
添加剂对南美白对虾冻藏期间蛋白质变性的影响
2016-12-02 06:55曹淑敏赵亚石启龙
食品与发酵工业 2016年10期
曹淑敏,赵亚,石启龙
(山东理工大学 农业工程与食品科学学院,山东 淄博,255000)
添加剂对南美白对虾冻藏期间蛋白质变性的影响
曹淑敏,赵亚,石启龙*
(山东理工大学 农业工程与食品科学学院,山东 淄博,255000)
以南美白对虾为研究对象,在虾肉中分别加入10%海藻糖、10%菊糖和5%麦芽糊精,将虾肉置于-18 ℃下冻藏2个月,每隔15 d测定虾肉肌原纤维蛋白的Ca2+- ATPase活性、盐溶性蛋白含量、活性巯基含量和肌原纤维蛋白表面疏水性,探讨糖类添加剂对虾肉冻藏期间蛋白质变性的影响。结果表明:海藻糖、菊糖和麦芽糊精均抑制了冻藏过程中虾肉肌原纤维蛋白Ca2+- ATPase活性、盐溶性蛋白含量、活性巯基含量的降低和表面疏水性升高的程度,延缓了南美白对虾肉冻藏期间蛋白质的冷冻变性。3种添加剂中,抗冻效果高低顺序依次为:海藻糖、菊糖、麦芽糊精,其中添加海藻糖与菊糖处理间无显著差异(P>0.05),但两者的抗冷冻变性效果均显著高于添加麦芽糊精处理组(P<0.05)。基于玻璃化转变理论,阐明了3种添加剂对虾肉抗冻效果的机制,为水产品抗冷冻变性剂的筛选提供理论依据。
南美白对虾;蛋白质变性;肌原纤维蛋白;生化特性
南美白对虾(PenaeusvannameiBoone)是世界养殖产量最高的三大虾类之一,具有丰富的营养价值与保健功能。南美白对虾蛋白质和水分含量高,内源性蛋白酶活性较强,蛋白质降解作用较快。目前,鲜虾冷冻是维持其品质的常用方法,但在冻藏过程中经常出现蛋白质变性,导致虾肉持水力和凝胶能力降低[1]。添加抗冻剂是缓解虾肉冷冻变性常采用的方法。李姣等[2]、李学鹏等[3]研究表明,Ca2+-ATPase活性、疏水性与活性巯基可以作为蛋白质变性程度的判定指标。马璐凯等[4]以焦磷酸钠为对照,研究了海藻糖、海藻胶与寡糖对虾肉蛋白质冷冻变性的影响,结果表明,3种添加剂能有效降低冷冻虾仁解冻汁液流失,防止肌原纤维蛋白Ca2+-ATPase活性降低。糖类作为抗冷冻保护剂在水产品冻藏过程中使用范围较广泛,工业上常用抗冻剂为蔗糖与山梨糖醇或者两者混合物,但其甜度和热量较高[5]。菊糖可促进肠道益生菌增殖,而且热量低,适合糖尿病、肥胖症患者等特殊人群食用[6]。海藻糖甜度与热量较低,而且具有提高机体免疫、抗肿瘤、抗辐射、降血脂等生物活性[7]。麦芽糊精甜度低、易消化,在食品工业中常用于乳化剂、增稠剂和成膜剂[8]。石启龙等[5,9]研究表明,添加菊糖、海藻糖、麦芽糊精可提高南美白对虾肉的最大冷冻浓缩溶液时的玻璃化转变温度,缩短最大冰晶形成带区域,理论上可提高虾肉贮藏稳定性,但有待于进一步验证。基于此,本文研究了海藻糖、菊糖和麦芽糊精对虾肉冻藏期间肌原纤维蛋白生化特性影响,为虾肉新型抗冻剂筛选提供理论依据。
