
谷瘟病菌生理小种鉴别及谷子标准品种体系的构建
2016-12-01 08:10李志江贾冠清李祥羽李易初马金丰李艳东刁现民
中国农业科学 2016年17期
李志江,贾冠清,李祥羽,李易初,马金丰,智 慧,汤 沙,张 硕,柴 杨,李艳东,刁现民
(1黑龙江省农业科学院作物育种研究所,哈尔滨 150086;2中国农业科学院作物科学研究所,北京 100081;3黑龙江省农业科学院植物保护研究所,哈尔滨 150086)
谷瘟病菌生理小种鉴别及谷子标准品种体系的构建
李志江1,贾冠清2,李祥羽1,李易初3,马金丰1,智 慧2,汤 沙2,张 硕2,柴 杨2,李艳东1,刁现民2
(1黑龙江省农业科学院作物育种研究所,哈尔滨 150086;2中国农业科学院作物科学研究所,北京 100081;3黑龙江省农业科学院植物保护研究所,哈尔滨 150086)
【目的】谷瘟病是谷子生产中的重要病害,确定谷瘟病菌不同生理小种对谷子抗病资源鉴定、抗病品种培育和生产上不同抗病品种的合理配置有重要意义。构建谷瘟病小种鉴别的谷子品种体系,为鉴别不同谷瘟病生理小种奠定基础。【方法】用采集自不同谷子产区的10份谷瘟病菌对苗期谷子核心种质资源的60份代表品种进行接种鉴定,分别对不同菌株进行致病性和谷子品种抗、感敏感性判定,按照谷子谷瘟病0—9级法划分标准进行划分,将不同的抗性结果分别记录为高抗、抗病、中抗、感病和高感5个抗病水平,其中,高抗、抗病和中抗记作0,感病和高感记作1,将抗感性结果转化为0/1数据,用NTSYS软件构建进化树,根据品种的抗感性分类,选择最少的品种数量构建谷瘟病小种鉴定的标准品种体系。【结果】根据谷子资源对接种病菌的抗感反应差异,用NTSYS软件开展谷瘟病菌致病性聚类分析,以相似系数0.70为界,将10个谷瘟病菌株分为3类,将供试品种对10个谷瘟病菌株的抗感反应数据同样用NTSYS软件进行聚类分析,以相似系数0.65为界,将谷子品种资源划分为5类,并筛选出单皮粘、金屏谷子、锦谷5号、大毛毛谷、龙爪谷、假金苗和牛头谷等7个品种作为谷瘟病小种鉴别品种体系。同时筛选出黄棒头、齐头黄、单皮粘、白谷、维子那谷和郑谷4号等6个抗广谱菌株的品种,可以作为谷子抗病育种的抗性基因来源。【结论】建立了一套谷瘟病生理小种鉴别的品种体系,并为谷子抗谷瘟病育种筛选出多份广谱抗性基础材料。
谷子;谷瘟病;核心种质资源;生理小种;抗病性
0 引言
【研究意义】谷瘟病菌(Pyricularia setariae Nisikado)对谷子(Setaria italica L.)品种的致病性与水稻稻瘟病菌一样,符合基因对基因假说,均具有小种特异性[1]。谷瘟病是中国谷子主产区的主要病害之一[2-3],可造成谷子生产严重减产甚至绝收[4-7]。谷瘟病生理小种的鉴定和划分有利于弄清谷瘟病菌小种类群、区系分布以及谷瘟病菌消长变化规律等,为选育和应用抗病品种以及合理布局和更换品种提供指导。【前人研究进展】谷子在整个生长期都可感染谷瘟病菌,分别引起苗瘟、叶瘟、节瘟、穗颈瘟和穗瘟,其中,叶瘟和穗瘟造成的减产程度最大。20世纪70年代和80年代,吉林省发生大面积的谷瘟病,仅德惠、九台、扶余三县受害面积就达1万公顷,一般减产20%—30%,严重的减产50%—70%,损失粮食达4 000吨[4-5,7]。进入21世纪,谷瘟病仍然危害严重,并呈现出同锈病、纹枯病等病害交替爆发的现象。近几年,谷瘟病发病有加重的趋势:河北、河南、山东等夏谷种植区域,谷瘟病和锈病混合发病,在谷子生产上造成严重减产,高感谷瘟病品种的推广,更加剧了谷瘟病的发生。选择抗病品种是防治谷瘟病最经济、有效的措施,但是单一抗病品种长时间大面积种植容易对田间病菌形成选择压[8],导致生理小种发生变异,寄主抗性减退甚至丧失。因此,需要了解谷瘟病菌田间生理小种组成,鉴定推广品种的抗性,同时挖掘新的抗病资源用于谷瘟病抗病育种。中国农业科学院、吉林省农业科学院、黑龙江省农业科学院等多家育种单位,自20世纪70年代以来先后对不同地区的种质资源和优良品种进行了抗谷瘟病鉴定,筛选出一部分抗谷瘟病材料[9-11]。【本研究切入点】1985年,YAN等[5]用6个谷子鉴别品种将采自10个省份的711个谷瘟病单孢分离菌分为7群32个小种,这是谷子上仅有的1套谷瘟病小种鉴别体系[3],且由于时间太久,该体系6个谷子品种的确切来源和基因型已无从考证,已不能够满足目前谷瘟病生理小种鉴定的需要。JIA等[12]构建了首个谷子核心种质群体,该群体涵盖了国内谷子主要栽培区域的农家品种和育成品种以及国外搜集的材料,最能代表谷子遗传群体结构,为谷瘟病抗性鉴定标准品种的筛选提供了基础。【拟解决的关键问题】本研究用采自谷子主产区的10个谷瘟病单孢菌对60份选自谷子核心种质资源的品种进行苗期接种鉴定,通过聚类和抗性分析,构建谷瘟病生理小种鉴别的标准品种体系,解决谷瘟病小种鉴定缺乏统一的标准品种体系的问题,对摸清中国谷子主产区谷瘟病主要生理小种的分布,以及开展抗病育种和有针对性地进行抗病品种布局都具有重要的意义。
