
干旱胁迫和施肥对巨能草生物量及C、N、P积累与分配的影响
2016-11-10 04:51张明锦陈良华胡相伟
西北农林科技大学学报(自然科学版) 2016年9期
张明锦,陈良华,张 健,胡相伟,徐 睿
(四川农业大学 生态林业研究所 林业生态工程重点实验室,四川 成都611130)
干旱胁迫和施肥对巨能草生物量及C、N、P积累与分配的影响
张明锦,陈良华,张健,胡相伟,徐睿
(四川农业大学 生态林业研究所 林业生态工程重点实验室,四川 成都611130)
【目的】 研究干旱胁迫和施肥对巨能草生物量及C、N、P积累与分配的影响,为巨能草栽培中的水肥管理提供理论依据。【方法】 以巨能草为研究对象,采用盆栽试验,利用控水、控肥的试验方法,设计干旱(田间持水量的25%)和正常水分(田间持水量的75%)2个水分梯度以及低肥(复合肥施肥量为2 g/盆)、中肥(复合肥施肥量为4 g/盆)、高肥(复合肥施肥量为8 g/盆)3个养分水平,进行完全随机组合试验,共计6个处理,研究不同处理下巨能草生物量及其分配比例,C、N、P积累与分配特征,N、P吸收和利用效率,以及C∶N、C∶P、N∶P的变化。【结果】 在正常水分条件下,随着施肥量的增加,巨能草的地上、地下生物量和总生物量都显著增加;干旱胁迫后,巨能草地上和地下生物量的积累显著降低,施肥对巨能草的地上、地下生物量和总生物量也有显著影响。在正常水分条件下,施肥对巨能草地上和地下生物量的分配比例没有显著影响;在干旱条件下,除低肥处理外,中施肥量和高施肥量均能显著提高巨能草地下生物量的分配比例。施肥和干旱胁迫处理对C、N、P积累量及分配比例也有一定的影响,与正常水分条件相比,干旱胁迫后巨能草的C、N、P积累总量均降低。在正常水分条件下,尽管低肥处理的N、P吸收效率显著低于中、高肥处理,但其N、P利用效率却显著高于中、高肥处理;干旱条件下,不同施肥处理的N、P吸收效率和P利用效率并无显著差异,但N的利用效率随着施肥量的增加而降低。正常水分条件下,施肥处理对地上部分的C∶P、N∶P无显著影响,但高肥处理显著降低了地下部分的N∶P,中、高肥处理显著降低了地上部分的C∶N和地下部分的 C∶P;干旱条件下,施肥处理对地上和地下部分的C∶P及N∶P的影响都不显著,但高肥处理显著降低了地上和地下部分的C∶N。【结论】 干旱胁迫处理减少了巨能草的生物量及其对C、N、P的积累,影响了C、N、P的分配,施肥在一定程度上降低了干旱胁迫对植物生长的影响,在巨能草的施肥管理中,应更多地注重对N素的管理。
干旱胁迫;施肥;巨能草;生物量;养分积累和分配
很多资料预测,到 21 世纪末期会出现更频繁更严重的极端气候事件[1-2]。而在干旱与半干旱地区,由于其贫瘠和脆弱的土壤特质,使得其特性对大范围的气候环境事件变得更为敏感[3-4]。然而,在干旱与半干旱系统中,水分并不是限制植物生长的惟一因子,植物的生长还常常受到养分(尤其是N和P)的限制[5-6]。由于水分与养分有效性之间存在密切关系,养分有效性的缺失在未来越来越干旱的环境下也可能变得越来越严重[7-8]。现在干旱与半干旱系统管理者面临的主要问题是怎样在提高资源利用效率的同时增加C的固定、减轻干旱环境对植物生长带来的伤害,从而提高干旱与半干旱地区植被的覆盖度,改善干旱与半干旱地区的生态环境,其中,施肥是最常见的农艺措施。有研究表明,适当施肥可以改善土壤的理化特性[9],增加受限制养分的有效性,进而可能改变系统特性,提高受限制资源的利用效率, 促进植物的生长, 因而可能成为贫瘠及干旱与半干旱地区改善受限制资源有效性、恢复植被的潜在措施和可行性方法[10-12]。有研究也认为,相对于改变水分有效性和水分利用效率,改变受限养分有效性可能更能改变群落或系统特性[9,13-15]。