1 材料与方法
1.1 材料与仪器
鲜活南美白对虾,2014.12月购于淄博市海盛水产品市场,长度(10.50±0.65) cm,质量(11.24±1.50) g。冰温猝死后,置于- 76℃冰箱冻藏待用。主要试剂:海藻糖、菊糖、麦芽糊精,食品级;牛血清蛋白、三羟甲基氨基甲烷(Tris)、马来酸(maleate)、三氯乙酸(TCA)、Elon试剂、二硫-2-硝基苯甲酸(DTNB)、三磷酸腺苷二钠、8-苯胺基-1-萘磺酸钠、钼酸铵、Na2HPO4、NaH2PO4、十二烷基硫酸钠、浓H2SO4、NaOH、HCl、KCl、Na2SO3等,分析纯。
DW-FL253型低温冰箱,中科美菱低温科技有限公司;GL-20G-Ⅱ型冷冻离心机,上海安亭科学仪器厂;T18 basic型分散机,德国IKA公司;UV-2102型紫外-可见分光光度计,尤尼柯(上海)仪器有限公司;970CRT荧光分光光度计,上海精密科学仪器有限公司。
1.2 实验方法
南美白对虾4 ℃解冻,去头、壳,将虾肉平均分成4份,按虾肉质量分别加入10%海藻糖(PV-T)、10%菊糖(PV-I)和5%麦芽糊精(PV-MD),以纯虾肉(PV)作为对照。将4份虾肉分别用搅拌机搅匀,然后密封于自封袋中,置于-18 ℃贮藏2个月,每隔15d取样,进行肌原纤维蛋白生化特性指标测定。
1.3 指标测定方法
1.3.1 肌原纤维蛋白提取
采用马璐凯等[4]的方法,略作修改。准确称取虾肉2.00 g,置于离心管中,加入20 mL冷却的Tris-maleate缓冲液(0.05 mol/L KCl-20 mmol/L Tris-maleate,pH=7.0),低温均质2 min;然后4 ℃、10 000 r/min下离心20 min,移除上清液,沉淀洗涤2~3次,收集沉淀,然后加入20 mL 冷却的Tris- maleate缓冲液(0.6 mol/L KCl-20 mmol/L Tris-maleate,pH=7.0),低温均质2 min。均质后样品在温度4 ℃下提取60 min,然后在温度4 ℃、10 000 r/min离心20 min,用纱布过滤所得上清液即为肌原纤维蛋白溶液。
1.3.2 肌原纤维蛋白含量
采用双缩脲法测定[2]。
1.3.3 盐溶性蛋白含量
参考胡亚琴等[10]方法。
1.3.4 Ca2+-ATPase活性
首先绘制无机磷酸盐溶液的标准曲线,用于无机磷的定量分析[11]。Ca2+-ATPase活性测定参考万建荣等方法[12],略作修改。按表1中比例配制反应体系,25 ℃反应10 min,加入1 mL 15%的TCA终止反应,将混合液6 500 r/min离心5 min,取上清液1.0 mL,分别加入1.0 mL硫酸钼酸溶液、0.50 mL Elon试剂和2.5 mL蒸馏水,混匀后25 ℃发色45 min,640 nm测定吸光度值(A)。空白组先加入1.0 mL 15% TCA使酶失活,再加ATP溶液,其它操作同上,640 nm测定吸光度值(B)。根据标准曲线求出磷酸量。Ca2+-ATPase活性计算公式:
Ca2+-ATPase活性=(A-B)/t·酶蛋白质量
(1)
式中:A为1.0 mL反应液中生成的磷酸量,μmol;B为空白值,μmol;t为反应时间,min;酶蛋白质量为1.0 mL反应液所含的酶量,mg。
1.3.5 活性巯基
参考缪函霖等[13]的方法,略作修改。取1.0 mL肌原纤维蛋白溶液(4 mg/mL),加入9 mL 0.20 mol/L的Tris-HCl缓冲液(2% SDS和10 mmol/L EDTA,pH=6.8),取4 mL混合液,加入0.4 mL、pH 8.0的0.2 mol/L Tris-HCl(含0.1% DTNB)。混合液40℃反应25 min,412 nm处测定吸光度,空白采用0.6 mol/L KCl取代样品。计算公式为:
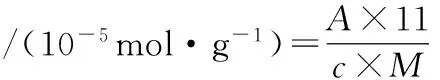
(2)
式中:A为412 nm处的吸光度值;11为稀释倍数;c为分子吸光系数,其值为13 600 L/(mol·cm);M为肌原纤维蛋白含量,mg/mL。

表1 Ca2+-ATPase活性反应体系
1.3.6 表面疏水性
采用荧光探针法测定表面疏水性[14]。
2 结果与分析
2.1 Ca2+-ATPase活性
Ca2+-ATPase活性是肌肉蛋白的重要属性,是评价肌球蛋白分子完整性的重要指标,被广泛用于评价肌肉蛋白质的变性[15]。虾肉冻藏过程中肌原纤维蛋白Ca2+-ATPase活性的变化如图1所示。
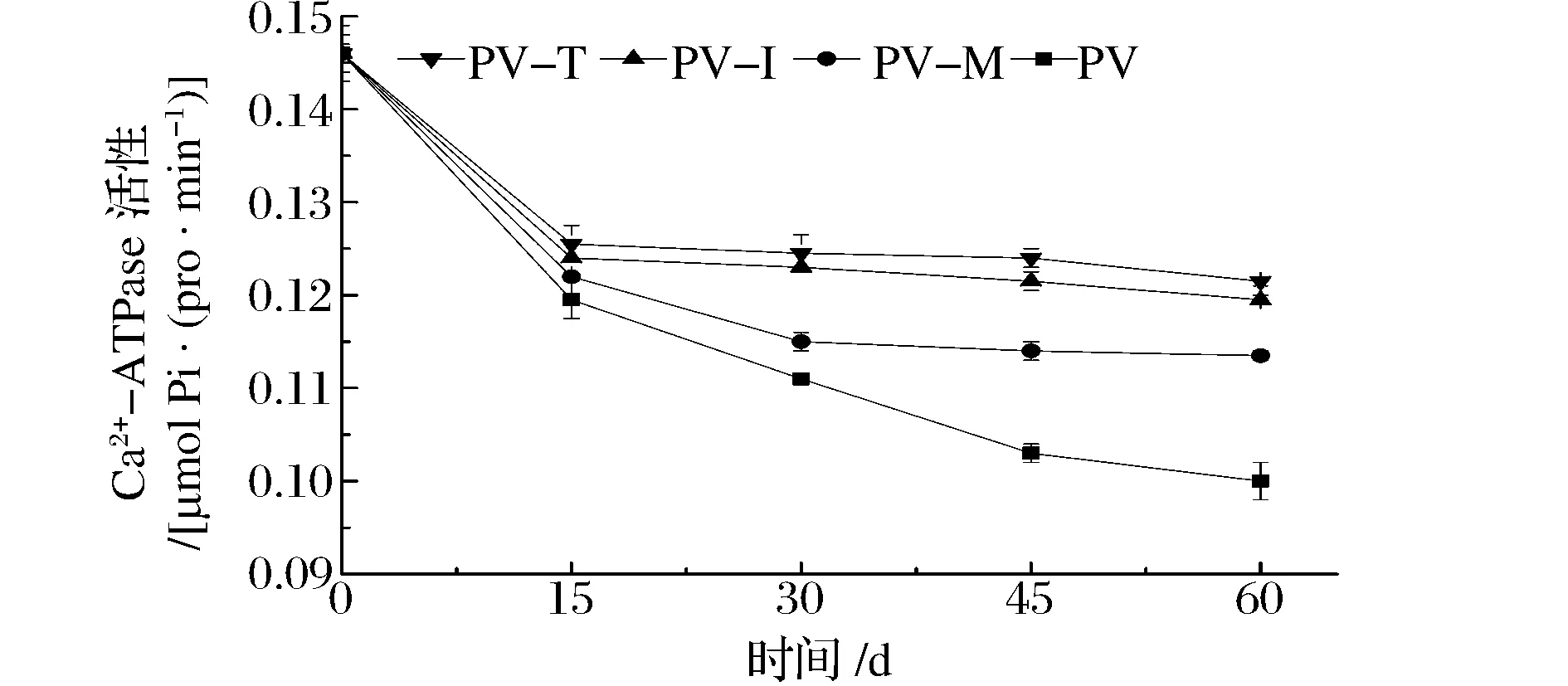
图1 冻藏过程中肌原纤维蛋白Ca2+-ATPase活性的变化Fig.1 Ca2+-ATPase activity change in myofibrillar protein during frozen storage
虾肉Ca2+-ATPase活性随贮藏时间延长而下降。冻藏前期,虾肉Ca2+-ATPase活性下降速度较快;冻藏中、后期,虾肉Ca2+-ATPase活性呈缓慢下降趋势。例如,冻藏15 d和60 d时,PV的Ca2+-ATPase活性分别降低了18.2%和31.5%。BENJAKUL等[16]、NOPIANTI等[17]分别对蟹肉、金线鱼鱼糜冻藏期间肌原纤维蛋白Ca2+-ATPase活性进行了研究,发现了类似的规律。Ca2+-ATPase活性源于肌球蛋白的球状头部区域,冻藏期间,虾肉肌原纤维蛋白的Ca2+-ATPase活性都发生了不同程度的下降,说明肌原纤维蛋白发生了不同程度的变性,尤其是肌球蛋白的头部区域。Ca2+-ATPase活性在冻藏前期下降速度较快可能是由于肌球蛋白头部区域变性所致。3种添加剂具有延缓Ca2+-ATPase活性下降的作用。例如,虾肉贮藏15d时,PV-T、PV-I和PV-M的Ca2+-ATPase活性分别下降了14.0%、15.1%和16.4%;冻藏60 d时,Ca2+-ATPase活性分别下降了16.7%、18.2%和22.3%。