1 材料与方法
1.1 材料来源
供试的60份谷子核心种质资源由中国农业科学院作物科学研究所提供,包含国内谷子主要栽培区域的农家品种和育成品种以及部分国外搜集的材料(表1)。
采用温室盆栽的方法进行鉴定。将谷子播种在装有肥沃黑土的营养钵内,每个营养钵种植1份材料,每份材料挑取籽粒饱满的谷粒20粒,均匀播种在营养钵内。

表1 60个谷子品种对10个谷瘟病菌株抗感反应Table 1 Sensitive reactions of 60 foxtail millet varieties to 10 millet blast isolates

续表1 Continued table 1
60个品种分别接种10个谷瘟病菌株,每个处理设3次重复,共计30个小区,随机区组排列,播种后10—15 d长出1—2片叶时进行间苗,每个营养钵定苗15株。
1.2 供试菌株的准备
10个谷瘟病菌菌株由中国农业科学院作物科学研究所在各地采集(图1),由黑龙江省农业科学院植物保护研究所分离,并接种高粱粒,经10—15 d,将长满菌丝体的高粱粒用无菌水洗下菌丝,滤干后,平摊在铺有1—2层湿纸的搪瓷盘内,上盖1—2层纱布,置于17—25℃室温下4—5 d,使其生长孢子,制成孢子悬浮液,在显微镜10×10倍的视野下观察,每个视野的孢子数量平均为20—30个,进行接种。每个菌株对60个供试品种均进行接种。
1.3 接种
于苗龄4—5叶期用喷雾器喷雾接种,接种后用遮阳网及塑料薄膜进行全面遮盖,保持高温、高湿条件12—15 h,促进发病。
1.4 病情鉴定和分级标准
于苗龄6叶期调查发病情况。病斑反应型是进行谷瘟病定级的主要依据。用目测法调查记载不同品种叶片上的病斑反应型,谷瘟病分级标准按照谷子谷瘟病0—9级法划分标准进行划分[13],免疫材料(0级)记为IM;高抗材料(1级)记为HR;抗病(3级)记为R;中抗(5级)记为MR;感病(7级)记为S;高感(9级)记为HS(图2)。
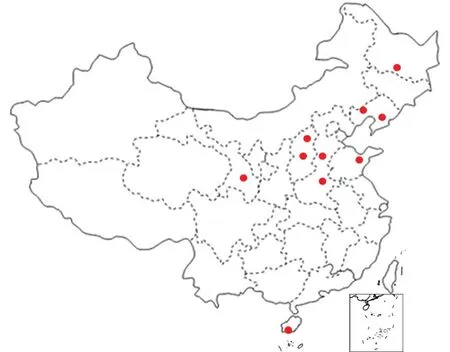
图1 10个谷瘟病菌在谷子主产区的分布Fig. 1 Distribution of 10 blast isolates in foxtail millet planting area
1.5 抗性频率、毒性频率和侵染率计算
参照水稻中稻瘟病抗性频率和毒性频率的计算方法,抗性频率 =(对供试品种表现抗病的菌株数/测试总菌株数)×100%[14];毒性频率 =(对供试品种表现感病菌株数/测试总菌株数)×100%[15]。某一菌株对品种的侵染率 = 能够侵染的品种数量/品种总数量×100%。
1.6 聚类分析
分别将供试的10个菌株与60份谷子品种的抗感性转换成0、1数据,用NTSYS软件进行系统聚类分析。

图2 不同品种谷子对谷瘟病菌的病斑反应类型Fig. 2 Reaction type of millet blast for different varieties of foxtail millet
1.7 鉴别品种选择原则
参照水稻鉴别品种的选择标准[16],遵循以下几个原则:(1)在3次重复中对菌株的抗感性表现明确并且相对稳定;(2)能够较容易的将不同的生理小种区分,即一个品种可以区分多个小种;(3)在生产和育种中有较高的应用价值;(4)尽量选择国内公知公用的品种。
2 结果
2.1 谷子品种对10个菌株抗性结果
接种后7 d,调查60个品种对10个菌株抗感反应(表1),对10个菌株都表现抗病的品种仅有单皮粘1个品种,黄棒头、齐头黄和白谷对多数菌株表现抗病,仅对其中1到2个菌株表现中抗,对谷瘟病抗病性表现明显;维子那谷和郑谷4号虽然对10个菌株均抗病,但是对4—6个菌株表现为中抗,抗病反应不是十分明确;对10个菌株均表现感病或高感的品种多达11个;其他品种则抗1—9个菌株不等,表现出不同品种谷子对谷瘟病菌株抗性的多样性。
2.2 鉴别品种的筛选及生理小种命名
2.2.1 鉴别品种的筛选 结合60个品种对10个谷瘟病菌的抗性频率(表2)和聚类分析(图3)结果,将参试品种分为高抗品种群、中抗品种群、中感品种群和高感品种群,从中选择鉴别品种。
