
添加前体物对虫草素产量的影响及动力学特征
2016-11-01 10:21:13闫瑞,周广麒
大连工业大学学报 2016年5期
闫 瑞, 周 广 麒
( 大连工业大学 生物工程学院, 辽宁 大连 116034 )
添加前体物对虫草素产量的影响及动力学特征
闫 瑞,周 广 麒
( 大连工业大学 生物工程学院, 辽宁 大连116034 )
研究腺苷等8种物质对CordycepsmilitarisFFCC 5111液体发酵生产虫草素的影响,并考察了其发酵动力学特征。结果表明,添加腺苷、苯丙氨酸、腺嘌呤、甘氨酸和L-谷氨酰胺均可提高虫草素产量,其中添加腺苷1.5 g/L的发酵液中虫草素产量最大,为772.5 mg/L,是对照组的9.39倍。在50 L发酵罐中,添加1.5 g/L腺苷的麦芽汁培养基进行分批发酵,建立动力学模型,其拟合值与实验值的相对误差小于8%,能良好地反映C.militarisFFCC 5111在补加腺苷的麦芽汁培养基中的发酵动力学特征。
蛹虫草;虫草素;腺苷;动力学
0 引 言
虫草素是蛹虫草产生的一种重要次级代谢产物,具有抗菌、抗癌和提高人体免疫力等药理功能[1]。采用液体深层发酵方法培养蛹虫草具有生产周期短、虫草素含量高等特点,是工业化生产虫草素的有效手段[2]。目前在液体深层培养过程中,许多研究者通过优化培养基组成及培养条件[3]、添加前体物质[4]、菌株诱变[5]等方法以提高虫草素产量,但使用的培养基多以化学试剂为主[6],不利于食品加工中的应用。目前国内尚无利用天然培养基通过优化补料方式提高虫草素产量方面的文献报道。本研究以蛹虫草菌株为研究对象,利用谷物原料发酵生产虫草素,保证了发酵产物在食品开发中的安全性[7]。通过液体发酵筛选生长因子,建立虫草素分批发酵动力学模型,对蛹虫草发酵生产虫草素的条件进行优化。
1 材料与方法
1.1材料
1.1.1菌种
CordycepsmilitarisFFCC 5111,由大连工业大学食品和发酵菌种培养中心提供。
1.1.2培养基及配方
种子培养基:马铃薯200 g/L,葡萄糖20 g/L,琼脂20 g/L。
基本培养基:12°麦芽汁培养基;制作方法:干燥麦芽→粉碎→加4倍体积水→52 ℃保温80 min→65 ℃保温糖化,至碘色反应不变色→煮沸→过滤→调糖度至12 °Bx。
1.2方法
1.2.1不同添加物对虫草素生成的影响
分别向装有100 mL基本培养基的三角瓶中添加0.5、1.0、1.5、2.0、2.5、3.0 g/L的腺嘌呤、腺苷、次黄嘌呤、苯丙氨酸、天冬氨酸、甘氨酸、甲硫氨酸及L-谷氨酰胺,并将蛹虫草种子液分别接种到不同添加物的基本培养基中,于25 ℃下培养20 d,测定虫草素含量。