2.2 盐溶性蛋白含量
肌动球蛋白的溶解度是评价肌肉蛋白质变性的常用指标。南美白对虾在冻藏过程中盐溶性蛋白含量随贮藏时间变化如图2所示。
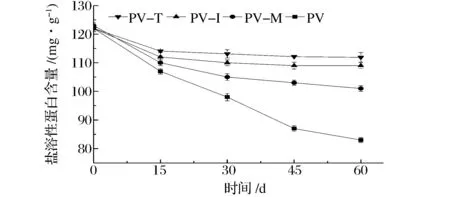
图2 冻藏过程中肌原纤维蛋白盐溶性蛋白含量的变化Fig.2 Salt extractable protein content change in myofibrillar protein during frozen storage
可以看出,盐溶性蛋白的含量随着冻藏时间的延长呈下降趋势,尤其是冻藏初始阶段下降幅度明显,冻藏后期呈缓慢下降趋势。冻藏15、30、45和60 d时,PV的盐溶蛋白含量分别降低了12.3%、19.7%、28.7%和32.0%。XIONG[18]、WU等[19]研究了鲤鱼鱼糜、鳙鱼冻藏期间盐溶蛋白变化规律,得到了类似的结论。添加海藻糖、菊糖和麦芽糊精显著抑制了虾肉盐溶蛋白含量下降的速度和幅度。冻藏15、30、45和60d时,PV-I盐溶蛋白含量分别降低了8.2%、9.8%、10.7%和10.7%。相同冻藏时间时,添加海藻糖和菊糖虾肉的盐溶蛋白含量要显著高于添加麦芽糊精的虾肉。例如,冻藏45d时,PV-T、PV-I和PV-M盐溶蛋白含量分别降低了8.1%、10.7%和15.6%。路钰希等[20]研究了鱿鱼冻藏期间盐溶蛋白变化规律基本一致。冻藏过程中,由于肌原纤维蛋白部分结合水形成冰晶,导致肌动球蛋白分子之间能够相互形成非共价键,进而形成超大分子的不溶性凝集体,使其溶出量在冻藏过程中不断下降[21]。此外,肌原纤维蛋白质变性后会产生一种在高离子强度下不能溶出,但在碱性条件下可以溶出的蛋白质,也会导致肌动球蛋白在冻藏过程中溶出量的下降[3]。KITTIPHATTANABAWON等[22]认为盐溶蛋白含量下降可能与疏水作用、二硫键以及离子相互作用有关。
2.3 活性巯基含量
南美白对虾冻藏过程中肌原纤维蛋白活性巯基含量随冻藏时间的变化如图3所示。随着冻藏时间的延长,虾肉活性巯基的含量呈下降趋势。冻藏初始阶段,虾肉的活性巯基含量下降速度较快。冻藏15 d和60 d时,PV的活性巯基含量分别减少了26.6%和44.8%。添加海藻糖、菊糖和麦芽糊精能显著抑制虾肉活性巯基含量下降的速度和幅度。冻藏15、30、45和60 d时,PV-T活性巯基含量分别降低了17.6%、19.7%、20.5%和21.8%。相同冻藏时间时,添加海藻糖和菊糖虾肉的盐溶蛋白含量要显著高于添加麦芽糊精的虾肉。例如,冻藏60 d时,PV-T、PV-I和PV-M盐溶蛋白含量分别降低了21.8%、24.1%和31.4%。WANG等[15]研究表明,草鱼活性巯基含量随随冻藏时间的延长呈下降趋势,添加魔芋葡甘聚糖水解物能抑制活性巯基含量的降低,这与本文研究结果基本一致。
