
氮素形态对烟苗根系生长及氮素利用的影响
2016-11-16 09:17邢瑶马兴华
中国烟草学报 2016年4期
邢瑶,马兴华
中国农业科学院烟草研究所,农业部烟草生物学与加工重点实验室,山东 青岛 266101
氮素形态对烟苗根系生长及氮素利用的影响
邢瑶,马兴华
中国农业科学院烟草研究所,农业部烟草生物学与加工重点实验室,山东 青岛 266101
为探讨不同形态氮素对烟草根系生长和氮素利用的影响,以烤烟品种NC55为材料,在水培条件下,设置相同氮浓度不同铵硝配比的5个处理,研究了不同氮素形态营养下根系形态生理特征及各器官氮素积累分配的差异,结果表明:处理后25d,纯铵态氮处理的烟苗地上部干重和总干重,总根长、根系总表面积、总体积、可溶性糖含量及茎和叶的含氮量、氮积累量与氮分配率均最低;纯硝态氮处理的烟苗根系干重、平均直径与活力及根系含氮量与氮积累量最低。处理20d内,铵硝比50:50的处理根系干重、氮积累量和植株总干重最大。处理后25d,铵硝比70:30的处理根系和地上部干重,根系总体积、总表面积、活力与可溶性糖含量及根系含氮量与氮积累量最高;铵硝比30:70的处理叶片含氮量与氮积累量最高。在2 mmol/L的氮浓度下,单施铵态氮或硝态氮抑制了烟苗生长及对氮素的利用,混施铵态氮和硝态氮促进烟苗的生长发育及对氮素的利用,其中铵硝比70:30促进烟苗根系与地上部的协调生长,铵硝比30:70促进烟苗地上部对氮素的吸收利用。
烟苗;氮素形态;根系;氮素利用
氮肥影响作物的产量和品质,调控氮肥的施用可达到增产稳产的目的[1-2]。当前多以向耕作土壤中施入氮肥,迅速提高无机态氮的含量来促进作物的生长,但如果作物和土壤微生物无法充分利用,易引起土壤氮的损失,最终导致严重的环境问题[3-4]。如何合理施用氮肥,提高作物对氮素的利用是兼顾产量和环境问题的关键。目前,利用营养元素调节根的形态和功能成为提高氮素利用率的重要途径[5]。
根系是植株吸收水分和养分的主要器官,在矿质营养中,氮素对其形态和生长的影响最大,同时根系的大小和形态学差异影响养分的吸收效率[6-7]。氮素供应充足时,植株根系较大,能充分利用深层土壤中的氮素,减少损失,有利于获得较高的产量[8-9]。虽然适量增施氮肥能够促进氮素的吸收[10],但也会增加土壤对氮素的固定和氮肥的损失[11]。因此通过氮肥调控根系生长,提高其对氮素的吸收利用是降低氮肥投入、减少损失、提高效率的重要途径。铵态氮和硝态氮是植物根系吸收无机态氮的主要形式。研究表明,不同氮素形态的氮源对根系长度、总表面积、根尖数和根系活力等形态、生理指标的影响不同,如铵态氮促使植株根系变短、侧根减少,硝态氮则会改变主根或侧根的长度[12-14]。不同形态氮肥会影响根系生长发育,进而影响根系吸收养分和水分的能力以及对氮素的利用效率[15-16]。
烟草是我国重要的经济作物,生产上普遍存在根系小、分布浅、发育不完全的现象[17]。不同形态氮素对烟草根系生长的影响研究较少,且氮素形态对烟草根系形态特征及氮素吸收利用的作用机制尚不明确。因此,本试验采用水培法,研究烟苗生长、根系形态生理特征以及各器官氮素吸收、积累与分配对不同形态氮素的响应,分析烟苗根系生长和氮素利用与氮素形态的相互关系,为生产上合理配施氮肥提供依据。
1 材料与方法
1.1 试验材料
供试烤烟品种为NC55。
1.2 试验设计