1.2.2动力学实验
在50 L通风搅拌发酵罐中,将种子液接种到含有腺苷1.5 g/L的基本培养基中,装液量为60%,培养温度为25 ℃,搅拌转速为150 r/min,通气量为0.6 m3/h,培养20 d,每2 d 取样检测。
1.3检测方法
1.3.1菌体质量浓度
取一定量发酵液,4 500 r/min离心20 min,弃上清液,再加去离子水洗涤,离心2次,获沉淀放入干燥洁净已知质量的培养皿中,80 ℃烘干至恒重。
式中:ρ为菌体质量浓度,g/L;m为菌体干重,g;V为发酵液体积,mL。
1.3.2虫草素及腺苷含量
样品前处理:取一定量发酵液,4 500 r/min离心20 min,收集上清液,浓缩,并加入3倍体积95%乙醇,4 ℃静置10 h,再经11 000 r/min、4 ℃下离心15 min,取上清液,浓缩后定容。0.22 μm滤膜过滤,按设置好的色谱条件进样。
高效液相色谱条件:流动相A,磷酸盐溶液(20 mmol/L NaH2PO4和Na2HPO4,pH=6.89);流动相B,色谱纯-甲醇溶液;梯度洗脱。体积流量,0.8 mL/min;进样量,10 μL;柱温,30 ℃;检测波长,260 nm。
2 结果与讨论
2.1添加物对虫草素生成的影响
陈长兰等[8]在液体摇瓶培养中分别添加20种不同的氨基酸,证明所有氨基酸都有增加虫草素含量的作用,且不同菌株间存在差异。荆留萍等[9]和文庭池等[10]分别比较了不同核苷、碱基及氨基酸对虫草素产量的影响,分别发现腺苷或腺嘌呤为最佳添加物,最适添加量为4或8 g/L。可见因蛹虫草菌株的不同,添加物的种类和添加量对虫草素产量表现出不同的影响。本实验根据添加物化学组分,比较了8种化合物对生成虫草素的影响,结果见表1。
表1不同营养物及质量浓度对培养基中虫草素产量的影响
Tab.1Effectsofdifferentadditivesandconcentrationsoncordycepinproduction
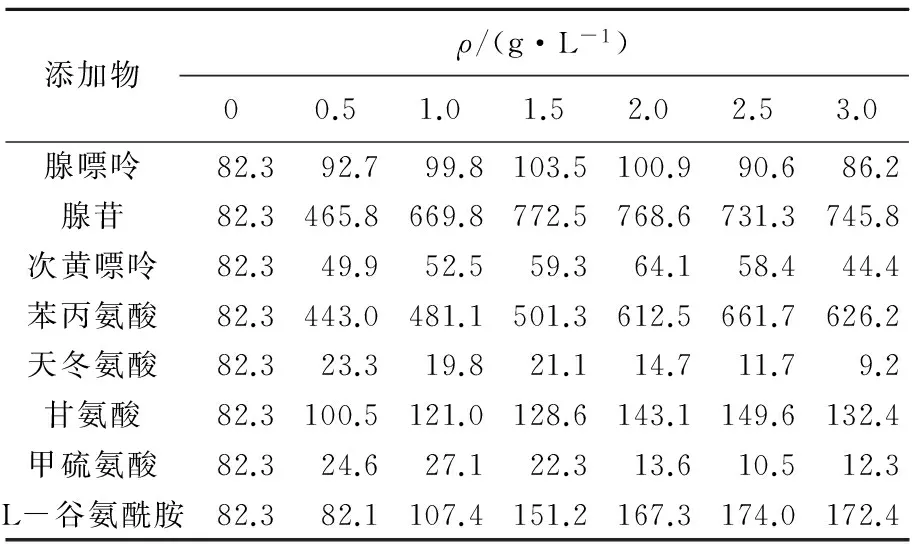
添加物ρ/(g·L-1)00.51.01.52.02.53.0腺嘌呤82.392.799.8103.5100.990.686.2腺苷82.3465.8669.8772.5768.6731.3745.8次黄嘌呤82.349.952.559.364.158.444.4苯丙氨酸82.3443.0481.1501.3612.5661.7626.2天冬氨酸82.323.319.821.114.711.79.2甘氨酸82.3100.5121.0128.6143.1149.6132.4甲硫氨酸82.324.627.122.313.610.512.3L-谷氨酰胺82.382.1107.4151.2167.3174.0172.4