巨能草(PueliasineseRoxb)为一年种、多年生禾本科植物,由二倍体美洲狼尾草(又称御谷、珍珠谷)作母本与四倍体象草(又称紫狼尾草)杂交选育而成,故又称杂交狼尾草,属三倍体C4植物[16]。其根系生长迅速,能在较短时间内形成须根网络,牢固地锁住水分和泥土,防止水土流失,对绿化荒山荒坡、防风固沙都具有积极作用[17]。近年来,其在生态环境治理(如干旱的荒山、荒坡、沙地)、水土保持和种草养畜等方面得到了广泛的推广应用[17-18]。
但目前缺乏对这一草种应对干旱胁迫的适应性研究,尤其是对干旱及不同养分条件下巨能草生长、资源积累与分配等方面的研究还较少。本研究采用盆栽试验,通过控水、控肥的方法,探讨干旱和施肥对巨能草生物量及C、N、P积累与分配的影响,旨在阐明巨能草在干旱和不同养分条件下的养分利用效率、营养平衡及受限特征,为干旱环境下巨能草的水肥管理提供指导。
1 材料与方法
1.1试验材料
选择健康的巨能草草种,将茎干截成长度大小一致的茎段(约20 cm),将其扦插在装有10 kg混合培养基质(m(冲积土)∶m(细沙)=3∶1)的聚乙烯圆台型花盆中。圆台型花盆的口径为30 cm,深25 cm,每盆扦插1截。于2013年5月底进行扦插,遮雨棚中生长。待其发芽缓苗后,于6月底选择48株高度和长势一致且健壮无病虫害的幼苗进行控制试验。培养基质背景值:基质含水量为23%,最大含水量为52%,pH值为8.86,有机质含量10.13 g/kg,铵态氮3.82 mg/kg,硝态氮0.22 mg/kg,有效磷37.72 mg/kg,速效钾11.47 mg/kg。
1.2试验处理
试验采用盆栽方式,以水分和施肥2个因素进行完全随机组合试验(表1)。其中,水分梯度的设置依据一般作物最适土壤含水量是土壤田间持水量75%的常规,所以将田间持水量的75%设为对照(即正常水分),将田间持水量的25%设为干旱处理;养分梯度依据一般作物的施肥量(200 kg/hm2),并考虑到每盆只栽种1株巨能草以及花盆的大小设置。具体处理过程为:根据水分梯度将植株分为C(田间持水量的75%,作为对照水分,模拟正常水分条件)和D(田间持水量的25%,模拟极端干旱条件)2组,每组24株,再将每组分成L(低施肥量,施肥量为2 g/盆)、M(中施肥量,施肥量为4 g/盆)、H(高施肥量,施肥量为8 g/盆)3个小组,即共有CL、CM、CH、DL、DM、DH等6个处理,每处理8个重复。6月底开始进行水分处理, 1个月后(8月初)在每盆植株的四周均匀地施撒设计中相应的肥量。因为这一阶段正值巨能草生长旺盛阶段,所以更利于监测施肥对植物生长和干旱胁迫的效应。所用的复合肥为美丰比利夫复合肥,N、P2O5、K2O含量比例为17-17-17。试验在四川农业大学成都校区的生态林业研究所基地大棚内进行,试验时间是从2013年6月底到8月底,在此期间,采用每天称重补水的方法控制盆中土壤水分含量。除试验因素(水分、养分)外,其他条件尽可能保持一致。

表 1 干旱胁迫与施肥对巨能草相关指标影响的试验设计组合Table 1 Combined treatments of drought stress and fertilization
1.3测定指标及方法
