
重金属及盐胁迫对绿金合果芋生理特性的影响
2016-10-31 04:35张家洋
浙江农业学报 2016年4期
张家洋
(新乡学院 生命科学技术学院,河南 新乡 453003)
重金属及盐胁迫对绿金合果芋生理特性的影响
张家洋
(新乡学院 生命科学技术学院,河南 新乡 453003)
以绿金合果芋(SyngoniumpodophyllumSchott)为研究对象,研究了重金属离子(Pb2+,Cd2+)、盐(NaCl)及其复合溶液对其光合系统、膜系统及抗氧化酶系统的影响。结果表明: 随着重金属及盐处理浓度的增加,绿金合果芋叶片内叶绿素、类胡萝卜素、可溶性蛋白、可溶性糖含量和POD活性均受到抑制;游离脯氨酸含量、CAT活性具应激效应,即在低浓度胁迫下升高,高浓度下降低;丙二醛含量和相对电导率均随胁迫浓度的增加呈上升趋势;SOD活性随重金属处理浓度的增加而降低,但在NaCl处理下则表现为低浓度下升高,高浓度下降低。通过本研究发现,绿金合果芋的光合系统、膜系统和抗氧化酶活性等指标对重金属胁迫的反应均较为灵敏,可作为评估重金属及盐胁迫对其危害的指标。
重金属;盐胁迫; 绿金合果芋; 生理特性
伴随着现代农业、工业、交通运输业等人类活动以及城市化进程的加快,土壤重金属污染及复合污染问题变得日趋严重,对植物的毒害作用逐渐显现,城市园林植物也难逃其害。目前,关于重金属污染对植物的影响研究多集中在农作物、经济作物,但近几年国内外学者对植物修复重金属污染技术方面的研究证实,观赏植物不仅具有美化环境的功能,而且具有修复重金属污染的作用[1]。
绿金合果芋(SyngoniumpodophyllumSchott),又名箭叶芋,为天南星科多年生常绿攀缘草本植物,原产于中南美洲热带地区,全世界广泛栽培,我国南方各省种植十分普遍;它除了广泛应用于室内盆栽观赏外,还可设立成绿色支柱造型,更多用于室外半阴处作地被覆盖,是一个极具发展潜力的观叶植物[2]。对于绿金合果芋的研究多集中在其繁殖方式和栽培技术等方面[3-4],关于其抗重金属及盐的研究报道尚少,尤其是重金属及盐胁迫对其伤害及植物的抗性机制更少。