将包衣种子播种于塑料培养盒中,基质为消毒的石英砂,培养液为完全营养液(N以NH4NO3形式加入)。待幼苗长至7叶1心期时,选择长势一致的烟苗,洗净根部砂粒,移栽于2L的塑料容器中(容器深约17cm),塑料容器上覆盖具孔盖板,每个容器15株,植株根茎结合部位用海绵包裹,固定在盖板上。先用1/4完全营养液(N以NH4NO3形式加入)培养3d后,使烟苗适应水培环境,再用1/4无氮营养液培养2d,然后用不同形态的氮素处理。所有烟苗放置于同一温室中,温室温度控制在25~28℃/18~20℃(光/暗),光照周期为12h/12h(光/暗)。完全营养液的成分 为:N 2.0 mmol/L、Mg 0.6 mmol/L、P 0.3 mmol/L、K 1.25 mmol/L、Ca 1.0 mmol/L、Cl 2.0 mmol/L、Fe(Fe-EDTA)15.00 µmol/L、B 10.00 µmol/L、Mn 1.00 µmol/L、Cu 0.10 µmol/L、Zn 0.50 µmol/L、Mo 0.05 µmol/L。设置5个处理,即其中N以NaNO3、(NH4)2SO4和NH4NO3的形式加入。每个处理30株,每次采集6株。每3d更换一次营养液,并用0.1mol/L的NaOH调节pH在6.8~7.0之间。烟苗自培养0d开始,每5d进行取样测定。
1.3 测定项目和方法
烟苗采样后,用蒸馏水冲洗植株,去除植株表面残留氮素。将样品按根、茎、叶分样,称量鲜量,并对根系进行扫描分析,后70℃烘干至恒重,称量干重,粉碎,过100目筛,用于测定总氮、可溶性糖含量。
根系扫描采用EPSON Perfection V700 Photo(分辨率为6400×9600 dpi),图像采用WinRHIZO Pro 2012b分析程序进行分析。
根系活力采用TTC还原法测定[18],根系可溶性糖含量采用蒽酮比色法测定[18]。氮含量采用碳氮硫元素分析仪(Elementar vario PYRO cube, Germany)测定。各器官的氮积累量=器官含氮量×器官干物质量,氮分配率=器官氮素积累量/根、茎、叶氮素积累量之和。
用Excel 2007软件进行数据整理和作图,用SAS 9.2数据分析软件进行统计分析,采用LSD法,差异有统计学意义的水平设定为α=0.05。
2 结果与分析
2.1 氮素形态对烟苗干物质积累的影响
由表1可见,随处理时间延长,各处理烟苗根系干重、地上部分干重以及植株总干重均呈增加的趋势。处理后5d,T3根系干重显著高于T1和T5,T3地上部干重和植株总干重显著高于T1、T4和T5;处理后10~20d,T3根系干重、地上部干重及总干重最高,T5根系干重最低,而T1的地上部及总干重最低;处理后25d,T2根系干重显著高于其他处理,且T2、T3和T4地上部干重及植株总干重显著高于T1和T5。处理周期内,T1的根冠比始终最高。
2.2 氮素形态对根系生长的影响
2.2.1 氮素形态对根系特征的影响
由表2可以看出,总根长、根系总表面积及总体积均随处理时间延长呈增加趋势。处理后5d、15d和20d,各处理根系平均直径差异没有统计学意义;处理后25d,T1和T2显著高于其他3个处理;处理后5d,T1和T4的总根长、根系总表面积和总体积均显著低于T2和T3。处理后10d,总根长、根系总表面积和总体积均以T3最高,T5最低;处理后15d,T3的根系总体积最大,T2的总根长和总表面积最大,均显著高于T1和T5;处理后20d,T3的总根长、根系总表面积、总体积均最高,T5均最低;处理后25d,T1的总根长和总表面积显著低于其他处理,但其平均直径较高,T2的根系总表面积、总体积及平均直径最大,但总根长以T3最高。