在2013年8月底试验处理结束后,按地上和地下部分分别收集每株巨能草的各组织样品, 105 ℃杀青30 min, 65 ℃烘干。待其烘干至质量恒定后,分别称量地上和地下部分的干质量。之后将样品粉碎,研磨,过孔径1 mm筛,放置于干燥器中储存,以备各元素的测定。
巨能草养分含量的测定参照《中华人民共和国林业行业标准LY/T 1271-1999》。有机碳含量用重铬酸钾氧化外加热法测定。样品用硫酸-高氯酸消煮,用凯氏定氮仪(浙江托普仪器有限公司KDN系列)测定全N含量。用钼锑抗比色法测定全P含量。C、N、P 含量乘以相应部位的生物量即可得到该部位的C、N、P 积累量。所有部位的积累量之和即为C、N、P的总积累量。C∶N、C∶P和N∶P则用每株的C、N积累量除以相应的N、P积累量。养分利用效率是植物吸收单位养分所获得的干物质量。采用Vitousek[19]指数法,即植物的干物质量(生物量)与植物养分贮量的比值计算巨能草的N、P利用效率(NUEN、NUEP)。N和P的吸收效率(NUtEN和 NUtEP)由养分元素总积累量除以细根生物量求得[20-21]。以上所有分析均做 3个重复。
1.4数据统计分析
采用SPSS 18.0统计软件对数据进行单因素方差分析(one-way ANOVA),并用Duncan’s(D)法进行处理间差异显著性检验。
2 结果与分析
2.1干旱胁迫和施肥对巨能草生物量及分配比例的影响
对巨能草生物量(图1)的观测结果表明,在正常水分条件下,巨能草的地上、地下生物量和总生物量都受到了施肥处理的显著影响(P<0.05),表现为随着施肥量的增加,巨能草的地上、地下生物量和总生物量都显著增加。干旱条件下,施肥对巨能草的地上、地下生物量和总生物量也有着显著影响(P<0.05)。此外,在低肥条件下,干旱处理下的地上、地下生物量和总生物量都显著低于正常水分;而在中、高肥条件下,虽然干旱处理下的地上生物量和总生物量均显著低于正常水分,但地下生物量与正常水分条件下并无显著差异。
从巨能草生物量的分配比例(图1)来看,在正常水分条件下,施肥对巨能草地上和地下生物量的分配没有显著影响(P>0.05)。但在干旱条件下,除低肥处理外,中肥和高肥处理均能显著提高巨能草地下生物量的分配比例,进而增大根冠比。
可见,干旱胁迫显著降低了巨能草地上和地下生物量的积累,施肥显著提高了巨能草地上和地下生物量的积累。干旱环境下,施肥显著增加了根冠比,可以增强巨能草对干旱环境的适应能力。

图 1干旱胁迫与施肥对巨能草生物量及其分配比例的影响

Fig.1Effects of different combinations of drought stress and fertilization on biomass ofPueliasineseRoxb and its allocation pattern
different treatments (P<0.05);different lowercase letters indicate significant differences between below-ground under different treatments(P<0.05).The same below
2.2干旱胁迫和施肥对巨能草C、N、P积累与分配特征的影响
图2显示,正常水分和干旱条件下,中、高施肥都显著增加了巨能草地上和地下部分的C积累量,但对C的分配比例无显著影响(P>0.05)。与正常水分条件相比,干旱处理显著降低了巨能草地上部分的C积累量。中、低肥环境下,干旱胁迫处理地下部分的C积累量与正常水分条件下无显著差异,但其地下部分的C分配比例显著高于正常水分处理。