为此,本研究采用不同浓度的NaCl溶液和Pb(NO3)2,CdCl2及复合溶液对绿金合果芋进行处理,研究重金属及盐胁迫对其生长、生理和抗氧化酶指标的影响,通过分析绿金合果芋在Pb2+,Cd2+,NaCl及其交互胁迫下各生理指标的响应情况,得出可作为抗重金属性能及耐盐评判的指标,进而得到绿金合果芋的抗重金属污染及土壤盐碱化的能力,为植物修复铅镉污染土壤和治理盐碱地开辟新的方法途径和提供科学依据,同时获得最大生态效应和美观效应。
1 材料与方法
1.1供试材料与处理
供试植物为天南星科,绿金合果芋属,绿金合果芋,由花卉市场统一采购,选取生长健壮,无病虫害,高度、大小、长势一致的植株。试验采用盆栽法。供试土壤为腐殖土和园土按质量比 2∶1均匀混合,以保证每盆土壤的均一性,在陶瓷盆装入风干的供试土壤,每盆25 kg。试验用陶瓷盆的上直径为65 cm,底直径 55 cm,高 45 cm。
将Pb(NO3)2,CdCl2·H2O 分析纯试剂折合成每盆土中Pb,Cd的施入量, 试验设4种处理(表1)。处理方法为:以不加Pb,Cd、NaCl为对照(CK),设NaCl1,NaCl2,NaCl3,NaCl4,Pb1,Pb2,Pb3,Pb4,Cd1,Cd2,Cd3,Cd4,Pb1+ Cd1,Pb2+Cd2,Pb3+Cd3,Pb4+Cd4,NaCl1+Pb1+ Cd1,NaCl2+ Pb2+Cd2,NaCl3+ Pb3+Cd3,NaCl4+Pb4+Cd4共20个处理,将试验苗木移载到盆中,每盆1株,每个处理栽植6盆,待苗木恢复生长(10 d)后,按设计质量浓度以水溶液的形式对植物进行统一浇灌;所有的处理及处理后苗木的生长均在育种大棚内进行,且每次用等量的水浇灌养护,除草、松土等管理措施均相同,以避免因受雨水或其他管理措施的影响而造成植株间处理浓度的差异。持续处理30 d后测定各项生理指标,每株以第4片功能叶为采样叶,距叶轴1/2半径处避开叶脉采样,每个样品重复3次。