高抗品种群:包括黄棒头、齐头黄、单皮粘、白谷、维子那谷、郑谷4号、新农691、嫩选十二号、广8、毛大粒、承农2号、红苗老来变、红谷、九谷14等14个品种,抗性频率为80%—100%,它们对大多数参试菌株表现高抗、抗病或中抗。其中单皮粘对10个菌株均表现抗病或高抗,抗性反应更明确、稳定,因此,选择能够代表东北谷子产区且抗性频率为100%的单皮粘作为鉴别品种。
中抗品种群:抗病频率为60%—70%,包括金屏谷子、黄苗谷子、SSR25、紫力根、锦谷5号、金德、三红苗等7个品种。其中金屏谷子、锦谷5号和金德对10个菌株抗病反应比较明确,仅对2—3个菌株抗病反应表现为中抗,其他品种对10个菌株抗病反应中多数表现为中抗,抗病反应不明确,不予选择,金德来源于美国,因此不予选择,最终选择能够代表夏谷种植区域抗性频率为70%的金屏谷子和抗性频率为60%的育成品种锦谷5号作为鉴别品种。
中感品种群:抗性频率为30%—50%,包括西1-2-4、大毛毛谷、公谷5号、黄大粒、茭谷、中谷1、黄谷、二白谷、龙辐60533、鲁谷9号、龙爪、晋谷20、安06h-8023、2015、晚猫爪粘、龙爪谷子等16个品种。大毛毛谷、中谷1、二白谷和安06h-8023抗病反应表现均比较明确,二白谷是新疆农家品种,不是谷子主产区,不予选择,大毛毛谷的抗性频率为50%,中谷1和安06h-8023都对GW8-2、GW10和GW2013有抗性,不具代表性,且中谷1和安06h-8023在种质资源库中无统一编号,因此,选择大毛毛谷作为鉴别品种。
高感品种群:抗性频率0—20%,有23个品种,占参试品种的38.33%。其中,龙爪谷和假金苗抗性频率均为20%,抗感反应十分明确,因此,选择甘肃镇原县的龙爪谷和内蒙古的假金苗作为鉴别品种。在高感品种中,牛头谷对10个菌株均表现高感,因此选择牛头谷作为是否接种上谷瘟病菌的感病指示品种。
根据4个品种群的分析,初步筛选出单皮粘、金屏谷子、锦谷5号、大毛毛谷、龙爪谷、假金苗和牛头谷等7个品种作为鉴别品种。

表2 参试品种的抗性频率Table 2 The resistance frequency of foxtail millet varieties tested
2.2.2 谷瘟病生理小种的分类和命名 选择的7个鉴别品种根据抗病程度,分别用英文字母A、B、C、D、E、F、G代表,并作为谷瘟病类群划分标识,进行谷瘟病生理小种分类时,在A品种单皮粘上呈感病反应的谷瘟病小种均属A群;在A品种表现抗病而在B品种金屏谷子上呈感病反应的谷瘟病小种均属B群;在A、B品种上表现抗病而在C品种锦谷5号上呈感病反应的谷瘟病小种均属C群;依次类推,分别划分D、E、F、G群,若遇到对全部鉴别品种均显示抗病反应时,另列为H群。按照GILMOUR[17]的八进位法进行谷瘟病生理小种命名,先将鉴别品种分为3组,每个鉴别品种编上固定的数码,第一组的单皮粘、金屏谷子、锦谷5号分别为1、2和4;第二组的大毛毛谷、龙爪谷、假金苗依次为10、20和40;第三组的牛头谷为100。被测的生理小种编号就是能够被菌株侵染的品种数字之和[18]。例如在1—7鉴别品种上的反应为RRRRSSS的菌株,在A、B、C和D品种上表现抗病,在E品种上表现感病,属于E群小种,在后面的其他鉴别品种上都表现为感病,则小种号是E、F和G这3个品种固定数码之和:20+40+100=160,为避免与第一套谷子鉴别体系生理小种命名混淆,用“中国”的“中”的汉语拼音zhong的第一个字母写在首位,Z字以后写上所属类群,小种编号在右下侧表示,因此,该生理小种记作ZE160。10个菌株根据命名规则,分别被分成8个生理小种(表3)。
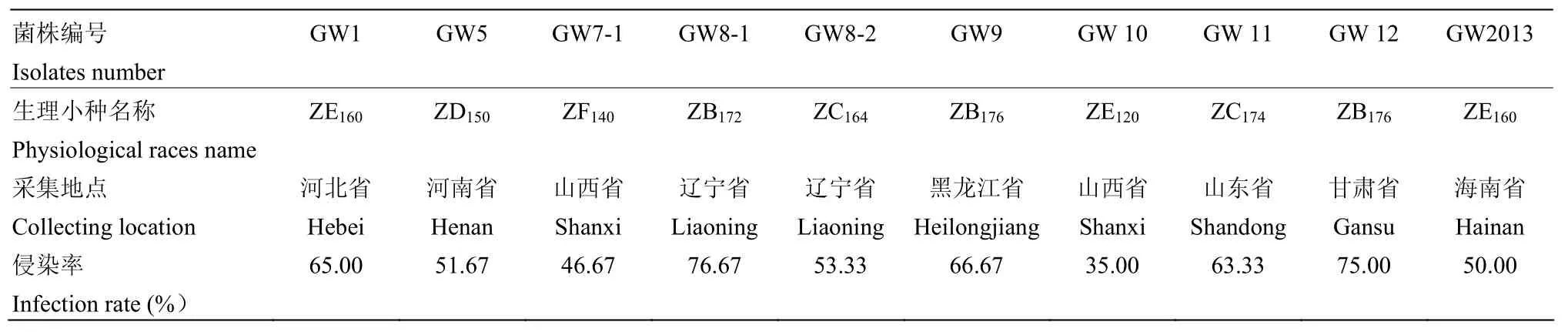
表3 8个谷瘟病菌生理小种侵染率Table 3 The infection rate of 8 millet blast physiological races
2.3 谷子品种的抗病性分析
根据60个品种对10个谷瘟病菌株抗感性不同,对10个谷瘟病菌株的抗性频率进行统计分析(表2)供试品种的抗性存在明显差异,其中,抗性频率100%的品种有6个,包括黄棒头、齐头黄、单皮粘、白谷、维子那谷、郑谷4号等,占参试品种的10.00%;抗性频率为0的品种有11个,占参试品种的18.33%;抗性频率低于30.00%的品种占到了53.33%,对大多数谷瘟病菌感病,抗谱较窄;其他43个品种对不同的谷瘟病生理小种表现了不同反应。