由图2还可知,正常水分条件下,中、高肥显著提高了巨能草地上和地下部分的N积累量。干旱条件下,高肥处理巨能草地上和地下部分N积累量也显著高于低肥处理。正常水分和干旱条件下,施肥对N的分配比例无显著影响(P>0.05)。中、高肥环境下,干旱胁迫显著降低了巨能草地上部分N积累量;与正常水分条件相比,干旱胁迫下高肥处理巨能草地上部分N分配比例也显著降低。
图2显示,正常水分条件下,低肥处理的巨能草地上和地下部分P积累量都显著低于高肥处理,但低肥处理的地上部分P分配比例显著高于高肥处理。另一方面,高肥环境下,与正常水分条件相比,干旱胁迫显著降低了地上部分P积累量,但对地下部分P积累量并无显著影响。除中肥条件外,干旱胁迫下地下部分P分配比例都显著高于正常水分条件。
图 2干旱胁迫与施肥对巨能草C、N、P积累及其分配特征的影响
Fig.2C, N and P accumulation and distribution characteristics ofPueliasineseRoxb under different combinations of drought stress and fertilization
2.3干旱胁迫和施肥对巨能草N、P吸收与利用效率及化学计量比的影响
干旱胁迫和施肥处理下,巨能草的N、P吸收和利用效率见表2。由表2可知,正常水分条件下,尽管低肥处理的N、P吸收效率显著低于中、高肥处理,但其N、P利用效率却显著高于中、高肥处理。而干旱条件下,不同施肥处理下的N、P吸收效率和P利用效率并无显著差异,但N的利用效率随着施肥量的增加而降低。低肥条件下,干旱处理的P吸收效率是正常水分的1.5倍,但N吸收效率之间无显著差异,干旱处理的N、P利用效率都显著低于正常水分;中肥条件下,正常水分和干旱处理间的P吸收效率和N利用效率都无显著差异,但正常水分下的N吸收效率和P利用效率都显著高于干旱处理;高肥条件下,正常水分和干旱处理间的P吸收效率无显著差异,但正常水分下的N吸收效率和N、P利用效率都显著高于干旱处理。
表3显示,正常水分条件下,施肥处理对地上部分的C∶P、N∶P无显著影响,高肥处理显著降低了地下部分的N∶P,中、高肥处理显著降低了地上部分的C∶N和地下部分的C∶P。干旱条件下,施肥处理对地上和地下部分的C∶P及N∶P的影响都不显著,但高肥处理显著降低了地上和地下部分的C∶N。低肥条件下,干旱胁迫显著降低了地下部分的C∶P和N∶P;中肥条件下,干旱胁迫显著提高了地下部分的C∶N;高肥条件下,干旱胁迫显著降低了地下部分的C∶N。
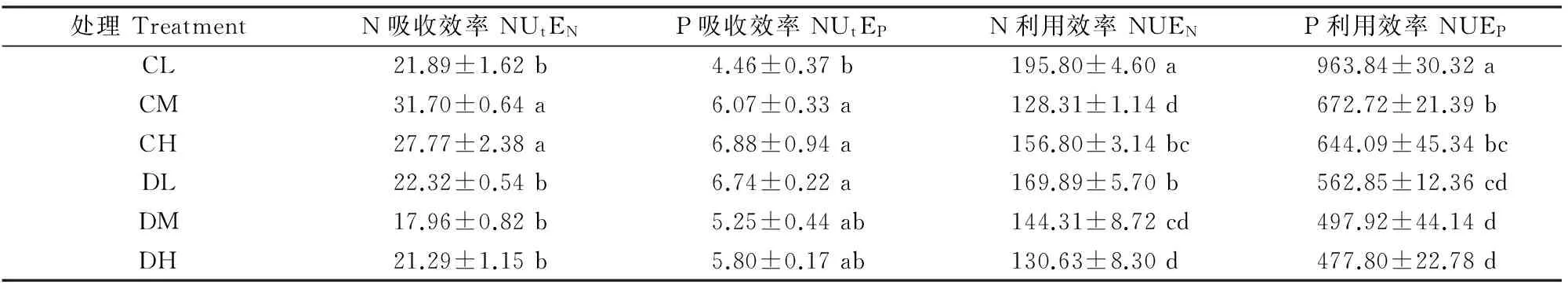
表 2 干旱胁迫与施肥对巨能草N、P吸收和利用效率的影响Table 2 Nitrogen and phosphorus absorption efficiency (NUtEN,NUtEP)and use efficiency (NUEN,NUEP) ofPuelia sinese Roxb under different combinations of drought stress and fertilization
注:同列数据后标不同小写字母表示差异显著(P<0.05),下表同。
Note:Different lowercase letters indicate significant difference atP<0.05.The same below.