1.2各项生理指标的测定方法
分别于处理后30 d测定每盆植物的9项生理指标:丙酮提取法检测叶绿素和类胡萝卜素[5];用蒽酮法测定可溶性糖(620 nm)[6];考马斯亮蓝染色法测定可溶性蛋白含量(595 nm)[6];酸性茚三酮法检测游离脯氨酸含量(520 nm)[7];愈创木酚法测定过氧化物酶(POD)活性(470 nm),以1 minD值变化(升高)0.01为1个酶活性单位(U)[5];氮蓝四唑光化还原法测定超氧化物歧化酶(SOD)活性(560 nm)[6],SOD活性单位以抑制NBT光化还原50%所需酶量为1个酶活单位(U);紫外吸收法测定过氧化氢酶(CAT)活性(240 nm),以1 minD值减少0.01为1个酶活性单位(U)[6];硫代巴比妥酸法检测丙二醛含量(532,600和450 nm)[8];相对电导率测定方法参照李合生方法[6]。
表1重金属及盐胁迫处理浓度
Table 1Concentrations of heavy metals and salt in the experiment
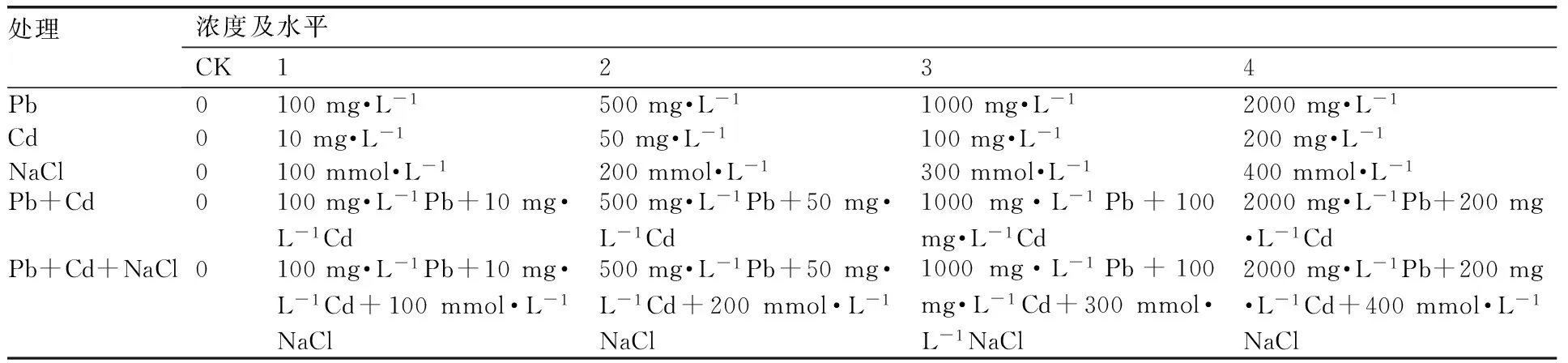
处理浓度及水平CK1234Pb0100mg·L-1500mg·L-11000mg·L-12000mg·L-1Cd010mg·L-150mg·L-1100mg·L-1200mg·L-1NaCl0100mmol·L-1200mmol·L-1300mmol·L-1400mmol·L-1Pb+Cd0100mg·L-1Pb+10mg·L-1Cd500mg·L-1Pb+50mg·L-1Cd1000mg·L-1Pb+100mg·L-1Cd2000mg·L-1Pb+200mg·L-1CdPb+Cd+NaCl0100mg·L-1Pb+10mg·L-1Cd+100mmol·L-1NaCl500mg·L-1Pb+50mg·L-1Cd+200mmol·L-1NaCl1000mg·L-1Pb+100mg·L-1Cd+300mmol·L-1NaCl2000mg·L-1Pb+200mg·L-1Cd+400mmol·L-1NaCl