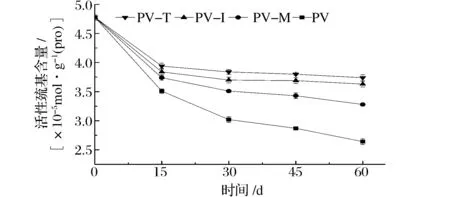
图3 冻藏过程中肌原纤维蛋白活性巯基含量的变化Fig.3 Reactive sulfhydryl content change in myofibrillar protein during frozen storage
巯基是肌球蛋白分子中的活性基团,位于肌动球蛋白头部区域,巯基含量降低与肌动球蛋白头部区域结构的改变有关。通过比较可知,活性巯基和Ca2+-ATPase活性的下降幅度要高于盐溶性蛋白含量的下降幅度,这可能是肌原纤维蛋白变性的过程中,肌动球蛋白的头部更易受到影响。低温贮藏过程中,虾肉蛋白质发生变性,肌球蛋白的分子结构特别是头部区域发生构象的变化,使埋藏在蛋白质分子内部的活性巯基暴露出来,进而被氧化成二硫键,导致巯基含量的减少[10,14-15]。本研究中,活性巯基含量的变化与Ca2+-ATPase活性的变化趋势基本一致,由此可以推测冻藏过程中,虾肉肌球蛋白的构象尤其是头部区域发生了变化,导致了蛋白质变性。海藻糖、菊糖和麦芽糊精的添加可较好地抑制南美白对虾肉肌原纤维蛋白构型的变化,防止活性巯基的暴露,抑制冻藏过程中巯基氧化和二硫键的形成。
2.4 表面疏水性
蛋白质的表面疏水性反映的是蛋白质分子表面疏水性氨基酸的相对含量,可用之衡量蛋白质的变性程度。南美白对虾在冻藏过程中肌原纤维蛋白的表面疏水性随时间变化如图4所示。可以看出,虾肉蛋白质表面疏水性随着冻藏时间的延长呈上升趋势。这与WANG等[15]、ZHAO等[23]对草鱼、大黄鱼变化规律基本一致。添加海藻糖、菊糖和麦芽糊精能显著抑制虾肉蛋白质表面疏水性升高的幅度。冻藏15、30、45和60d时,PV的表面疏水性分别增加了43.8%、51.2%、59.4%和68.3%;而PV-I的表面疏水性分别增加了12.2%、14.7%、19.7%和25.0%。3种添加剂中,海藻糖和菊糖抑制表面疏水性增加的效果要好于麦芽糊精,而且冻藏过程中,蛋白质表面疏水性上升幅度较为平缓。冻藏的过程中,蛋白质的空间结构发生了变化,导致肌原纤维蛋白质分子伸展开,从而使亲水基团和疏水基团的相对位置发生变化,非极性的氨基酸残基暴露在蛋白质分子的表面,使蛋白质的疏水性增加[3,14]。
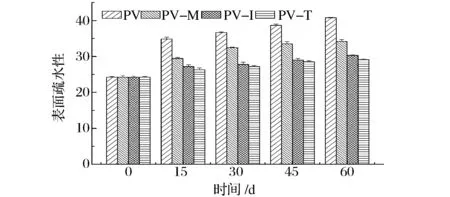
图4 冻藏过程中肌原纤维蛋白表面疏水性的变化Fig.4 Surfacehydrophobicity change in myofibrillar protein during frozen storage