表 3 干旱胁迫与施肥对巨能草C∶N、C∶P和N∶P的影响Table 3 C∶N,C∶P,and N∶P of Puelia sinese Roxb under different combinations of drought stress and fertilization
3 讨论与结论
3.1干旱胁迫和施肥对巨能草生物量积累与分配的影响
在本研究中,不同施肥条件下,干旱处理巨能草的地上生物量和总生物量都显著低于正常水分。可见,干旱胁迫显著降低了巨能草生物量的积累,影响了巨能草的生产潜力,这与前人的一些研究结果[22-23]相似。但为了能在干旱环境下得以生存,植物形成了相应的生存防御系统。其中,植物生物量的积累与分配特征是植物适应环境的一种重要生存机制[24]。由于各器官的位置和功能不同,它们对环境变化的响应也有所差异。植物地下部分的根系不仅是植物吸收水分和盐类的主要器官,而且是多种物质同化、转化和合成的重要器官[25],并且能最早最直接地感受到土壤水分含量的变化,从而能对干旱胁迫作出迅速反应,这是植物抗旱性的基础。这在本研究中得到了进一步证实:中、高肥条件下,干旱胁迫处理地下生物量的分配比例都显著高于正常水分,其根冠比显著增大。这些研究结果说明,与正常水分条件相比,在干旱条件下同化物质优先供给地下部分,促进其生长,以便吸收更多的水分来满足植物生长的需求;同时也说明巨能草在干旱环境下表现出了一定的适应性,通过增加同化物质向根系的分配,扩大根系表面积,以便于吸取土壤深层的水分,维持体内水分平衡。
施肥是保证贫瘠地区植物养分需求和植物生产力最常见的管理方法[10],能在一定程度上增加土壤有效养分。土壤有效养分改变的同时也会改变植物各方面的适应对策,而生长和形态学方面的适应特性则是植物适应环境的基本机制[26]。本研究中,在正常水分和干旱胁迫下,巨能草的地下、地上生物量和总生物量都随着施肥量的增加而显著增加。在正常水分条件下,施肥对巨能草生物量的分配比例并无显著影响;但在干旱胁迫下,施肥量增加显著提高了巨能草地下生物量的分配比例。这些研究结果说明,适宜施肥能够促进养分吸收器官 (地下部分)的生长,进而促进光合器官(地上部分)的生长。并且在干旱胁迫下,施肥能显著促进巨能草地下部分根系的生长,进而增加其生物量,增大根冠比,从而在一定程度上降低了干旱胁迫对植物生长和生物量积累的影响。
3.2干旱胁迫和施肥对巨能草C、N、P积累与分配的影响
为了适应胁迫环境导致的资源限制,植物会通过改变资源分配与利用方式,将有限的资源分配到不同结构和功能的器官上,从而更好地适应环境[27]。本研究中,干旱胁迫显著降低了巨能草地上部分的C、N、P积累量。与正常水分处理相比,低肥环境下,干旱胁迫显著提高了地下部分C、P分配比例;中肥环境下,干旱胁迫显著提高了地下部分C的分配比例;高肥环境下,干旱胁迫显著提高了地下部分N、P养分的分配比例。施肥是提供外源养分的重要管理措施。在本研究中,正常水分和干旱环境下,施肥都能显著提高巨能草地上和地下部分的C、N、P积累量,但对C、N、P的分配比例并无显著影响。这些结果表明,一定程度的干旱胁迫会抑制植物的C、N、P积累,进而迫使植物通过改变C、N、P的分配与利用方式,以适应干旱环境。虽然施肥能促进植物对C、N、P的积累,但不会影响C、N、P的资源分配。
3.3干旱胁迫和施肥对巨能草养分吸收及利用效率的影响
本研究中,与正常水分环境相比,干旱胁迫总体上显著降低了巨能草对N的吸收和对N、P的利用效率,干旱胁迫对巨能草的养分吸收和利用效率产生了显著的负面影响。这是因为植物N、P等养分的利用效率容易受土壤湿度和水分的影响,土壤水分条件越好,植物对养分的利用效率越高[28]。本研究中,在正常水分环境下,低肥处理巨能草的N、P吸收效率最低,但为了满足其生长对养分的需求,巨能草提高了对N、P的利用。干旱环境下,低肥处理N、P利用效率明显较高,这是因为低肥的贫瘠土壤会刺激巨能草对N、P的利用效率,以满足植物的生长需求。这在一定程度上证实了植物在营养元素供应缺乏的情况下往往具有较高的养分利用效率[29],这是贫瘠养分状态的一种生存策略[30]。
3.4干旱胁迫和施肥对巨能草化学计量比及养分限制的影响
本研究中,正常水分条件下,高肥显著降低了地上部分的C∶N及地下部分的C∶P和N∶P。干旱胁迫下,高肥处理也显著降低了地上和地下部分的C∶N,但对地上和地下部分的C∶P与N∶P影响不显著。这表明随着施肥量的增加,巨能草降低了对N和P的利用效率。N和P是陆地生态系统植物生长的主要限制性元素。植物N∶P特征能够较好地反映N、P养分的限制作用[31-32]。有研究表明,植物器官N∶P大于16时,植物生产力受P限制较大,N∶P小于14时则受N限制较多,N∶P为14~16时则植物生产力受N和P共同限制[33-34]。本研究中,不同干旱胁迫与施肥处理下巨能草地上和地下部分的N∶P都小于14,且施肥对巨能草的N∶P并无显著影响,表明巨能草的生长更多地受到了N的限制,且施用复合肥并不能有效地解决N对巨能草生长的限制。因此,在施肥管理中,应更多地注重对N肥的施用,且以施加外源氮为主。
[1]Saxe H,Cannell M G R,Johnsen Ø,et al.Tree and forest functioning in response to global warming [J].New Phytol,2001,149:369-400.