由表1可见,添加苯丙氨酸、腺苷、腺嘌呤、甘氨酸和L-谷氨酰胺对虫草素产量均有促进作用,其中苯丙氨酸和腺苷效果最为明显。在各组中,添加苯丙氨酸和腺苷的实验组虫草素产量均高于其他组,其中腺苷添加量为1.5 g/L时,虫草素产量达到最大,是对照组的9.39倍。添加次黄嘌呤、天冬氨酸及甲硫氨酸的实验组结果低于对照组。除天冬氨酸外,均表现出随添加物质量浓度的增大,虫草素产量先升后降的趋势。腺嘌呤在各个质量浓度范围对虫草素产量均无显著影响。
2.2虫草素分批发酵过程动力学特征
2.2.1动力学特征
添加1.5 g/L腺苷的12 °Bx麦芽汁为培养基,在50 L发酵罐中进行分批发酵。根据分批发酵实验数据绘制发酵过程曲线,并得到每个时间间隔的菌体平均比生长速率、虫草素平均比生成速率和腺苷平均比消耗速率及它们的拟合曲线,分别如图1~3所示。比速率曲线拟合的结果均在95%置信区间内。
从图1可知,延滞期为0~2 d,对数生长期为2~12 d,稳定期在14 d之后;菌体比生长速率在2 d时达到最大,为0.542 d-1,之后随时间逐渐减小,当发酵进行到14 d时,比生长速率出现负值,基本保持不变,表示菌体量呈现负增长。
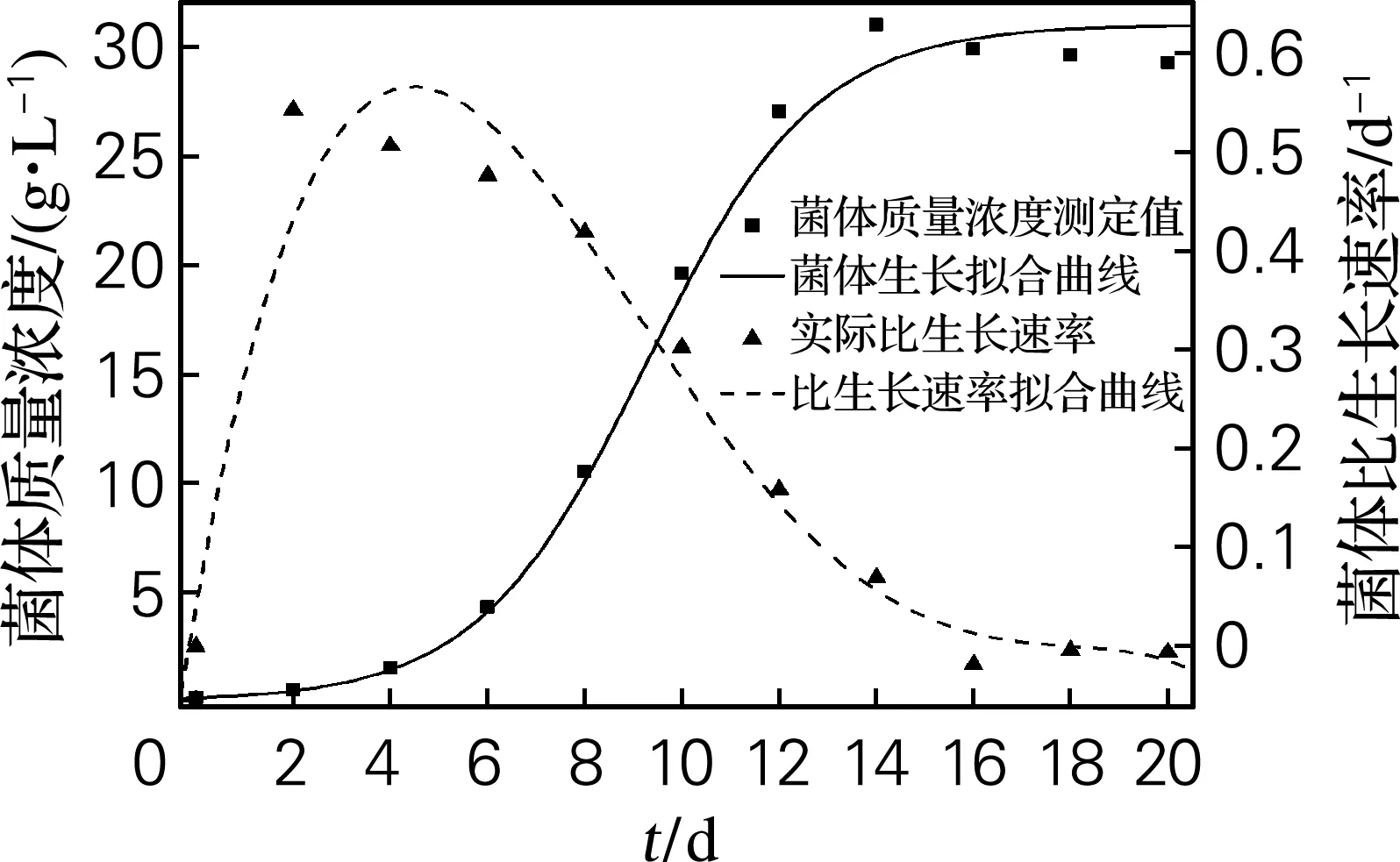
图1菌体生长动力学拟合曲线及比生长速率曲线
Fig.1ComparisonofkineticmodelwithbiomassandchangingofthegrowthrateduringsubmergedfermentationofC. militaris
由图2可以看出,在延滞期和对数生长中前期,虫草素生成很少,随着发酵时间的延长,生成速率迅速增大;虫草素比生成速率从发酵开始随时间上升,在10~20d达到稳定,比生成速率在1.75~2.02mg/(g·d);虫草素产量在20d时达到最大值625.1mg/L,但与摇瓶实验结果相比,下降了19.1%。
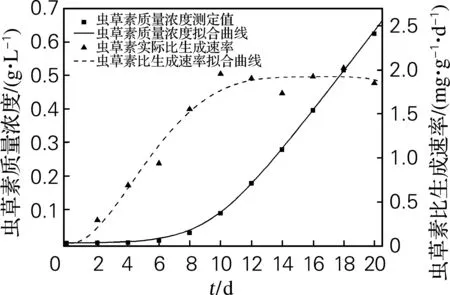
图2虫草素生成动力学拟合曲线及虫草素比生成速率曲线
Fig.2ComparisonofkineticmodelwithcordycepinproductionandchangingofthecordycepinproductionrateduringsubmergedfermentationofC. militaris
由图3可以看出,发酵液中腺苷质量浓度随发酵时间逐渐降低;腺苷比消耗速率在6d达到最大,之后随发酵时间不断下降,在14d趋于稳定,此时腺苷的产生和消耗达到动态平衡。