3 讨论
冷冻是水产品保藏常用方法,但在冷冻过程中会逐渐发生冰结晶成长、重结晶、蛋白质变性等引起品质劣变的反应。这些受分子扩散运动控制的变化反应与玻璃化转变密切相关。鲜活水产品冻结过程中,随着温度降低,冰晶体的数量逐渐增多,液相中溶质浓度和体系黏度逐渐增加,当温度降低到一定程度,液相不再结冰,此时体系状态为玻璃态,对应的温度为最大冷冻浓缩溶液时的玻璃化转变温度(Tg′)。根据玻璃化转变理论,当贮藏温度低于Tg′时,体系处于玻璃态,分子间流动受到限制,各种理化反应受到抑制;反之,当贮藏温度高于Tg′时,体系处于橡胶态,分子流动性加强,受分子扩散运动控制的各种化学反应速度加快[24]。AGUSTINI等[25]研究表明,金枪鱼肉在低于-84 ℃贮藏时,体系处于玻璃态,鱼肉中各种分子移动速率显著降低,酶促反应受到抑制,导致鱼肉三磷酸腺苷(ATP)降解速率受到抑制。AKKÖSE和AKTA[26-27]研究表明,虹鳟鱼肉在高于玻璃化转变温度(Tg)贮藏6个月,总挥发性盐基氮(TVB-N)和硫代巴比妥酸反应物(TBARS)值较高,鱼肉劣变速度加快;而在低于Tg贮藏时,鱼肉保持较高品质。添加抗冻剂是目前有效的抗鱼肉蛋白质冷冻变性的方法。OSAKO等[28]研究报道,添加海藻糖可使冷冻鱼糜冻藏期间保持较高的Ca2+-ATPase活性,而且鱼糜中的非冻结水含量显著增加。CARVAJAL等[29]研究了不同分子量的麦芽糊精对狭鳕鱼糜蛋白质抗冻作用的影响。结果表明,所有分子量的麦芽糊精都具有很好的抗冻效果。高分子量的麦芽糊精在低温冻藏时抗冻效果显著,这可能是低温下水分子的流动性降低的缘故。石启龙等[5, 9]研究表明,南美白对虾肉的最大冷冻浓缩溶液时的玻璃化转变温度Tg′随着海藻糖、菊糖、麦芽糊精的添加而提高,且提高效果海藻糖和菊糖高于麦芽糊精,而海藻糖和菊糖间差别不大[5, 9]。此外,南美白对虾肉的最大冰晶形成带的区域随糖类添加而缩短[2, 27]。因此,糖类尤其是菊糖和海藻糖添加能显著提高南美白对虾肉的贮藏稳定性,这与本文研究结论基本吻合,进一步验证了玻璃化转变理论的正确性。
4 结论
(1)添加海藻糖、菊糖和麦芽糊精均抑制了冻藏过程中虾肉肌原纤维蛋白Ca2+- ATPase活性、盐溶性蛋白含量、活性巯基含量的降低和表面疏水性升高的速度和程度,延缓了南美白对虾肉冻藏期间蛋白质的冷冻变性。
(2)虾肉中添加菊糖、海藻糖时,其抗冷冻变性效果要高于添加麦芽糊精处理组,但是菊糖与海藻糖处理组间差异不显著。
(3)基于玻璃化转变理论,揭示了菊糖、海藻糖与麦芽糊精提高虾肉抗冻作用以及3种不同添加剂间抗冻效果间的差异的机制,为水产品尤其是南美白对虾肉的抗冷冻变性剂的筛选提供理论依据。
[1] 林雯雯,石启龙,牛东泽,等. 预处理对冻干南美白对虾仁贮藏品质的影响[J]. 食品工业科技,2014,35(15):330-333.