1.3数据处理
所有的试验数据采用Excel 2007 和SSPS13.0 软件进行统计和分析。
2 结果与分析
2.1盐和重金属胁迫对绿金合果芋光合色素含量的影响
叶绿素是植物光合作用的主要色素,其含量高低直接反映植物光合作用能力的强弱。类胡萝卜素除参加光合作用以外,还具有清除活性氧的作用。多种逆境胁迫作用下可导致二者含量的下降[9]。不同盐及重金属胁迫下对绿金合果芋叶片光合色素的影响见表2。结果显示:在短期单一胁迫下,叶绿素a、叶绿素b、类胡萝卜素含量均呈现下降的趋势,均在重度下达到最小值,其中, NaCl4,Pb4,Cd4胁迫下叶绿素a分别为对照的31.77%,35.87%,34.32%;NaCl4,Pb4,Cd4胁迫下叶绿素b分别为对照的35.62%,40.83%,41.99%;NaCl4,Pb4,Cd4胁迫下类胡萝卜素分别为对照的67.85%,46.71%,45.43%。表明中度、重度重金属及盐胁迫处理对绿金合果芋叶片光合色素的合成具有一定的破坏作用,其中叶绿素a对胁迫处理更为敏感。
重金属及盐复合胁迫下,绿金合果芋叶片叶绿素a、叶绿素b和类胡萝卜素含量随浓度的增加快速降低,均在重度胁迫下达到最小值。Pb4+Cd4处理组绿金合果芋叶片叶绿素a、叶绿素b和类胡萝卜素含量分别为对照的13.16%,30.21%和54.77%;NaCl4+Pb4+Cd4处理组绿金合果芋叶片叶绿素a、叶绿素b和类胡萝卜素含量分别为对照的10.42%,27.51%和44.64%,和单一胁迫一样,叶绿素a对复合胁迫处理表现得更敏感。
2.2盐和重金属胁迫对绿金合果芋可溶性糖含量的影响
可溶性糖可以对很多非盐植物进行渗透调节,有稳定细胞膜和原生质胶体的作用,在渗透胁迫作用下植物体内可溶性糖的积累可有效地降低渗透势,以达到适应外界环境变化的作用[10]。
表2盐及重金属胁迫处理对绿金合果芋光合色素含量的影响