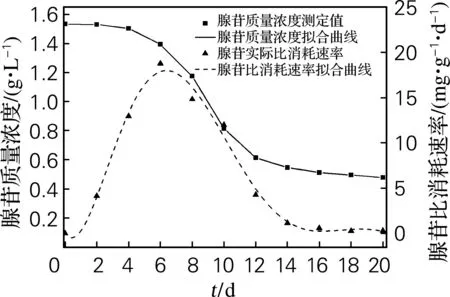
图3 腺苷质量浓度及腺苷比消耗速率曲线
Fig.3Changing of the adenosine concentration and consumption rate during submerged fermentation ofC.militaris
2.2.2菌体生长动力学模型
采用Logistic方程描述C. militarisFFCC5111菌体生长动力学模型[11],它是典型的S形曲线,能真实反映分批发酵过程中因菌体增加而抑制自身生长的作用,对拟合菌体生长过程具有广泛的适用性。Logistic方程为
(1)
对Logistic方程进行积分得,
(2)
式中:cx为菌体浓度;初始条件t=0时,cx=cx,0;cx,max为菌体的最大浓度;μmax为最大比生长速率;t为发酵时间。
以lncx-t作图,确定对数生长期为t=2~10 d。在对数生长期,以ln (cx/(cx,max-cx))-t作图,进行线性拟合,所得直线斜率即为μmax,得μmax=0.590 1,作为模型拟合的初始值。
2.2.3产物生成动力学模型
根据Luedeking-Piret方程
(3)
并由图1、2可知,C.militarisFFCC 5111虫草素的生成与菌体的生长无直接相关性,属于非生长偶联型[12]。因此,虫草素生成动力学模型可简化为
(4)
式中:cp为产物浓度;当t=0时,cp=cp,0;β为产物动力学模型参数,随发酵条件的变化而不同。
2.2.4动力学模型拟合及适用性
应用MATLAB软件的模型拟合工具包对实验数据进行非线性拟合。微分方程组采用四阶Runge-Kutta法求解,模型参数优化采用全局性收敛的经Levenberg-Marquart修正的高斯-牛顿法,根据实验数据和模型参数的初估值,以误差平方和最小为目标,获得全局性收敛的最优化参数估计值。求得分批发酵动力学模型参数μmax=0.574 4,β=1.863 4,拟合曲线见图1、2。
为验证模型的可靠性,根据所建立发酵动力学模型,可得分批发酵过程中菌体质量浓度及虫草素质量浓度随发酵时间变化的拟合值,进而计算出实测值与拟合值之间的相对误差,结果如表2所示。
表2菌体及虫草素质量浓度模型的拟合值与实验值对比
Tab.2Comparisonbetweenexperimentalandcalculateddataofcellandcordycepinconcentration
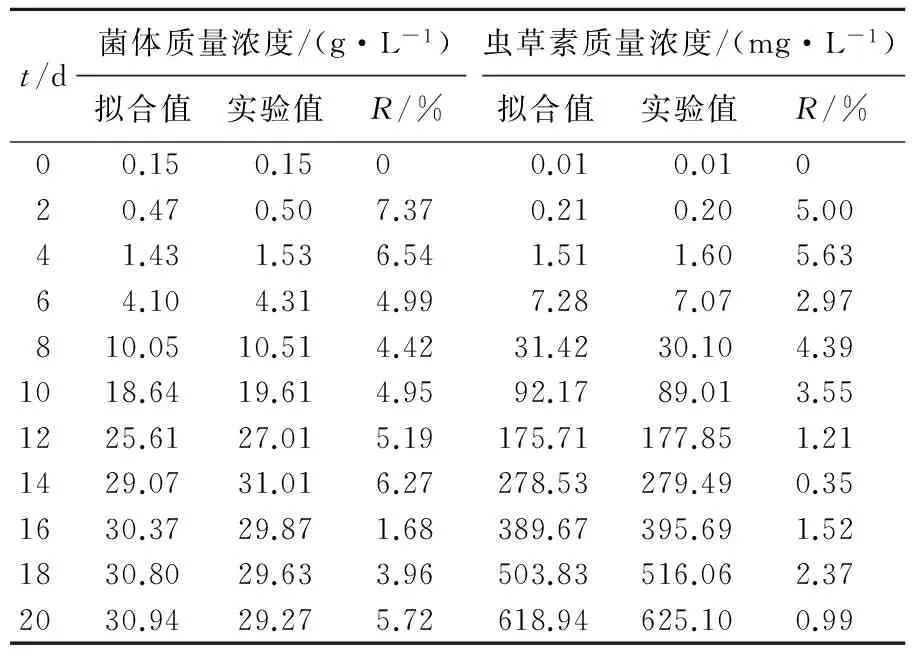