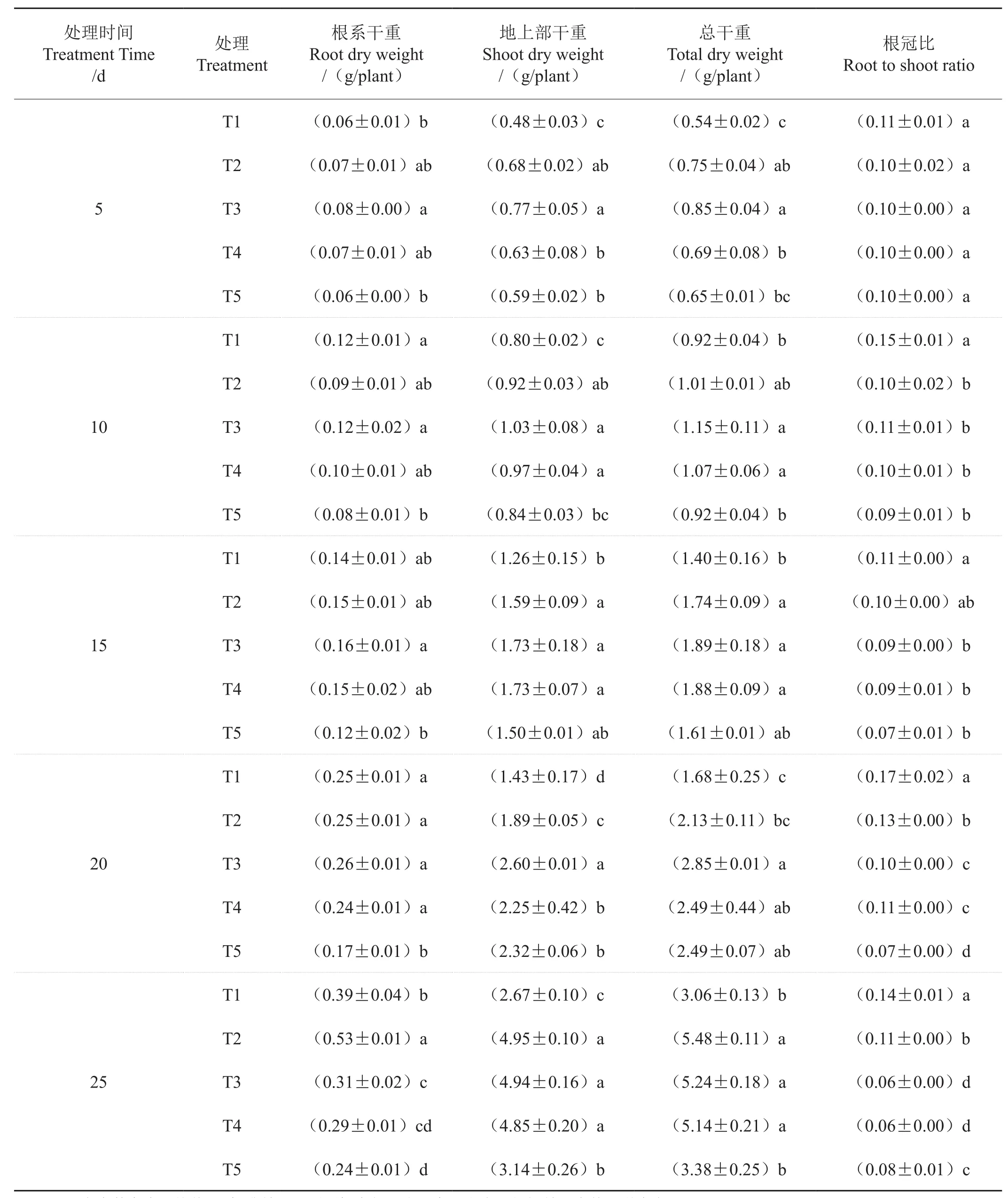
表1 氮素形态对烟苗干物质积累的影响Tab. 1 Effect of nitrogen form on dry matter accumulation of tobacco seedlings
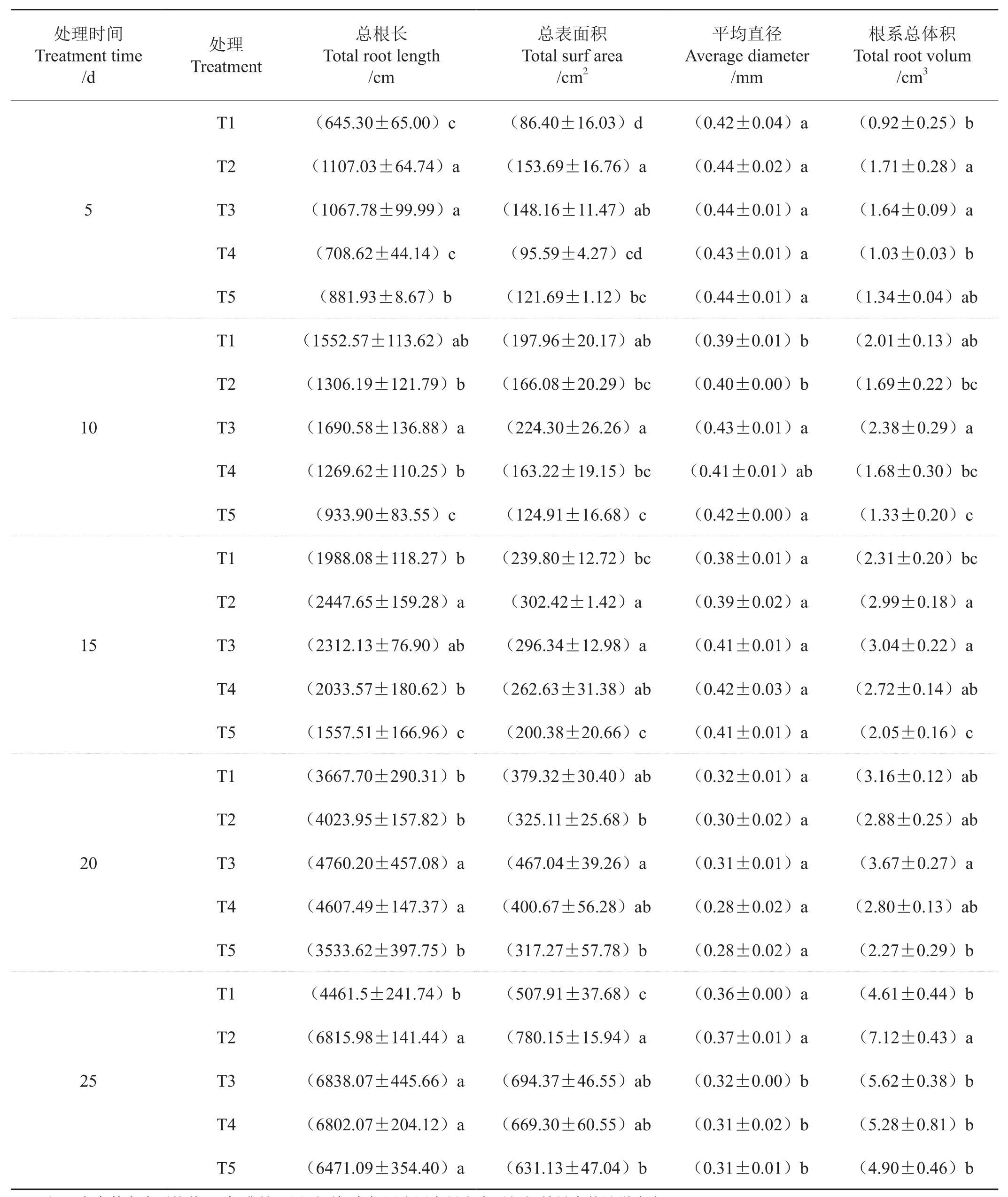
表2 氮素形态对根系形态的影响Tab. 2 Effect of nitrogen form on root morphology
2.2.2 氮素形态对根系生理指标的影响
由图1可以看出,处理后5d,T1根系活力最高,且处理间差异均有统计学意义;处理15d,T3根系活力显著高于其他处理,T1根系活力显著高于T2;处理后20d,T2根系活力最高,T1次之,T4最低;处理后25d,处理间根系活力差异均有统计学意义,表现为T2>T3>T4>T1>T5。随处理时间的延长,根系可溶性糖含量呈增加趋势。处理后5d,T4可溶性糖含量显著高于其他处理;处理后20d,T5根系可溶性含量最高,显著高于T1、T2和T3;处理后10d和25d,根系可溶性糖含量均以T2最高,T1最低,T3显著高于T4和T5。
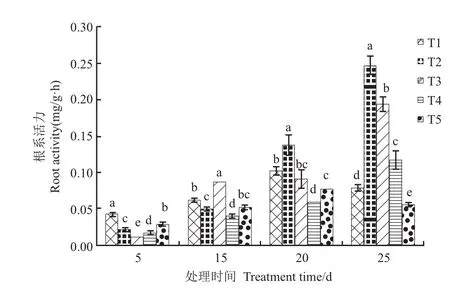
图1 氮素形态对根系活力的影响Fig. 1 Effect of nitrogen form on root activity
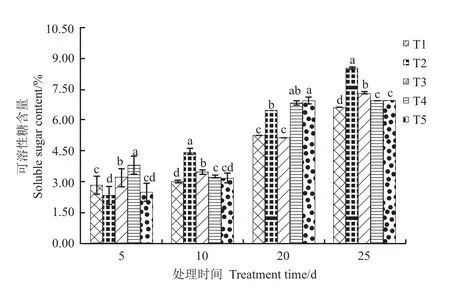
图2 氮素形态对根系可溶性糖含量的影响Fig. 2 Effect of nitrogen form on root soluble sugar content
2.3 氮素形态对烟苗氮素积累分配的影响
2.3.1 氮素形态对烟苗含氮量的影响
处理后5d,T2和T5根系含氮量显著高于T1和T4(表3);处理后10d,T3和T5根系含氮量显著高于T1;处理后15d,T1和T5显著高于T2、T3和T4;处理后20d,T3根系含氮量最高,显著高于T4和T5;处理后25d的含氮量T4和T5显著低于其他处理。处理后5d和10d,T1和T4茎含氮量显著低于其他处理;处理后15d和20d,T4茎含氮量显著高于其他处理;处理后25d,T5茎含氮量最高,T1和T2最低。叶片含氮量处理后5d,处理间差异没有统计学意义;处理后10d除T1外,其他处理间差异没有统计学意义;处理后15d,T4叶含氮量显著高于T1和T5;处理后20d,处理间叶含氮量差异有统计学意义,以T4最高;处理后25d,T3、T4和T5叶含氮量显著高于T1和T2。
2.3.2 氮素形态对烟苗氮素积累量的影响