[2] 李姣,李学鹏,励建荣,等. 冷藏条件下中国对虾肌肉蛋白质的生化特性[J]. 食品科学,2011,32(5): 16-21.
[3] 李学鹏,陈杨,王金厢,等. 中国对虾冷藏过程中肌肉组织结构与蛋白质生化性质的变化[J]. 中国食品学报,2014,14(5): 72-79.
[4] 马璐凯,张宾,王强,等. 海藻糖、海藻胶及寡糖对南美白对虾蛋白质冷冻变性的抑制作用[J]. 现代食品科技,2014,30(6): 140-145.
[5] 石启龙,赵亚,曹淑敏,等. 糖类对南美白对虾肉玻璃化转变温度与状态图的影响[J].农业机械学报,2015,46(11):236-244.
[6] 苗晓洁,董文宾,代春吉,等. 菊糖的性质、功能及其在食品工业中的应用[J]. 食品科技,2006,31(4):9-11.
[7] Pawar S N, Edgar K J. Alginate derivatization: A review of chemistry, properties and applications [J]. Biomaterials,2012,33(11):3 279-3 305.
[8] 刘文慧,王颉,王静,等. 麦芽糊精在食品工业中的应用现状[J]. 中国食品添加剂,2007(2):183-186.
[9] SHI Q L, LIN W W, ZHAO Y, et al. Thermal characteristics and state diagram of Penaeus vannamei meat with and without maltodextrin addition [J]. Thermochimica Acta, 2015, 616: 92-99.
[10] 胡亚芹,胡庆兰,杨水兵,等. 不同冻结方式对带鱼品质影响的研究[J]. 现代食品科技,2014,30(2): 23-30.
[11] 李姣. 中国对虾贮藏过程中肌肉蛋白质生化特性变化规律研究 [D]. 杭州:浙江工商大学,2011.
[12] 万建荣,洪玉菁,奚印慈,等编译. 水产食品化学分析手册[M].上海:上海科技出版社,1993.
[13] 缪函霖,包海蓉,赵路漫. 金枪鱼肉冷藏过程中理化特性的变化[J]. 江苏农业科学,2013,41(8): 248-251.
[14] LIU Q, CHEN Q, KONG B H, et al. The influence of superchilling and cryoprotectants on protein oxidation and structural changes in the myofibrillar proteins of common carp (Cyprinuscarpio) surimi [J]. LWT-Food Science and Technology, 2014, 57(2): 603-611.
[15] WANG L, XIONG G Q, PENG Y B, et al. The cryoprotective effect of different konjac glucomannan (KGM) hydrolysates on the glass carp (Ctenopharyngodonidella) myofibrillar during frozen storage [J]. Food and Bioprocess Technology, 2014, 7(12): 3 398-3 406.
[16] BENJAKUL S, SUTTHIPAN N. Muscle changes in hard and soft shell crabs during frozen storage [J]. LWT-Food Science and Technology, 2009, 42(3): 723-729.
[17] NOPIANTI R, HUDA N, NORYATI I, et al. Cryoprotective effect of low-sweetness additives on protein denaturation of threadfin bream surimi (Nemipterusspp.) during frozen storage [J]. CyTA-Journal of Food, 2012, 10(3): 243-250.
[18] XIONG G Q, CHENG W, YE L X, et al. Effects of konjac glucomannan on physicochemical properties of myofibrillar protein and surimi gels from grass carp (Ctenopharyngodonidella) [J]. Food Chemistry, 2009, 116(2): 413-418.
[19] WU H, WANG Z Y, LUO Y K, et al. Quality changes and establishment of predictive models for bighead carp (Aristichthys nobilis) fillets during frozen storage [J]. Food and Bioprocess Technology, 2014, 7(12): 3 381-3 389.
[20] 路钰希,沈萍,李学英,等. 保鲜剂对冻藏鱿鱼品质变化的影响[J]. 食品工业科技,2014,35(19): 274-279.