Table 2Effects of salt and heavy metals stress on chlorophyll content ofSyngoniumpodophyllumSchott
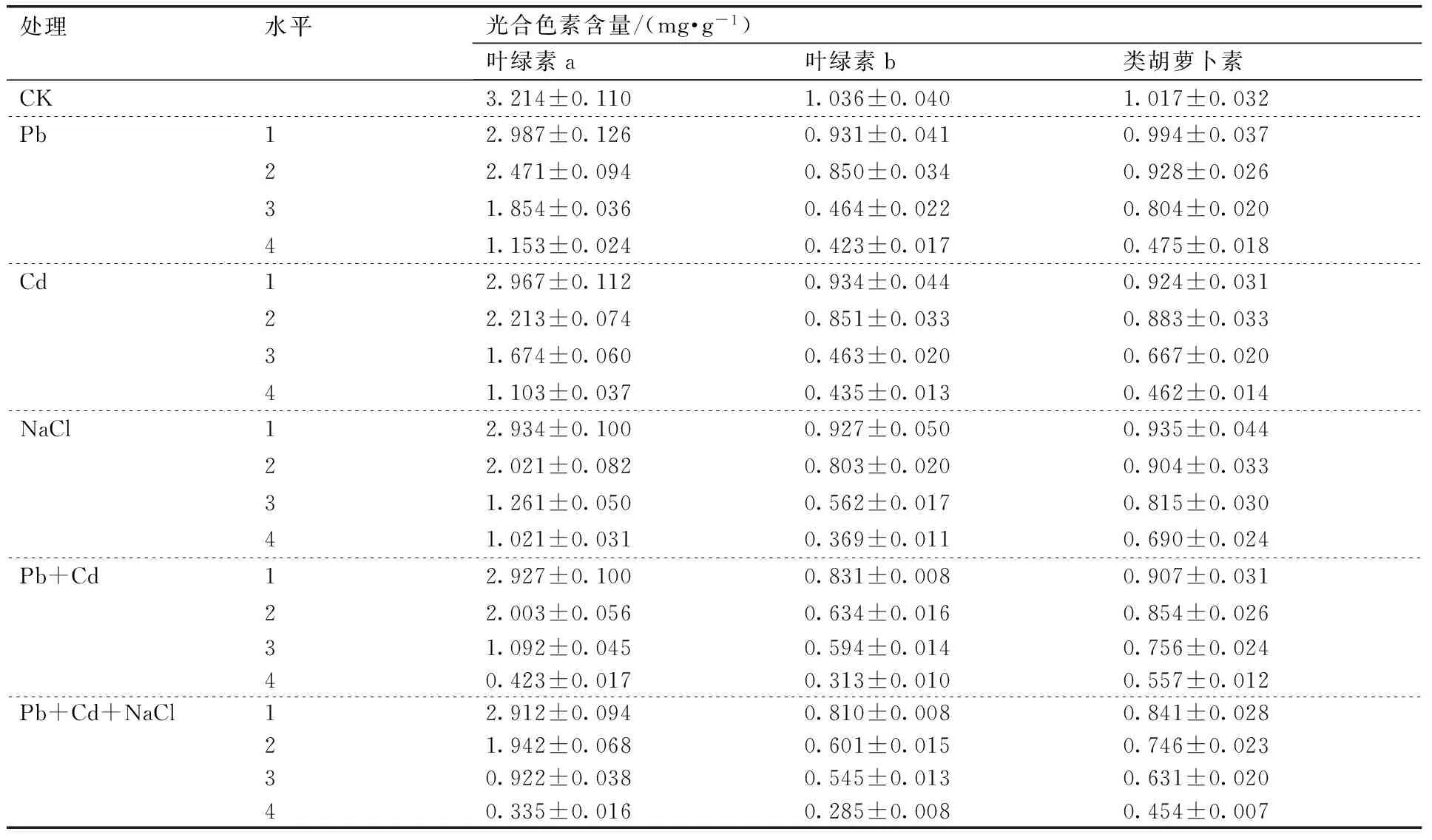
处理水平光合色素含量/(mg·g-1)叶绿素a叶绿素b类胡萝卜素CK3.214±0.1101.036±0.0401.017±0.032Pb12.987±0.1260.931±0.0410.994±0.03722.471±0.0940.850±0.0340.928±0.02631.854±0.0360.464±0.0220.804±0.02041.153±0.0240.423±0.0170.475±0.018Cd12.967±0.1120.934±0.0440.924±0.03122.213±0.0740.851±0.0330.883±0.03331.674±0.0600.463±0.0200.667±0.02041.103±0.0370.435±0.0130.462±0.014NaCl12.934±0.1000.927±0.0500.935±0.04422.021±0.0820.803±0.0200.904±0.03331.261±0.0500.562±0.0170.815±0.03041.021±0.0310.369±0.0110.690±0.024Pb+Cd12.927±0.1000.831±0.0080.907±0.03122.003±0.0560.634±0.0160.854±0.02631.092±0.0450.594±0.0140.756±0.02440.423±0.0170.313±0.0100.557±0.012Pb+Cd+NaCl12.912±0.0940.810±0.0080.841±0.02821.942±0.0680.601±0.0150.746±0.02330.922±0.0380.545±0.0130.631±0.02040.335±0.0160.285±0.0080.454±0.007