由表3可见,各器官氮素积累量随处理时间延长而增加。处理后5d和10d,T3根系氮积累量最高;处理后15d,处理间差异没有统计学意义;处理后20d,T2和T3根系氮积累量显著高于T4和T5;处理后25d,T2根系氮积累量最高,显著高于T1和T3,且这3个处理都显著高于T4和T5。处理后5d,T2、T3和T4处理间茎氮积累量差异没有统计学意义,但都显著高于T1,T2和T3的叶片氮积累量显著高于T1和T4;处理后10d,T5茎氮积累量最高,显著高于T1,T3叶片氮积累量显著高于T1、T2和T5;处理后15d,T3、T4和T5之间茎氮素积累量没有统计学意义,但显著高于T1和T2,T3和T4叶片氮素积累量显著高于T1和T5;处理后20d,T5茎氮积累量显著高于T1、T2和T3,T3的叶氮积累量最高,且处理间差异均有统计学意义;处理后25d,除T3与T4之间叶氮积累量没有统计学意义外,各处理间茎和叶的氮积累量差异均有统计学意义,其中T3的茎氮素积累量最高,T4的叶氮素积累量最高。
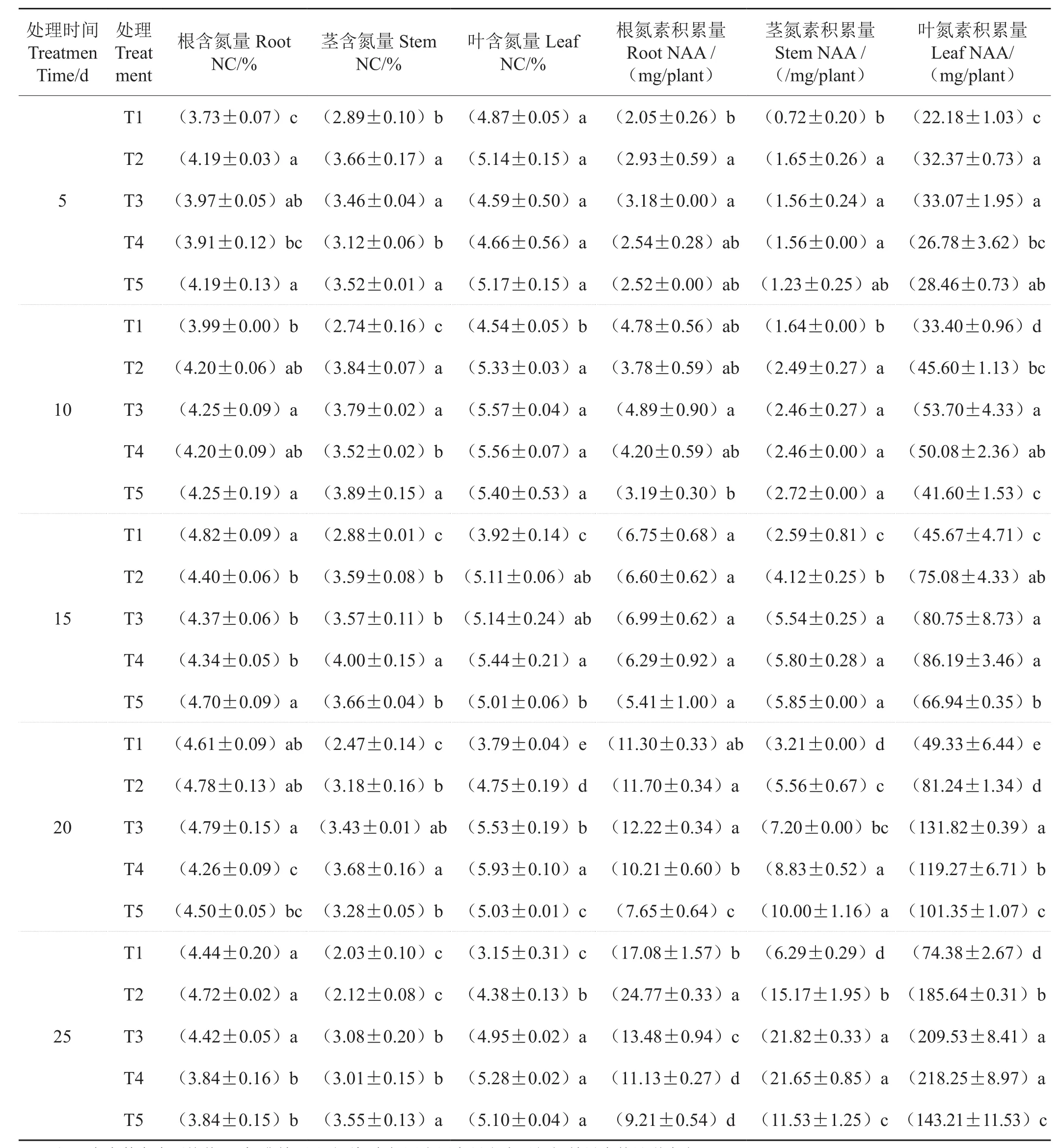
表3 氮素形态对烟苗氮素含量和积累量的影响Tab. 3 Effect of nitrogen form on nitrogen content and nitrogen accumulation amount of tobacco seedlings
2.3.3 氮素形态对烟苗氮素分配率的影响
从表4可见,处理后5d,各处理根系氮素分配率差异没有统计学意义;处理后10d和15d,除T1显著高于其他处理外,其他处理间差异没有统计学意义;处理后20d和25d,T1根系分配率显著高于T2;T3、T4和T5均显著低于T2。处理后5d,T4茎氮素分配率最高,且显著高于T1。处理后10d和20d,T5茎的氮素分配率显著高于其他处理,T3最低;处理后15d,T5茎的氮素分配率显著高于T1和T2;处理后25d,T3和T4的茎氮分配率显著高于其他处理。处理后5d,处理间叶片氮素分配率差异没有统计学意义;处理后10d,T1叶片氮素分配率显著低于其他处理;处理后15d,T4叶片氮分配率最高,显著高于T1和T5;处理后20d,T3、T4和T5叶片氮分配率显著高于T1和T2,且T2显著高于T1;处理后25d,随硝态氮比例增加,叶片氮分配率升高,除T4外,其他处理均达显著水平。
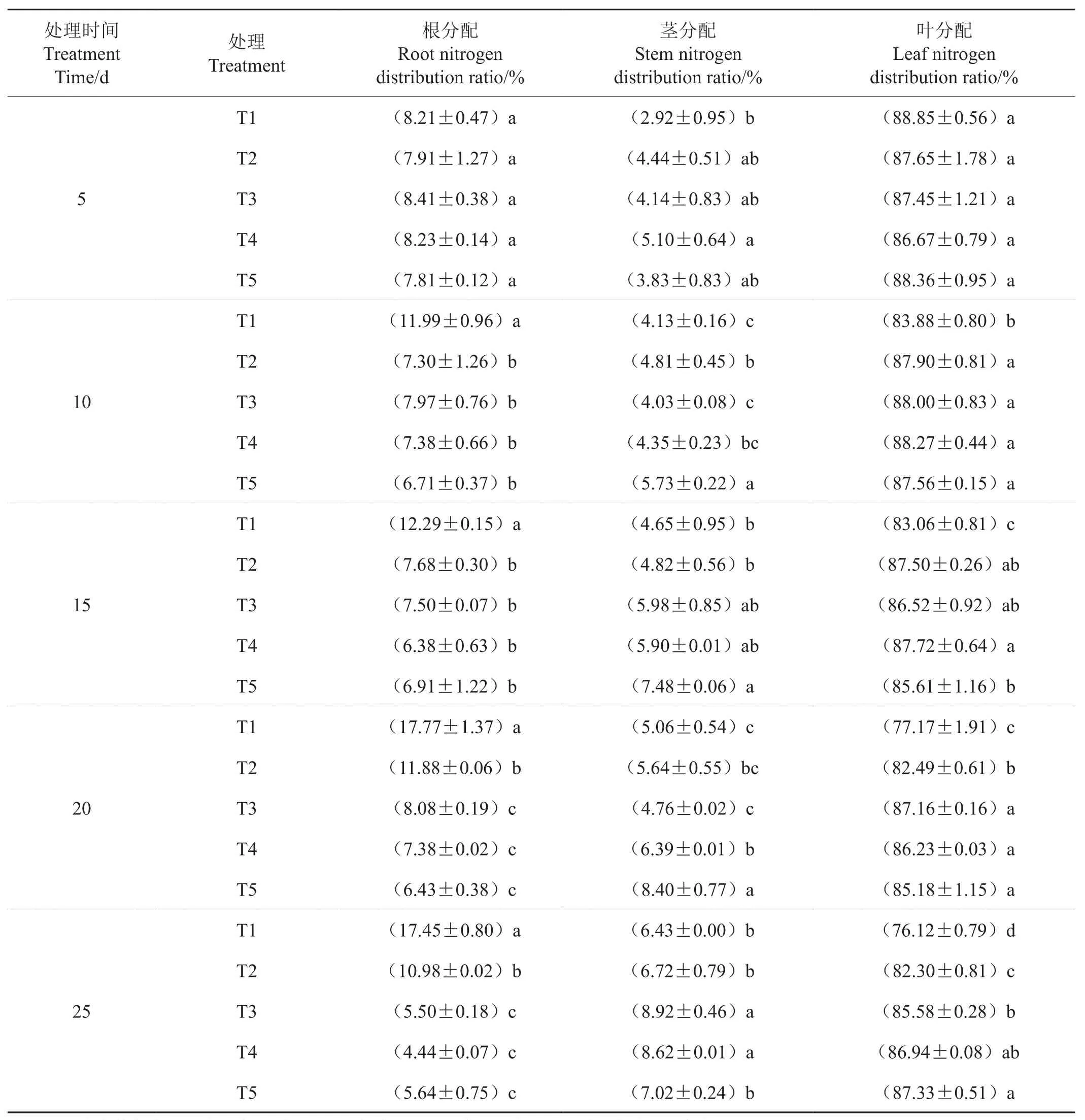
表4 氮素形态对烟苗氮素分配率的影响Tab. 4 Effect of nitrogen form on nitrogen distribution ratio of tobacco seedlings
3 讨论
3.1 不同氮素形态对根系的影响
近年来,许多学者从环境胁迫、氮素用量、氮素与其他环境因素耦合等方面对不同作物的根系形态、根系生理特性进行了深入的研究[19-21]。但关于氮素形态对根系形态、生理的影响方面研究较少。对大豆的研究表明,单一形态的氮素对大豆根系的生长都有抑制作用,而铵硝比1∶1的氮素营养促进植株的生长[22]。对紫花苜蓿的研究也表明,铵硝比1∶1的处理根系生物量、根表面积、体积、根系活力、根瘤数等指标最优,纯施用铵态氮处理次之,纯施用硝态氮的处理最低[23]。