t/d菌体质量浓度/(g·L-1)虫草素质量浓度/(mg·L-1)拟合值实验值R/%拟合值实验值R/%00.150.1500.010.01020.470.507.370.210.205.0041.431.536.541.511.605.6364.104.314.997.287.072.97810.0510.514.4231.4230.104.391018.6419.614.9592.1789.013.551225.6127.015.19175.71177.851.211429.0731.016.27278.53279.490.351630.3729.871.68389.67395.691.521830.8029.633.96503.83516.062.372030.9429.275.72618.94625.100.99
由表2可知,除发酵初期各模型的误差值较高外,其他时期拟合值与实验值的误差相对较小,均在8%以内,说明所建数学模型能较好地反映C.militarisFFCC 5111在补加腺苷的天然培养基中的发酵动力学特征。
3 结 论
通过对8种供试前体及营养物的筛选,确定可有效提高C.militarisFFCC 5111胞外虫草素产量的5种物质,分别是苯丙氨酸、腺苷、腺嘌呤、甘氨酸和L-谷氨酰胺,其中添加1.5 g/L腺苷,虫草素产量达到最大,为772.5 mg/L。根据对实验结果及腺苷化学结构的分析,可以看出腺苷与虫草素化学结构十分相似,能迅速被蛹虫草吸收和转化。
对C.militarisFFCC 5111分批发酵动力学特征进行分析,确定虫草素生成与菌体生长呈非偶联型,并采用Logistic方程、Luedeking-Piret方程建立虫草素分批发酵动力学模型,所建模型能较好地映分批发酵过程中菌体生长及产物生成随时间变化的关系。
[1] 秦建春,李晓明,张鞍灵,等.蛹虫草发酵液抗菌活性初步研究[J].西北植物学报,2006,26(2):402-406.
[2] KANG C, WEN T C, KANG J C, et al. Effects of additives and different culture conditions on cordycepin production by the medicinal fungusCordycepsmilitaris[J]. Mycosystema, 2012, 31(3): 389-397.
[3] 岳翠翠,沈健增,蔡宇杰,等.蛹虫草CordycepsmilitarisJN168产虫草素液态发酵条件的优化[J].食品与生物技术学报,2013,32(2):135-141.