从图1中可以看出:随着处理浓度的增加,绿金合果芋叶片可溶性糖含量有所下降,在轻中度污染(处理水平1和2)胁迫下,变化并不明显,但当达到重度污染(处理水平3和4)时降幅较大,较为明显的是在重金属处理水平4处,其中Pb4处理下其可溶性糖的含量是对照的29.37%,Cd4处理下其可溶性糖的含量是对照的20.06%,NaCl4处理下其可溶性糖含量是对照的33.87%。
重金属及盐复合胁迫下,绿金合果芋叶片可溶性糖含量随着处理浓度的增加而急剧下降,均在处理水平4处达到最小值,其中Pb4+Cd4处理组可溶性糖含量仅为对照的16.85%,NaCl4+Pb4+Cd4处理组可溶性糖含量的降幅更大,是对照的7.22%。
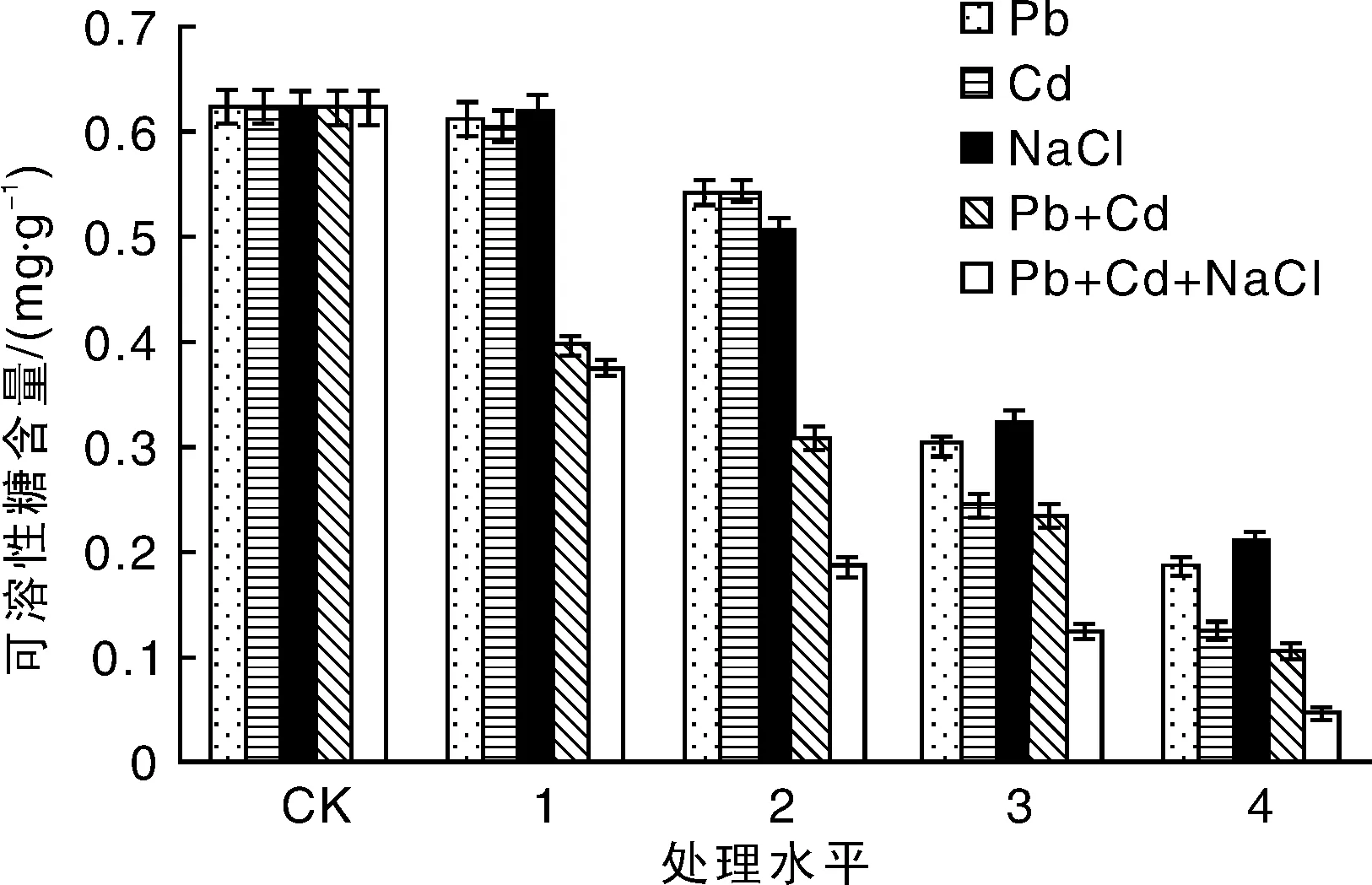
图1 盐及重金属胁迫处理对绿金合果芋可溶性糖含量的影响Fig.1 Effects of salt and heavy metals stress on soluble sugar content of Syngonium podophyllum Schott
2.3盐和重金属胁迫对绿金合果芋可溶性蛋白含量的影响
可溶性蛋白含量变化是反映植物叶片功能及衰老的重要指标之一,叶片中的可溶性蛋白的含量与植物生长代谢活动呈正相关,可溶性蛋白是植物相对抗性的一种指标[11]。从图2中可以看出,与可溶性糖变化情况类似,随着胁迫浓度的增加,重金属及盐单独处理组绿金合果芋叶片可溶性蛋白含量有所下降,在轻度污染(处理水平1)下略微降低,但当达到中度和重度污染(处理水平2~4)时降幅十分明显,即Pb4处理下其可溶性蛋白的含量是对照的10.46%,Cd4处理下其可溶性蛋白的含量是对照的8.35% ,NaCl4处理下其可溶性蛋白的含量是对照的15.00%。
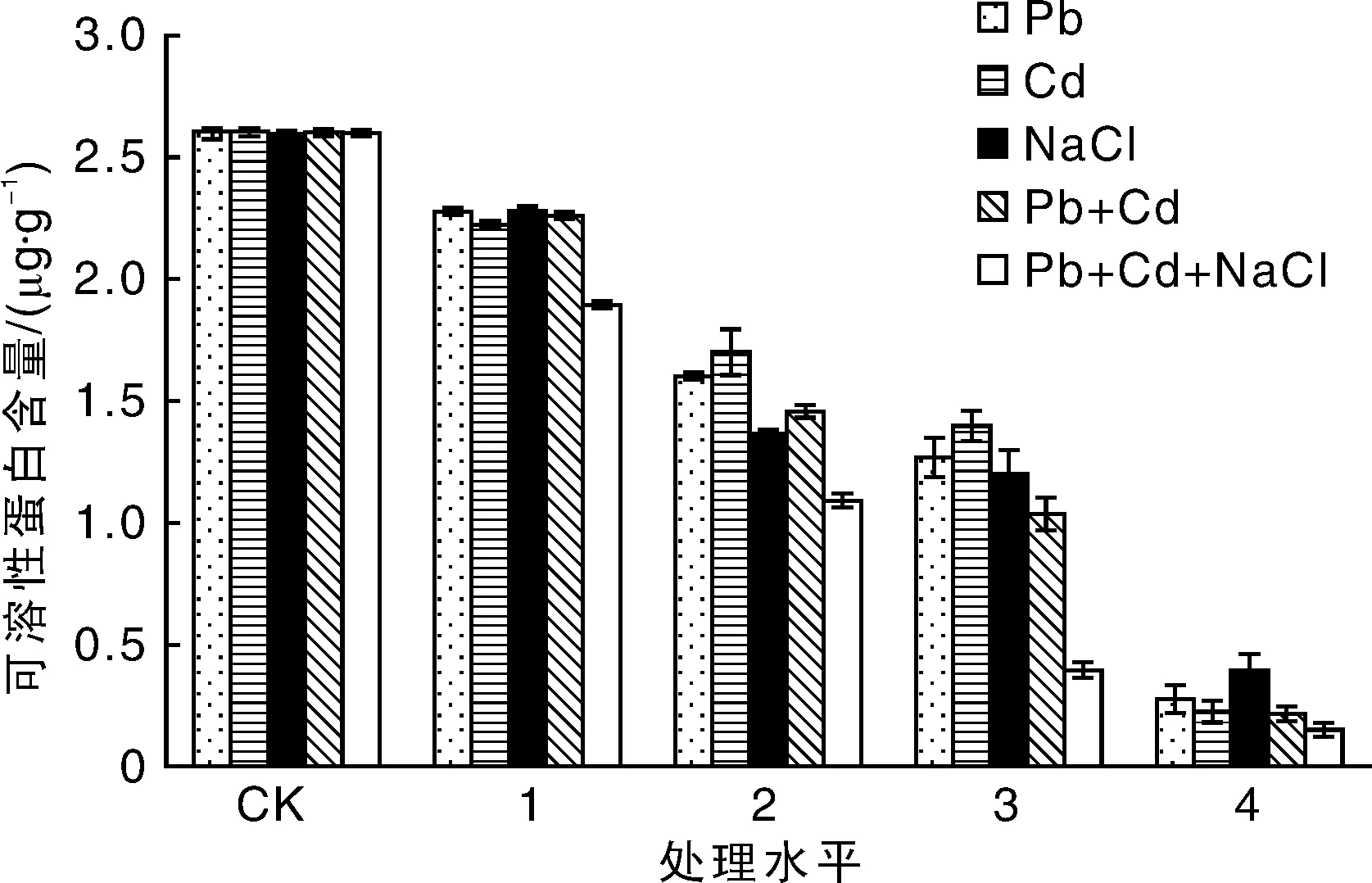