本研究结果表明,在处理周期内,铵硝比为70∶30或50∶50的处理根系干重高于其他处理,单施硝态氮处理根系干重最低。说明硝态氮处理抑制了烟苗根系生长,铵硝配施有利于促进根系生长,且铵态氮比例略高作用更明显。从根系形态特征分析,虽然单施铵态氮处理的根系干重、平均直径较高,特别是处理后期根系直径显著高于其他处理,但根系总长度、根系表面积和体积最低,说明铵态氮抑制根系的伸长生长,促进根系的加粗生长。这可能是由于铵态氮抑制根系细胞的伸长,但不抑制根系细胞的分裂造成的[24]。根系的体积和表面积是表征根系吸收养分和水分能力的重要标准,虽然单施硝态氮处理的根系总长度与混合态氮素处理的差异没有统计学意义,但根系体积和表面积低于混合态氮素处理,因此养分和水分的吸收能力差,导致单施硝态氮处理的地上部干重显著低于铵硝配施处理。处理10d后,在所有处理中,单施铵态氮处理的根系干重相对较大,而单施硝态氮处理的根系干重最低,但两者的地上部生物量均较低,这也说明单纯以根系干重作为衡量生长的标准并不确切,必须考虑根系的形态结构。
处理结束时,铵硝比为70∶30的处理根系不仅表面积和体积最高,而且根系活力最高,这可能是导致地上部生长最优的原因。随处理时间的延长铵硝配施处理的根系活力增加,而单施铵态氮或硝态氮处理的根系活力在处理20d后下降。处理结束时,根系可溶性糖含量表现为铵硝比70∶30处理>铵硝比50∶50处理>单施硝态氮处理>铵硝比30∶70处理>单施铵态氮处理。单施铵态氮处理的根系可溶性糖含量较低可能是由于根系利用铵态氮会消耗大量可溶性糖。当营养液中硝态氮比例大于30%后,可溶性糖含量下降,可能与硝态氮影响碳水化合物在体内的运输有关[25-26]。
3.2 不同氮素形态对烟苗氮素利用的影响
王小丽等[27]对小白菜的研究发现,单施铵态氮地上部分的含氮量高于单施硝态氮的。而本研究中,长时间单施铵态氮地上部含氮量低于单施硝态氮,而根系含氮量高。处理结束时,铵态氮比例较高的处理根系含氮量较高,而硝态氮比例较高的处理叶片含氮量较高。氮素的供应与植物体内细胞分裂素含量存在密切联系,氮素含量的增加会引起木质部汁液中细胞分裂素含量的增加[28-29]。硝酸盐可诱导细胞分裂素的合成,而当植物体内细胞分裂素含量过高时,其对氮素吸收相关基因起负调控作用[30-31]。这可能是单施硝态氮植株根系含氮量较低的原因。而增加一定比例的硝态氮,可以促进细胞分裂素的合成,进而促进植株生长。处理结束时,铵硝比为100∶0和70∶30的处理根的氮素分配率显著高于其他处理,而叶的氮素分配率较低,且随硝态氮比例增加叶片氮素分配率呈增加趋势。由此可以看出,氮素形态影响各器官氮素的利用,铵态氮促进氮素在根系利用,而硝态氮则更利于促进氮素在地上部的利用。处理结束时,铵硝比为30∶70的处理地上部分的含氮量和氮素积累量较高,但地上部干重低于铵硝比70∶30的处理。这可能是由于铵态氮不仅有利于增加根系表面积和体积促进氮素吸收,也利于叶绿素的合成,进而有利于增强光合能力,但单施铵态氮消耗过多的光合产物,使地上部生长受到抑制,增加一定比例硝态氮,既可以缓解铵态氮对地上部分生长的抑制现象,积累较多的碳水化合物,也促进地上部对氮素的利用。而硝态氮比例过高,植株利用硝态氮会消耗大量的能量[26],且氮素形态的不同引起植株体内源激素水平的差异,进而抑制氮素转运蛋白的表达[31],可能导致氮素向地上部运输受到限制,影响地上部的生长[32]。
4 结论