将供试品种对10个谷瘟病菌株的抗感反应数据用NTSYS软件进行聚类分析,以相似系数0.65为界,将60个品种分为五组(图3):第一组有13个品种,这一组的抗性频率从80%到100%,高抗谷瘟病;第二组有8个品种,抗性频率从50%到80%,中抗谷瘟病;第三组包括5个品种,抗性频率从40%到60%,中感谷瘟病;第四组仅包含4个品种,抗性频率从20%到40%,中感谷瘟病;第五组包含30个品种,抗性频率从0到30%,其中牛头谷、饿死牛、陇谷3号等11个品种对8个生理小种都表现感病,高感谷瘟病。
2.4 生理小种的致病性分析
对8个生理小种的侵染率进行统计(表3),结果表明,不同地理来源的生理小种侵染率存在较大差异。大多数生理小种的侵染率较高,致病能力较强,其中来自辽宁水保所试验基地的谷瘟病菌致病能力最强,侵染率达76.67%,只有在山西采集的2个谷瘟病菌株致病能力相对较弱,侵染率均低于50%。这说明不同菌株在侵染能力和致病性方面表现了较丰富遗传多样性。
通过分析10个谷瘟病菌株对60个谷子核心种质资源的致病性,用NTSYS软件对结果进行聚类分析,以相似系数0.70为界,10个谷瘟病菌株被分为Ⅲ类(图4),第Ⅰ类侵染率46.67%—76.67%,包含6个生理小种:ZE160、ZD150、ZF140、ZB172、ZB174和ZB176;第Ⅱ类包括2个生理小种:ZE120和ZE160;侵染率分别为35.00%和50.00%;第Ⅲ和只包括一个生理小种ZC164,侵染率为53.33%。10个谷瘟病菌株被分为B、C、D、E和F等5群8个生理小种,其中B群、C群和E群各有2个生理小种;D群和F群各有1个生理小种。谷瘟病菌株侵染率均较高,如黑龙江、山东、辽宁、甘肃等谷子主产区的生理小种侵染率均超过了60%,不同种群的生理小种分布不存在地理偏向性。
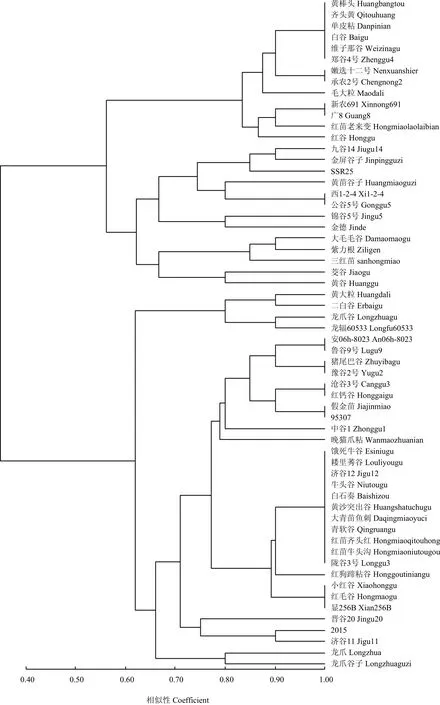
图3 60个品种对10个谷瘟病菌株抗感性构建的聚类图Fig. 3 The cluster of the 60 varieties based on the resistance to 10 millet blast isolates

图4 10个谷瘟病菌株之间的致病性关系Fig. 4 The relationship of pathogenicity among 10 millet blast isolates
3 讨论
谷瘟病对谷子生产造成巨大危害,其生理小种由于环境和寄主的变化不断发生变异,导致寄主抗性逐渐减退甚至丧失。因此,从谷子核心种质资源库筛选抗病品种,建立谷瘟病鉴别品种体系,明确谷子主产区谷瘟病菌的田间生理小种组成以及优势生理小种的分布和状态,监测推广品种的抗性,根据品种对不同生理小种的抗性,合理种植对当地优势生理小种具有相应抗性的品种[7],对防范谷瘟病严重发生具有重要意义。
3.1 广谱抗性材料的筛选
对核心种质资源进行抗谷瘟病鉴定,筛选广谱抗谷瘟病材料,是挖掘谷子中蕴含的抗谷瘟病基因的有效方法。本研究采用的是温室苗期鉴定法,从60份核心种质资源中筛选出6份广谱抗性材料,这些宝贵的抗谷瘟病资源可以作为杂交亲本用于抗病育种;也可以用于谷瘟病抗性基因的标记和克隆等基础研究。抗性频率越高,品种携带多个谷瘟病抗性基因的几率越大,品种对谷瘟病生理小种的抗谱越广,品种的抗病性越好,在谷子生产中应用价值越大。本研究中,抗性频率大于50%的有24个品种,占参试品种的40%,它们可以作为抗性资源,用于改良谷子品种的谷瘟病抗性;抗性频率较低的品种抗谱较窄,不宜在谷子主产区大规模种植,育种选作亲本也应当慎重。在60份核心种质资源中,沧谷3号和红钙谷、假金苗和95307、齐头黄和单皮粘等品种对10个谷瘟病菌的抗感反应表现完全相同,表明它们对这些谷瘟病菌株的抗性基因可能是完全相同的,或者说是等效的。齐头黄和单皮粘分别是江苏和吉林的农家品种,地理环境相差巨大,但对谷瘟病的抗性则表现一致,说明谷子对谷瘟病的抗性和地理分布没有直接关系,而跟品种本身的基因组成有关,因此,今后可以通过挖掘国内外不同地理来源的谷子品种蕴含的广谱抗谷瘟病基因,为农业生产服务。
谷子品种资源多数对谷瘟病菌生理小种抗谱不广,谷瘟病生理小种分布规律尚不明确,需要在挖掘现有抗谷瘟病资源基础上,利用远缘杂交等手段,寻找更多的抗源。水稻种质资源蕴涵丰富的抗稻瘟病资源[19-20],杨健源等[21]研究发现,在水稻三黄占2号EMS诱变突变体库中包含多种稻瘟病抗性类型,因此,也可利用谷子EMS突变体库,挖掘新的抗谷瘟病资源,为抗谷瘟病基因的克隆和育种应用奠定物质基础。
3.2 谷瘟病生理小种鉴定的标准体系建立