[2]Monclus R,Dreyer E,Villar M,et al.Impact of drought on productivity and water use efficiency in 29 genotypes ofPopulusdeltoids×Populusnigra[J].New Phytol,2006,169:765-777.
[3]Stursova M,Crenshaw C L,Sinsabaugh R L.Microbial responses to long-term N deposition in semiarid grassland [J].Microb Ecol,2006,51:90-98.
[4]Reyes-Reyes B G,Alcántara-Hernández R,Rodríguez V,et al.Microbial biomass in a semi arid soil of the central highlands of Mexico cultivated with maize or under natural vegetation [J].Eur J Soil Biol,2007,43:180-188.
[5]Hooper D U,Johnson L.Nitrogen limitation in dryland ecosystems:responses to geographical and temporal variation in precipitation [J].Biogeochemistry,1999,46:247-293.
[6]Maestre F T,Valladares F,Reynolds J F.Is the change of pla-nt-plant interactions with abiotic stress predictable? A meta-analysis of field results in arid environments [J].J Ecol,2005,93:748-757.
[7]Vitousek P M,Turner D,Parton W,et al.Litter decomposition on the Mauna Loa Environmental Matrix,Hawaii:patterns,mechanisms and models [J].Ecology,1994,75:418-429.
[8]Schimel D S,Vemap P,Braswell B H.Continental scale variability in ecosystem processes:models,data,and the role of disturbance [J].Ecol Monograph,1997,67:251-271.
[9]James J J,Tiller R L,Richards J H.Multiple resources limit plant growth and function in a saline-alkaline desert community [J].Journal of Ecology,2005,93:113-126.
[10]Baligar V C,Fageria N K,He Z L.Nutrient use efficiency in plants [J].Communications in Soil Science and Plant Analysis,2001,32(7/8):921-950.
[11]Singh S P,Bargali K,Joshi A,et al.Nitrogen resorption in leaves of tree and shrub seedlings in response to increasing soil fertility [J].Current Science,2005,89(2):389-396.
[12]Dang T H,Cai G X,Guo S L,et al.Effects of nitrogen management on yield and water use efficiency of rain fed wheat and maize in northwest China [J].Pedosphere,2006,16(4):495-504.
[13]Melillo J M,Mcguire A D,Kicklighter D W,et al.Global climate change and terrestrial net primary production [J].Nature,1993,363:234-240.
[14]Smith D S,Huxman T E,Zitzer S F,et al.Elevated CO2increases productivity and invasive species success in an arid ecosystem [J].Nature,2000,408:79-82.
[15]Xiao C,Janssens I A,Liu P,et al.Irrigation and enhanced soil carbon input effects on below-ground carbon cycling in semiarid temperate grasslands [J].New Phytol,2007,174:835-846.
[16]Magcale-Macandog D B,Predo C D,Menz K M,et al.Napier grass strips and livestock:a bioeconomic analysis [J].Agroforestry Systems,1998,40(1):41-58.
[17]马崇坚,刘发光.皇竹草在生态环境治理中的应用研究进展 [J].中国水土保持,2012(1):41-44.
Ma C J,Liu F G.The research progress ofPennisetumhydridumin the application of ecological environmental governance [J].Soil and Water Conservation in China,2012(1):41-44.
[18]谢永良,何光武,熊建平.皇竹草生产性能与开发利用 [J].四川畜牧兽医,1999(4):23.