处理15d后,单施铵态氮或硝态氮的烟苗根系长度小于铵硝配施的处理。处理结束时,单施铵态氮的烟苗根系平均直径较大,单施硝态氮的烟苗根系表面积和体积低于铵硝配施的3个处理,铵硝配施中以铵硝比70∶30的处理根系总表面积、体积及平均直径最大。说明单一形态的氮肥不利于根系的伸长生长,而铵硝配施特别是铵态氮比例较高时有利于根系的伸长及表面积和体积的增加。处理结束时,硝态氮比例较高的处理根系含氮量较低,茎和叶的含氮量较高;铵硝比70∶30的处理烟苗根系氮素积累量最高,铵硝比50∶50和铵硝比30∶70的处理茎和叶的氮积累量较高。说明,提高氮肥中硝态氮比例有利于氮素在烟苗地上部利用,而提高铵态氮比例有利于氮素在根系中积累。
[1]杨阳,钟晓敏,闫志刚,等.氮素形态对巨峰葡萄果实品质的影响[J].植物营养与肥料学报,2010,16(4):1037-1040.YANG Yang,ZHONG Xiaomin, YAN Zhigang, et al. E ff ects of nitrogen forms on nitrate accumulation and quality of“Kyoho” grape fruit[J]. Journal of Plant Nutrition and Fertili zer,2010,16(4):1037-1040.
[2]刘赵帆,张国斌,郁继华,等.氮肥形态及配比对花椰菜产量、品质和养分吸收的影响[J].应用生态学报,2013,24(7):1923-1930.LIU Zhaofan, ZHANG Guobin, YU Jihua, et al. Effects of different nitrogen forms and their ratios on broccoli yield,quality, and nutrient absorption[J]. Chinese Journal of Applied Ecology, 2013,24(7):1923-1930.
[3]Zhao Y Y, Guo L, Lu W J,et al. Expression pattern analysis of microRNA in root tissue of wheat(Triticum aestivum L.) under normal nitrogen and low nitrogen conditions[J].Journal of Plant Biochemistry and Biotechnology,2015,24(2):143-153.
[4]Bowles T M, Raab P A, Jackson L E. Root expression of nitrogen metabolism genes re fl ects soil nitrogen cycling in an organic agroecosystem[J].Plant and Soil,2015,392:175-189.
[5]Ogawa S,Valencia M O, Ishitani M, et al. Root system architecture variation in response to di ff erent NH4+contrations its association with nitrogen-de fi cient tolerance traits in rice[J].Acta Physiologiae Plantarum,2014,36:2361-2372.