1974年,曹功懋等[22]首先报道了粟瘟病菌存在致病性分化,阎万元等[5]用吉林省地区的18个单孢分离菌,对246个谷子品种进行测定,最后选定了白沙粘310、衡研130、鲁谷2号、新农761、罗谷6号、金镶玉等6个品种作为鉴别品种,将来自北方10个省(区)的711株单孢分离菌划分为7群32个生理小种,建立了第一套鉴别谷瘟病菌生理小种的鉴别品种,明确了中国北方谷子产区谷瘟病的优势小种,在当时起到非常重要的作用。但是,随着谷瘟病菌致病性的演化[23-24],目前,一套鉴定品种已经很难适应中国不同省(市,地区)鉴别谷瘟病菌生理小种的需要[5],且这套谷子品种材料因多年无人应用和管理,确切基因型和种子来源已不清楚。本研究利用谷子主产区的10个谷瘟病单胞菌株,对60份谷子核心种质资源接种鉴定,从谷子核心种质资源中初步筛选出7份谷子品种作为鉴别品种,建立了第二套谷瘟菌小种鉴别标准品种。但限于本研究涉及的谷瘟病采样数目,本研究所建立的小种鉴定标准品种体系也需进一步核实验证,在应用中逐步完善。
在抗病育种中,品种所携带的基因数目越多越好,而鉴别标准品种则需要为单基因控制[25],谷子的两套鉴别品种均未实现抗病基因的克隆,仅仅是按照菌株分类选择的品种,随着谷子测序的完成以及谷子功能基因组研究平台的建立,利用抗性频率低的谷子品种携带抗谷瘟病单基因可能性较大的特点,通过与普感品种杂交,可快速获得携带单抗基因的材料,并应用于今后的近等基因系构建及抗病基因的定位与克隆研究。
4 结论
选用60份谷子核心种质资源,从中选出单皮粘、金屏谷子、锦谷5号、大毛毛谷、龙爪谷、假金苗和牛头谷7个品种作为鉴别品种,建立了第二套谷瘟病生理小种鉴别标准品种。将来自包括河北、河南、山西、内蒙古、黑龙江等谷子主产区采集分离的10份谷瘟病菌单孢分成8个生理小种,筛选出黄棒头、齐头黄、单皮粘、白谷、维子那谷和郑谷4号等6个抗广谱菌株的基础材料,作为谷子抗谷瘟病育种的抗病基因来源。
[1] MURAKAMI J, TOMITA R, KATAOKA T, NAKAYASHIKI H, TOSA Y, MAYAMA S. Analysis of host species specificity of Magnaporthe grisea toward foxtail millet using a genetic cross between isolates from wheat and foxtail millet. Phytopathology, 2003, 93(1): 42-45.
[2] 刘辉, 林汝法, 徐婀娜, 杨守文, 李志良. 粟种质资源抗粟瘟病鉴定. 陕西农业科学, 1990(3): 24-25. LIU H, LIN R F, XU E N, YANG S W, LI Z L. Screening germplasm resoures of foxtail millet for millet blast resistance. Shaanxi Journal of Agricultural Sciences, 1990(3): 24-25. (in Chinese )
[3] NISHIKADO Y. Studies on the rice blast diseases. Japanese Journal of Botany, 1927(3): 239-244.
[4] 梁平彦, 李玉麟, 沈丽明. 粟瘟病的研究. 植物病理学报. 1959, 5(2): 89-99. LIANG P Y, LI Y L, SHEN L M. Studies on millet blast caused by Piricularia setariae Nishikado. Acta Phyto-Pathologica Sinica, 1959, 5(2): 89-99. (in Chinese)
[5] 阎万元, 谢淑仪, 金莲香, 刘洪江, 胡吉成. 粟瘟病菌生理小种研究初报. 中国农业科学, 1985(3): 57-62. YAN W Y, XIE S Y, JIN L X, LIU H J, HU J C. A preliminary study on the physiological races of millet blast(Pyricularia setariae Nishikado). Scientia Agricultura Sinica, 1985(3): 57-62. (in Chinese)
