruption
2015-12-24
2016-02-24
国家自然科学基金(31371961); 国家现代农业产业技术体系(CARS-27)
S 436.66
A
10.3969/j.issn.0529-1542.2016.06.016
* 通信作者 E-mail:hyli@zju.edu.cn
猜你喜欢
林产化学与工业(2022年4期)2022-09-13
生物信息学(2022年1期)2022-04-01
湖南饲料(2019年5期)2019-10-15
科技创新导报(2018年1期)2018-05-07
中成药(2017年12期)2018-01-19
纺织导报(2017年4期)2017-05-05
浙江农业科学(2016年11期)2016-05-04
中国酿造(2016年12期)2016-03-01
中国畜牧兽医文摘(2015年9期)2015-12-29
中国医科大学学报(2015年10期)2015-03-01