Xie Y L,He G W,Xiong J P.The production performance and utilization ofPennisetumhydridum[J].Sichuan Animal and Veterin Sciences,1999(4):23.
[19]Vitousek P M.Nutrient cycling and nutrient use efficiency [J].The American Naturalist,1982,119:553-572.
[20]Sorgonà A,Abenavoli M R,Gringeri P G,et al.A comparison of nitrogen use efficiency definitions inCitrusrootstocks [J].Scientia Horticulturae,2006,109:389-393.
[21]Elliot G C,Läuchli A.Phosphorus efficiency and phosphate-iron interactions in maize [J].Agronomy Journal,1985,77:399-403.
[22]闫海霞,方路斌,黄大庄.干旱胁迫对条墩桑生物量分配和光合特性的影响 [J].应用生态学报,2011,22(12):3365-3370.
Yan H X,Fang L B,Huang D Z.Effects of drought stress on the biomass distribution and photosynthetic characteristics of cluster mulberry [J].Chinese Journal of Applied Ecology,2011,22(12):3365-3370.
[23]安玉艳,梁宗锁,郝文芳.杠柳幼苗对不同强度干旱胁迫的生长与生理响应 [J].生态学报,2011,31(3):716-725.
An Y Y,Liang Z S,Hao W F.Growth and physiological responses of thePeriplocasepiumBunge seedlings to drought stress [J].Acta Ecologica Sinica,2011,31(3):716-725.
[24]吴福忠,鲁叶江,杨万勤,等.缺苞箭竹密度对养分元素贮量、积累与分配动态的影响 [J].生态学报,2005,25(7):1663-1669.
Wu F Z,Lu Y J,Yang W Q,et al.Effects ofFargesiadenudatedensity on its nutrient element stocks,accumulation and allocation dynamics [J].Acta Ecologica Sinica,2005,25(7):1663-1669.
[25]李朝苏,刘鹏,蔡妙珍,等.荞麦对酸铝胁迫生理响应的研究 [J].水土保持学报,2005,19(3):105-109.
Li C S,Liu P,Cai M Z,et al.Physiological response of buckwheat to acid-aluminum stress in growth [J].Journal of Soil Water Conservation,2005,19(3):105-109.
[26]Patterson T B,Guy R D,Dang Q L.Whole plant nitrogen and water relations traits,and their associated tradeoffs,in adjacent muskeg and upland boreal spruce species [J].Oecologia,1997,110:160-168.
[27]吴福忠,包维楷,吴宁.外源施 N对干旱河谷白刺花 (Sophoradavidii)幼苗生长,生物量及C、N、P积累与分配的影响 [J].生态学报,2008,28(8):3817-3824.
Wu F Z,Bao W K,Wu N.Growth,accumulation and partitioning of biomass,C,N and P ofSophoradavidiiseedlings in response to N supply in dry valley of upper Minjiang River [J].Acta Ecologica Sinica,2008,28(8):3817-3824.
[28]邢雪荣,韩兴国,陈灵芝.植物养分利用效率研究综述 [J].应用生态学报,2000,11(5):785-790.
Xing X R,Han X G,Chen L Z.A review on research of plant nutrient use efficiency [J].Chinese Journal of Applied Ecology,2000,11(5):785-790.
[29]Bridgham S D,Pastor J,Mc Claugherty C A,et al.Nutrient-use efficiency:a litterfall index,a model,and a test along a nutrient-availability gradient in North Carolina Peatlands [J].American Naturalist,1995,145:1-21.
[30]Bowman W D.Accumulation and use of nitrogen and phosph-orus following fertilization in two alpine tundra communities [J].Oikos,1994,70:261-270.
[31]Tessier J T,Raynal D J.Use of nitrogen to phosphorus ratios in plant tissue as an indicator of nutrient limitation and nitrogen saturation [J].Journal of Applied Ecology,2003,40:523-534.
[32]Güsewell S,Koerselman W,Verhoeven T A.Biomass N∶P ratios as indicators of nutrient limitation for plant populations in Wetlands [J].Ecological Applications,2004,13:372-384.
[33]Koerselman W,Meuleman A F M.The vegetation N∶P ratio:a new tool to detect the nature of nutrient limitation [J].Journal of Applied Ecology,1996,33:1441-1450.