[6] 阎万元, 谢淑仪, 金莲香, 刘洪江, 李乃明, 胡吉成. 吉林省粟瘟病菌生理小种研究. 吉林农业科学, 1986(3): 49-54. YAN W Y, XIE S Y, JIN L X, LIU H J, LI N M, HU J C. Study on the physiological races of millet blast in jilin province. Jilin Agricultural Sciences, 1986(3): 49-54. (in Chinese)
[7] 阎万元, 谢淑仪, 刘洪江, 李乃明. 吉林省谷子品种资源及生产品种对粟瘟病专化抗性鉴定. 吉林农业科学, 1988(1): 6-10. YAN W Y, XIE S Y, LIU H J, LI N M. Evaluation for the specialized resistance of millet germplasm resources and cultivars to millet blast in jilin province. Jilin Agricultural Sciences, 1988(1): 6-10. (in Chinese)
[8] 汤智鹏. 59份水稻种质资源对稻瘟病的抗性及农艺性状评价[D].雅安: 四川农业大学, 2011. TANG Z P. Evaluation of resistance to rice blast and agronomic traits to 59 rice germplasm resources[D]. Yaan: Sichuan Agricultural University, 2011. (in Chinese )
[9] 王雅儒, 褚菊征, 宋燕春, 谢淑仪, 阎万元, 金莲香, 刘辉, 谢生瑞.谷子品种抗粟瘟病的鉴定研究. 植物保护学报, 1985, 12(3): 175-180. WANG Y R, CHU J Z, SONG Y C, XIE S Y, YAN W Y, JIN L X, LIU H, XIE S R. Screening tests of millet varieties for resistance to blast pathogen. Acta Phytophylacica Sinica, 1985, 12(3): 175-180. (in Chinese )
[10] 吴秀兰. 谷子谷瘟病鉴定与筛选初报. 黑龙江农业科学, 1985(1): 46-50. WU X L. A preliminary study on the identification and screening of millet blast. Heilongjiang Agricultural Sciences, 1985(1): 46-50. (in Chinese)
[11] 张秀雯. 河南省谷子品种资源抗谷瘟病鉴定初报. 河南农业科学, 1987(7): 12-14. ZHANG X W. A preliminary identification of foxtail millet germplasm to the millet blast in Henan province. Henan Agricultural Sciences, 1987(7): 12-14. (in Chinese)
[12] JIA G Q, HUANG X H, ZHI H, ZHAO Y, ZHAO Q, LI W J, CHAI Y, YANG L F, LIU K Y, LU H Y, ZHU C R, LU Y Q, ZHOU C C, FAN D L, WENG Q J, GUO Y L, HUANG T, ZHANG L, LU T T, FENG Q, HAO H F, LIU H K, LU P, ZHANG N, LI Y H, GUO E H, WANG S J, WANG S Y, LIU J R, ZHANG W F, CHEN G Q, ZHANG B J, LI W, WANG Y F, LI H Q, ZHAO B H, LI J Y, DIAO X M, HAN B. A haplotype map of genomic variations and genome-wide association studies of agronomic traits in foxtail millet (Setaria italica). Nature Genetics, 2013, 45(8): 957-961.