[6]孙敬权,薛明德,李桐,等.剪叶育苗与明水深栽配套对烟株生长发育的影响[J].烟草科技,1998,1:36-38.SUN Jingquan, XUE Mingde, LI Tong, et al. Studies on the effects of clipping tobacco seedling and deep transplanting on plant growth and development[J].Tobacco Science &Technology,1998,1:36-38.
[7]Marschner H, Hausslings M, George E. Ammonium and nitrate uptake rates and rhizosphere pH in non-mycorrhizal roots of Norway sprucePicea abies(L.)[J].Trees,1991,5:14-21.
[8]Sattelmacher B, Klotz F, Marschner H. Influence of the nitrogen level on root growth and morphology of two potato varieties differing in nitrogen acquisition[J].Plant and Soil,1990,123:131-137.
[9]姜琳琳,韩立思,韩晓日,等.氮素对玉米幼苗生长、根系形态及氮素吸收利用效率的影响[J].植物营养与肥料学报,2011,17(1):247-253.JIANG Linlin, HAN Lisi, HAN Xiaori, et al. Effects of nitrogen on growth, root morphological traits, nitrogen uptake and utilizaiton efficiency of maize seedlings[J]. Journal of Plant Nutrition and Fertilizer,2011,17(1):247-253.
[10]刘国顺,赵春华,王彦亭,等.施氮量对烤烟根系发育和某些生理指标的影响[J].河南农业大学学报,2007,41(2):134-137.LIU Guoshun, ZHAO Chunhua, WANG Yanting, et al. E ff ects of nitrogen rates on root growth and some physiological indexes of fl ue-cured tobacco[J]. Journal of Henan Agricultural University, 2007, 41(2):134-137.
[11]郭培国,陈建军,郑燕玲,等. 不同施氮水平下烤烟对氮素的利用研究[J].热带亚热带土壤科学,1996,5(3):145-148.GUO Peiguo, CHEN Jianjun, ZHENG Yanling, et al. Study on utilization of nitrogen by flue-cured tobacco at different nitrogen fertilizer levels[J]. Tropical and Subtropical Soil Science,1996,5(3):135-148.
[12]严君,韩晓增,祖伟. 不同形态氮肥对大豆根系形态及磷效率的影响[J]. 大豆科学,2010,29(6):1003-1007.YAN Jun, HAN Xiaozeng, ZU Wei. E ff ects of nitrogen forms on root morphology and phosphorous efficiency in soybean(Glycinemax L.)[J]. Soybean Science, 2010, 29(6):1003-1007.[13]Walch-Liu P, Forde B G. Nitrate signaling mediated by the NRT1.1 nitrate transporter antagonizes L-glutamate-induced changes in root architecture[J]. Plant Journal,2008,54:820-828.[14]Tian Q, Chen F, Liu J, et al. Inhibition of maize root growth by high nitrate supply is correlated with reduced IAA levels in roots[J]. Journal of Plant Physiology,2008,165:942-951.
[15]Domenicano S, Coll L, Messier C,et al. Nitrogen forms a ff ect root structure and water uptake in the hybrid popular[J].New Forests,2011,42:347-362.
[16]Mclnenly L E, Merrill E H, Cahill J F, et al. Festuca campestris alters root morphology and growth in response to simulated grazing and nitrogen form[J].Functional Ecology,2010,24:283-292.
[17]晁逢春,张福锁,杨宇虹,等. 影响烟草根系的几个因素探讨[J].中国烟草科学,2003,2:5-8.CHAO Fengchun, ZHANG Fusuo, YANG Yuhong, et al. E ff ect of factors on the growth of tobacco root[J]. Chinese Tobacco Science,2003,2:5-8.
[18]李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000.LI Hesheng. Principle and technology of plant physiological and biochemical experiments[M]. Beijing: Higher Education Press,2000.
[19]齐曼·尤努斯,木合塔尔·扎热,塔依尔·艾合买提.干旱胁迫下尖果沙枣幼苗的根系活力和光合特性[J].应用生态学报,2011,22(7):1789-1795.QIMAN·Yunus, MUHTAR·Zari, TAYER·Ahmat. Root activity and photosynthetic characteristics of Elaeagnus oxycarpa seedlings under drought stress[J]. Chinese Journal of Applied Ecology, 2011,22(7):1789-1795.
[20]丁红,张智猛,戴良香,等.干旱胁迫对花生根系生长发育和生理特性的影响[J].应用生态学报,2013,24(6):1586-1592.DING Hong, ZHANG Zhimeng, DAI Liangxiang, et al. E ff ects of drought stress on the root growth and development and physiological characteristics of peanut[J]. Chinese Journal of Applied Ecology, 2013, 24(6):1586-1592.
[21]徐国伟,王贺正,翟志华,等.不同水氮耦合对水稻根系形态生理、产量与氮素利用的影响[J].农业工程学报,2015,31(10):132-141.XU Guowei, WANG Hezheng, ZHAI Zhihua, et al. Effect of water and nitrogen coupling on root morphology and physiology, yield and nutrition utilization for rice[J].Transactions of the Chinese Society of Agricultural Engineeri ng,2015,31(10):132-141.
[22]乔云发,苗淑杰,韩晓增.氮素形态对大豆根系形态性状及释放H+的影响[J].大豆科学,2006,3:265-269.QIAO Yunfa, MIAO Shujie, HAN Xiaozeng. Effects of nitrogen forms on the root morphology and proton extrusion in soybean[J].Soybean Science,2006,3:265-268.
[23]刘晓静,叶芳,张晓玲.外源氮素形态对紫花苜蓿不同生育期根系特性的影响[J].草业学报,2015,24(6):53-63.LIU Xiaojing, YE Fang, ZHANG Xiaoling. Effects of exogenous nitrogen forms on root characteristics of alfalfa at different growth stages[J].Acta Prataculturae Sinica,2015,24(6):53-63.
[24]Li Q, Li B H,Shi W M,et al. Root growth inhibition byin Arabidopsis is mediated by the root tip and is linked toefflux and GMPase activity[J].Plant Cell and Environment,2010,33:1529-1542.