[21] 崔珺. 速冻方式和贮藏温度对带鱼品质影响的研究[D]. 舟山: 浙江海洋学院,2011.
[22] KITTIPHATTANABAWON P, BENJAKUL S, VISESSANGUAN W, et al. Cryoprotective effect of gelatin hydrolysate from blacktip shark skin on surimi subjected to different freeze-thaw cycles[J]. LWT-Food Science and Technology, 2012, 47(2): 437-442.
[23] ZHAO J, LYU W J, WANG J L, et al. Effects of tea polyphenols on the post-mortem integrity of large yellow croaker (Pseudosciaenacrocea) fillet proteins [J]. Food Chemistry, 2013, 141(3): 2 666-2 674.
[24] 刘红英,高瑞昌,戚向阳. 食品化学[M].北京:中国质检出版社,2013.
[25] AGUSTINI T W, SUZUKI T, HAGIWARA T, et al. Change of K value and water state of yellowfin tunaThunnusalbacaresmeat stored in a wide temperature range (20℃ to-84℃) [J]. Fisheries Science, 2001, 67(2): 306-313.
[26] AKKÖSE A, AKTAN. Determination of glass transition temperature of rainbow trout (Oncorhynchusmykiss) and effects of various cryoprotective biopolymer blends on some chemical changes [J]. Journal of Food Processing and Preservation, 2009, 33(5): 665-675.
[28] OSAKO K, HOSSAIN M A, KUWAHARA K, et al. Effect of trehalose on the gel- forming ability, state of water and myofibril denaturation of horse mackerelTrachurusjaponicussurimi during frozen storage [J]. Fisheries Science, 2005, 71(2): 367-373.
[29] CARVAJAL P A, MACDONALD G A, LANIER T C. Cryostabilization mechanism of fish muscle proteins by maltodextrins [J]. Cryobiology, 1999, 38(1): 16-26.
Effects of additives on protein denaturation of Penaeus vannamei during frozen storage
CAO Shu-min, ZHAO Ya, SHI Qi-long*
(School of Agricultural Engineering and Food Science, Shandong University of Technology, Zibo 255000, China)
Penaeusvannameiwere used to study the effect of cryoprotective on denaturation of frozen seafood.Penaeusvannameimeat with addition of 10 % trehalose, 10 % inulin and 5% maltodextrin were stored at -18 ℃ for up to 2 months. Ca2+-ATPase activity, salt solubility, reactive sulfhydryl content and surface hydrophobicity of myofibrillar protein were analyzed every 15 days during the frozen storage. The results showed that addition of trehalose, inulin and maltodextrin inhibited the decrease of Ca2+-ATPase activity, salt solubility, reactive sulfhydryl content and rising trend of surface hydrophobicity. The additives delayed the degree of protein denaturation during frozen storage ofPenaeusvannameiand the order of cryoprotective effect from high to low is: trehalose, inulin and maltodextrin. Cryoprotective effect between trehalose and inulin had no significant (P>0.05), they were both better than maltodextrin treatment (P<0.05). The cryoprotective effect of three additives onPenaeusvannameiwas interpreted based on glass transition theory, which will offer theoretical foundation for screening of cryoprotective agent.
Penaeusvannamei;protein denaturation;myofibrillar protein;biochemical properties
10.13995/j.cnki.11-1802/ts.201610017
硕士研究生(石启龙教授为通讯作者,E-mail:qilongshi@sdut.edu.cn)。
国家自然科学基金资助项目(31171708)
2016-01-29,改回日期:2016-03-24
猜你喜欢
食品工业科技(2022年19期)2022-09-27
安徽化工(2021年2期)2021-05-15
食品工业科技(2019年15期)2019-08-28
食品工业科技(2018年19期)2018-10-22
安徽农业科学(2018年8期)2018-05-14
饮食与健康·下旬刊(2017年1期)2017-02-08
大连工业大学学报(2015年4期)2015-12-11
北京青年周刊(2015年7期)2015-09-10
食品工业科技(2014年23期)2014-03-11
食品工业科技(2014年23期)2014-03-11