[13] 陆平. 谷子种质资源描述规范和数据标准. 北京: 中国农业出版社, 2006: 18.LU P. Descriptors and Data Standard for Foxtail Millet[Setaria italica(L. ) Beuauv. ]. Beijing: China Agricultural Press, 2006: 18. (in Chinese)
[14] 刘文德, 易运萍, 鲁国东, 王宗华. 福建省近年来主栽水稻品种抗瘟性评价. 西南农业学报, 2008, 21(4): 993-995. LIU W D, YI Y P, LU G D, WANG Z H. Blast resistance evaluation for major rice varieties currently deployed in fujian province. Southwest China Journal of Agricultural Sciences, 2008, 21(4): 993-995. (in Chinese)
[15] 鲁国东, 郑武, 阮志平, 陈在杰, 王宗华. 福建稻瘟菌毒性类型组成及其对水稻几个Pi基因的毒性频率. 植物病理学报, 2003, 33(3): 248-253. LU G D, ZHENG W, RUAN Z P, CHEN Z J, WANG Z H. Pathotypes of Magnaporthe grisea Barr. and their virulence to several rice Pi-genes in Fujian province. Acta Phytophylacica Sinica, 2003, 33(3): 248-253. (in Chinese)
[16] 全国稻瘟病菌生理小种联合试验组. 我国稻瘟病菌生理小种研究.植物病理学报, 1980, 10(2): 71-82. All China Corporation of Research on Physiological Races of Pyricularia oryzae. Research on physiological races of rice blast fungus in China. Acta Phytophylacica Sinica, 1980, 10(2): 71-82. (in Chinese )
[17] GILMOUR J. Octal notation for designating physiologic races of plant pathogens. Nature, 1973, 242: 620.
[18] 凌忠专, 雷财林, 王久林. 稻瘟病菌生理小种研究的回顾与展望.中国农业科学, 2004, 37(12): 1849-1859. LING Z Z, LEI C L, WANG J L. Review and prospect for study of physiologic races on rice blast fungus(Pyricularia grisea). Scientia Agricultura Sinica, 2004, 37(12): 1849-1859. (in Chinese )
[19] 沈瑛, ADREIT H, 朱旭东, MILAZZO J, 陈红旗, THARREAU D.中国部分杂交稻和常规早籼、晚粳品种(系)的抗瘟性. 中国农业科学, 2004, 37(3): 362-369. SHEN Y, ADREIT H, ZHU X D, MILAZZO J, CHEN H Q, THARREAU D. Evaluation of resistance of some hybrid rices, conventional early Indica and late Japonica rice to Magnaporthe grisea in China. Scientia Agricultura Sinica, 2004, 37(3): 362-369. (in Chinese )
[20] 黄富, 刘成元, 谢戎, 叶华智. 水稻种质资源抗稻瘟病及其恢保特性评价. 中国农业科学, 2005, 38(4): 831-836. HUANG F, LIU C Y, XIE R, YE H Z. Evaluation of blast resistance and restoring or remaining characteristics of rice germplasm resources. Scientia Agricultura Sinica, 2005, 38(4): 831-836. (in Chinese )
[21] 杨健源, 张少红, 姜先芽, 黄显良, 黄章慧, 曾列先, 李传瑛, HEI LEUNG, 朱小源, 刘斌. 三黄占2号EMS诱变突变体库的构建与稻瘟病抗性初步评价. 广东农业科学, 2011, 16: 1-3. YANG J Y, ZHANG S H, JIANG X Y, HUANG X L, HUANG Z H, ZENG L X, LI C Y, HEI L, ZHU X Y, LIU B. Development of EMS-induced mutants of indica rice Sanhuangzhan-2 and their preliminary evaluation of blast resistance. Guangdong Agricultural Science, 2011, 16: 1-3. (in Chinese )
[22] 曹功懋, 阎万元. 粟瘟病防治研究. 吉林农业科学, 1982(2): 53-58. CAO G M, YAN W Y. Studies on control of the millet blast. Jilin Agricultural Sciences, 1982(2): 53-58. (in Chinese )
[23] 卫丽, 王同朝, 张桂兰. 谷子品种资源营养品质分析及抗病性鉴定. 华北农学报, 1999, 14(22): 1-5. WEI L, WANG T C, ZHANG G L. A study on protein and fat contents and characterization of disease resistance in millet varieties. Acta Agriculturae Boreali-Sinica, 1999, 14(22): 1-5. (in Chinese )
[24] 王妍, 郑文静, 王辉, 张丽霞, 王世维, 赵家铭, 刘志恒. 辽宁省主栽水稻品种抗稻瘟病基因的鉴定及分析. 植物遗传资源学报, 2015, 16(3): 640-648. WANG Y, ZHENG W J, WANG H, ZHANG L X, WANG S W, ZHAO J M, LIU Z H. Identification and analysis of rice blast resistance gene alleles in 24 rice cultivars from Liaoning province. Journal of Plant Genetic Resources, 2015, 16(3): 640-648. (in Chinese )
[25] 凌忠专. 浅谈抗稻瘟病育种问题. 作物杂志, 1989, 3: 35-36. LING Z Z. The problem of rice breeding resistant to rice blast. Crops, 1989, 3: 35-36. (in Chinese )
(责任编辑 李莉)
Determination of Standard Varieties for Identifying Physiological Races of Foxtail Millet Blast Fungus