[4] 王蕾,罗巍,胡瑕,等.虫草素高产菌株的筛选及不同添加物对虫草素产量的影响研究[J].菌物学报,2012,31(3):382-388.
[5] MASUDA M, DAS S K, FUJIHARA S, et al. Production of cordycepin by a repeated batch culture of aCordycepsmilitarismutant obtained by proton beam irradiation[J]. Journal of Bioscience and Bioengineering, 2011, 111(1): 55-60.
[6] 尹萍,涂艳丽,王飞,等.北虫草液体发酵培养基优化研究[J].江西农业学报,2006,18(4):102-103.
[7] 解春艳,顾振新,顾颖娟,等.北虫草液态发酵天然培养基的优化研究[J].食品工业科技,2012,33(4):223-226.
[8] 陈长兰,孟程程,佟丽,等.不同种类氨基酸对蛹虫草菌丝体生长和虫草素含量的影响[J].食品科学,2012,33(23):236-239.
[9] 荆留萍,杜双田,金凌云,等.8种物质对蛹虫草液体发酵中虫草素及多糖含量的影响[J].西北农林科技大学学报(自然科学版),2010,38(11):156-160.
[10] 文庭池,康冀川,雷帮星,等.前体及营养物提高蛹虫草虫草菌素产量的研究[J].食品科学,2010,31(5):175-179.
[11] 刘霞,白占涛,杨亮,等.甘草内生菌Z4菌株的分批发酵动力学研究[J].西北农林科技大学学报(自然科学版),2013,41(9):164-168.
[12] 于田,王际辉,庞宇辰,等.嗜酸乳杆菌和嗜热链球菌混菌发酵合成共轭亚油酸的动力学[J].大连工业大学学报,2013,32(1):11-14.
Effects of additives on cordycepin production and kinetics models of batch fermentation byCordycepsmilitaris
YANRui,ZHOUGuangqi
( School of Biological Engineering, Dalian Polytechnic University, Dalian 116034, China )
TheeffectsofdifferentadditivesoncordycepinproductionbysubmergedcultureofCordyceps militarisFFCC5111wereinvestigated.Theresultindicatedthattheadditivesofadenosine,phenylalanine,adenine,glycineandL-glutaminecouldresultinalargeimprovementincordycepinproductionbysubmergedfermentationofC. militaris,especiallyadding1.5g/Ladenosine,whichcouldresultinthehighestcordycepinproductionof772.5mg/L,exhibitinga9.39-foldincreasethanthatobtainedinthebasalmediumculture.Adding1.5g/Ladenosineasamediumina50Lbatchsystem,thekineticmodelforC. militarisFFCC5111wasbasedontheLogisticandLuedeking-Piretequationsforcellgrowthandproductformation.Therelativeerrorbetweentheexperimentalvalueandfittedvaluewaslessthan8%,indicatingthatthemodelcouldreflectthekineticsfeatureofbatchfermentationofC. militarisFFCC5111inmaltosemediumsupplementedwithadenosine.
Cordycepmilitaris; cordycepin; adenosine; kinetics
2015-03-04.
辽宁省高校创新团队支持计划项目(LN2012011).
闫 瑞(1989-),男,硕士研究生;通信作者:周广麒(1957-),男,教授.
TS218;Q939.97
A
1674-1404(2016)05-0321-04
闫瑞,周广麒.添加前体物对虫草素产量的影响及动力学特征[J].大连工业大学学报,2016,35(5):321-324.
YAN Rui, ZHOU Guangqi. Effects of additives on cordycepin production and kinetics models of batch fermentation byCordycepsmilitaris[J]. Journal of Dalian Polytechnic University, 2016, 35(5): 321-324.
猜你喜欢
中老年保健(2022年2期)2022-08-24 03:20:44
当代水产(2022年1期)2022-04-26 14:35:38
检察风云(2022年5期)2022-04-05 13:42:39
中老年保健(2021年12期)2021-11-30 02:58:01
黑龙江大学自然科学学报(2021年4期)2021-11-19 07:05:02
中国调味品(2017年2期)2017-03-20 16:18:21
现代检验医学杂志(2016年3期)2016-11-15 01:59:48
国外医药(抗生素分册)(2016年3期)2016-07-12 14:25:18
中央民族大学学报(自然科学版)(2016年3期)2016-06-27 07:55:28
山东医药(2015年16期)2016-01-12 00:40:08