[25]戴廷波,曹卫星,孙传范,等.增铵营养对小麦光合作用及硝酸还原酶和谷氨酰胺合成酶的影响[J].应用生态学报,2003,14(9):1529-1532.DAI Tingbo, CAO Weixing, SUN Chuanfan, et al. Effect of enhanced ammonium nutrition on photosynthesis and nitrate reductase and glutamine synthetase activities of winter wheat[J]. Chinese Journal of Applied Ecology,2003,14(9):1529-1532.
[26]张辰明,徐烨红,赵海娟,等.不同氮形态对水稻苗期氮素吸收和根系生长的影响[J].南京农业大学学报,2011,34(3):72-76.ZHANG Chenming, XU Yehong, ZHAO Haijuan, et al. E ff ects of di ff erent nitrogen forms on nitrogen uptake and root growth of rice at the seedling stage[J]. Journal of Nanjing Agricultural University, 2011, 34(3):72-76.
[27]王小丽,杨丹妮,黄丹枫.氮素形态对小白菜生长和碳氮积累的影响[J].应用生态学报,2012,23(4):1042-1048.WANG Xiaoli, YANG Danni, HUANG Danfeng. Effect of different applied nitrogen forms on pakchoi (Brassica chinensis) growth and its carbon and nitrogen accumulation[J].Chinese Journal of Applied Ecology, 2012, 23(4):1042-1048.
[28]Singh S, Letham D S, Zhang R, et al. Cytokinin biochemistry in relation to leaf senescence:IV. E ff ect of nitrogenous nutrient on cytokinin levels and senescence of tobacco leaves[J].Physiologia Plantarum, 1992,84:262-268.
[29]Takei K, Sakakibara H, Taniguchi M, et al. Nitrogen dependent accumulation of cytokinins in root and the translocation to leaf: implication of cytokinin species that induces gene expression of maize response regulator[J]. Plant and Cell Physiolgoy,2001,42:85-93.
[30]Miyawaki K, Tarkowski P, Matsumoto K M, et al. Roles of Arabidopsis ATP/ADP isopentenyltransferases and tRNA isopentenyltransferases in cytokinin biosynthesis[J]. Proceeding of the National Academy of Sciences, USA,2006,103:16598-16603.
[31]Takei K, Toru K, Mikiko K, et al. Hormonal control of nitrogen acquisition: roles of auxin, abscisic acid, and cytokinin[J].Journal of Experimental Botany,2011,62(4):1399-1409.
[32]Gao Y P, Motosugi H, Sugiura A. Rootstock e ff ects on growth and fl owering in young apple trees grown with ammonium and nitrate nitrogen[J]. Journal of American Society of Horticultural Science,1992,117:446-452.
Effect of different nitrogen forms on tobacco seedling root growth and nitrogen utilization
XING Yao, MA Xinghua
Tobacco Research Institute, Chinese Academy of Agricultural Sciences, Key Laboratory of Tobacco Biology and Processing,Ministry of Agriculture, Qingdao 266101, China
In order to reveal the in fl uence mechanism of di ff erent nitrogen forms on tobacco seedling root growth and nitrogen utilization,and to provide a theoretical basis for rational use of fertilizer in tobacco cultivation, hydroponic experiment with equal amounts of N in fi ve di ff erent ratios of NH4+-N:NO3--N (100:0, as 100-0AN; 70:30, as 70-30AN; 50:50, as 50-50AN; 30:70 as 30-70AN; 0:100, as 0-100AN)was conducted to study the di ff erences of root morphology and nitrogen utilization and distribution of tobacco seedlings by using fl uecured cultivar NC55. Results indicated that shoot dry weight, total dry weight, root total length, root total surf area and volume, root soluble sugar content, nitrogen content, nitrogen accumulation of stem and leaf and nitrogen distribution ratio of stem and leaf of 100-0AN were lowest among all treatments 25 days after ammonium nitrogen treatment (DAT). Root dry weight, root average diameter, root activity, root nitrogen content and accumulation of 0-100AN were the lowest among all treatments 25 days after nitrate nitrogen treatment. Within 20 DAT, root dry weight, nitrogen accumulation of root and plant total dry weight of 50-50AN were the highest. On 25 DAT, root and shoot dry weight, root total surf area and volume, root activity, root soluble sugar content, root nitrogen content and accumulation of 70-30AN were the highest; nitrogen content and nitrogen accumulation of leaf of 30-70AN were the highest. Tobacco seedling growth and nitrogen utilization were inhibited with single ammonium or single nitrate under nitrogen concentration of 2 mmol/L, but mixture of ammonium and nitrate promoted growth and nitrogen utilization. The ratio of ammonium to nitrate at 70:30 could synergistically improve growth of root and shoot, the ratio of ammonium to nitrate at 30:70 could promote nitrogen utilization and absorption of shoot.
tobacco seedlings; nitrogen form; root; nitrogen utilization
邢瑶 ,马兴华. 氮素形态对烟苗根系生长及氮素利用的影响[J]. 中国烟草学报,2016,22(4)
公益性行业(农业)科研专项“烟草增香减害关键技术研究与示范”项目(201203091);国家烟草专卖局低危害烟叶开发项目(110201101006(ts-06));中国农业科学院科技创新工程(ASTIP-TRIC03)
邢 瑶(1990— ),硕士研究生,研究方向为作物栽培,Email:xingyao19901205@163.com
马兴华(1979— ),副研究员,博士,主要从事烟草栽培与施肥研究,Email:maxinghua@caas.cn
2015-12-30
:XING Yao, MA Xinghua. E ff ect of di ff erent nitrogen forms on tobacco seedling root growth and nitrogen utilization [J]. Acta Tabacaria Sinica, 2016,22(4)
猜你喜欢
中国畜牧业(2022年20期)2022-11-19
山西农业大学学报(自然科学版)(2021年6期)2021-12-28
腐植酸(2021年2期)2021-12-04
云南农业(2021年5期)2021-06-19
上海金属(2020年4期)2020-08-25
重型机械(2020年2期)2020-07-24
福建茶叶(2019年12期)2020-01-19
江苏农业科学(2019年20期)2019-12-23
分析化学(2018年8期)2018-11-01
中国集体经济(2017年11期)2017-04-24