LI Zhi-jiang1, JIA Guan-qing2, LI Xiang-yu1, LI Yi-chu3, MA Jin-feng1, ZHI Hui2, TANG Sha2, ZHANG Shuo2, CHAI Yang2, LI Yan-dong1, DIAO Xian-min2
(1Institute of Crop Breeding, Heilongjiang Academy of Agricultural Sciences, Harbin 150086;2Institute of Crop Sciences, Chinese Academy of Agricultural Sciences, Beijing 100081;3Institute of Plant Protection, Heilongjiang Academy of Agricultural Sciences, Harbin 150086)
【Objective】Foxtail millet blast (Pyricularia setariae Nisikado) is an important disease, which could cause reduced production and quality of foxtail millet, and the identification of different millet blast physiological races are very important for evaluating the resistant resources, breeding of resistant varieties and reasonable allocation of different resistant varieties for cultivation. The goal of this study is to determine standard varieties for the identification of physiological races of millet blast fungus, which would establish a basis for the evaluation of resistance of foxtail millet to different blast races. 【Method】 Sixty foxtail millet varieties collected from different geographical regions in China were inoculated using ten foxtail millet blast isolates sampled fromdifferent eco-regions. Statistical results of the sensitive reaction of 60 foxtail millet varieties to 10 blast isolates were recorded into excel format according to 0-9 grade standards of millet blast. The different resistant results were recorded as high resistance, resistance, middle resistance, sensitivity and high sensitivity. The high resistance, resistance, middle resistance were denoted with 0, and the sensitivity and high sensitivity were denoted with 1. The results were changed to 0/1 to construct the phylogenetic tree by NTSYS. Based on the resistant and infected results of sixty varieties to ten blast isolates, the standard identification system of foxtail millet blast were constructed. 【Result】Clustering analysis based on the pathogenicity of ten blast isolates and resistance of sixty varieties was conducted, respectively. The foxtail millet blast isolates were grouped into three clusters by similarity coefficient 0.70 as boundary, and foxtail millet varieties were divided into five categories by similarity coefficient 0.65 as boundary. Seven varieties, including Danpinian, Jinpingguzi, Jingu5, Damaomaogu, Longzhuagu, Jiajinmiao and Niutougu, were applied as standard identification varieties. The resistant varieties, Huangbangtou, Qitouhuang, Danpinian, Baigu, Weizinagu, and Zhenggu4 with broad spectrum resistance were screened, which could be used as the resources of resistant gene in foxtail millet blast disease breeding.【Conclusion】This study screened a set of standard varieties for foxtail millet blast fungus identification, and identified several genetic stocks with broad-spectrum resistance to foxtail millet breeding.
foxtail millet; Pyricularia setariae; core germplasm; physiological race; disease resistance
2016-04-11;接受日期:2016-06-06
国家现代农业产业技术体系(CARS-07-12.5-B15)
联系方式:李志江,Tel:0451-86668742;传真:0451-86667448;E-mail:lizhijiang12@163.com。通信作者刁现民,Tel:010-62126889;E-mail:diaoxianmin@caas.cn
猜你喜欢
电脑迷(2022年7期)2022-07-27
今日农业(2022年4期)2022-06-01
南方农业学报(2022年11期)2022-03-18
农业科技与信息(2021年4期)2021-12-05
生物灾害科学(2020年4期)2020-12-24
植物保护(2017年4期)2017-08-09
浙江农业学报(2017年1期)2017-05-17
中国马铃薯(2017年1期)2017-03-02
新农业(2016年20期)2016-08-16
中国茶叶(2016年4期)2016-01-18