[34]贺金生,韩兴国.生态化学计量学:探索从个体到生态系统的统一化理论 [J].植物生态学报,2010,34(1):2-6.
He J S,Han X G.Ecological stoichiometry:searching for unifying principles from individuals to ecosystems [J].Chinese Journal of Plant Ecology,2010,34(1):2-6.
Effect of drought stress and fertilization on accumulation and partitioning of biomass,C,N and P ofPueliasineseRoxb
ZHANG Mingjin,CHEN Lianghua,ZHANG Jian,HU Xiangwei,XU Rui
(KeyLaboratoryofEcologicalForestryEngineering,InstituteofEcology&Forestry,SichuanAgriculturalUniversity,Chengdu,Sichuan611130,China)
【Objective】 A controlled pot-culture experiment was conducted to investigate effects of drought stress and fertilization on accumulation and partitioning of biomass,C,N and P ofPueliasineseRoxb.【Method】 The experiment was a completely randomized design with 6 factorial combinations of 2 moisture gradients (drought,25% field capacity and control,75% field capacity) and 3 levels of fertilization (low,2 g/pot;moderate,4 g/pot;and high,8 g/pot).【Result】 Under controlled condition,with the increase of fertilization amount,the below-ground biomass,above-ground biomass and total biomass ofPueliasineseRoxb increased significantly.Under drought stress,the below-ground biomass and above-ground biomass ofPueliasineseRoxb decreased significantly,indicating that fertilization had significantly positive effect on the below-ground biomass,above-ground biomass and total biomass.Under controlled condition,the effects of fertilization on the allocation proportion of the below-ground and above-ground biomass were not significant.Under drought stress,both moderate and high fertilizations could significantly improve the allocation proportion of the below-ground biomass,except for low fertilization.Fertilization and drought stress also had certain influences on the accumulation and distribution of C,N and P.Compared with controlled condition,the accumulation quantities of C,N and P decreased after drought stress.Under controlled condition,the absorption efficiencies of N and P under low fertilization treatment were lower than moderate and high fertilizations,while the use efficiencies of N and P were higher under low fertilization treatment than moderate and high fertilizations.Under drought stress,the absorption efficiencies of N and P and the use efficiency of P among different fertilization treatments had no significant difference,but the use efficiency of N decreased with the increase of fertilization amount. Under controlled condition,fertilization had no significant effect on the C∶P and N∶P of above-ground part,but high fertilization significantly reduced the N∶P of underground part,while medium and high fertilizations significantly reduced the C∶N of above-ground part and the C∶P of underground part.Under drought stress,fertilization had no significant effect on the C∶P and N∶P of above-ground and underground part,but high fertilization significantly reduced the C∶N of above-ground and underground part.【Conclusion】 Drought stress decreased biomass,C,N and P accumulation,and affected the allocation of biomass,C,N and P.Fertilization could mitigate the effects of drought stress on plant growth and maintain a higher accumulation of biomass,C,N and P.More attention should be paid to the N fertilization management ofPueliasineseRoxb.
drought stress;fertilization;PueliasineseRoxb;biomass;nutrient accumulation and partitioning
时间:2016-08-0909:40DOI:10.13207/j.cnki.jnwafu.2016.09.014
2015-01-30
国家科技支撑计划项目“长江上游低山丘陵区生态综合整治技术及示范”(2011BAC09B05)
张明锦(1990-),女,四川芦山人,在读硕士,主要从事退化人工林生态恢复与重建研究。E-mail:958192283@qq.com
张健(1957-),男,重庆江津人,教授,主要从事森林生态、森林培育、水土保持研究。
E-mail:sicauzhangjian@163.com
S543
A
1671-9387(2016)09-0105-08
网络出版地址:http://www.cnki.net/kcms/detail/61.1390.S.20160809.0940.028.html
猜你喜欢
现代农村科技(2022年9期)2022-08-16
林业科技(2020年3期)2021-01-21
农民致富之友(2019年4期)2019-03-13
今日农业(2019年14期)2019-01-04
热带农业科学(2018年2期)2018-06-21
速读·中旬(2017年11期)2017-11-11
理论导刊(2017年8期)2017-08-30
作物研究(2014年6期)2014-03-01
植物营养与肥料学报(2012年5期)2012-10-26
植物营养与肥料学报(2012年1期)2012-10-26