图2 盐及重金属胁迫处理对绿金合果芋可溶性蛋白含量的影响Fig.2 Effects of salt and heavy metals stress on soluble protein content of Syngonium podophyllum Schott
复合胁迫下,绿金合果芋叶片可溶性蛋白的含量随着处理浓度的增加而急剧下降,其最小值出现在处理水平4,Pb4+Cd4处理组和NaCl4+Pb4+Cd4处理组可溶性蛋白的含量分别是对照的8.08%和5.73%。
2.4盐和重金属胁迫对绿金合果芋游离脯氨酸含量的影响
除了可溶性糖和可溶性蛋白,游离脯氨酸是植物体另外一种重要和有效的有机渗透调节物质,逆境胁迫下会引起植物体内脯氨酸的积累,从而增强植物对环境的适应性。在逆境胁迫下,植物启动防御机制,通过脯氨酸的积累来调节植物细胞渗透势,降低伤害程度,保护原生质,减轻质膜的伤害程度[12]。不同重金属及盐胁迫下绿金合果芋叶片中游离脯氨酸含量的变化见图3。
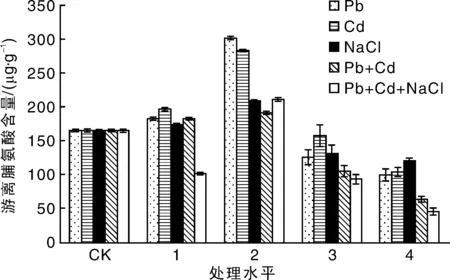
图3 盐及重金属胁迫处理对绿金合果芋游离脯氨酸含量的影响Fig.3 Effects of salt and heavy metals stress on free proline content of Syngonium podophyllum Schott
在Pb,Cd和NaCl单一短期胁迫下,绿金合果芋叶片游离脯氨酸含量均呈现先上升后下降的趋势,均在处理水平2处达到最大值, Pb2,Cd2和NaCl2分别是对照的1.88倍、1.76倍和1.33倍;在处理水平4下达到最小值,NaCl4,Pb4和Cd4分别是对照的60%,63.03%和72.73%。
在复合胁迫下,绿金合果芋叶片游离脯氨酸含量均呈现先急速上升后下降的趋势,均在处理水平2处达到最大值,Pb2+Cd2处理组和NaCl2+Pb2+Cd2处理组分别是对照的1.47倍和1.53倍;在处理水平4下达到最小值,Pb4+Cd4处理组和NaCl4+Pb4+Cd4处理组分别是对照的38.18%和27.88%。
2.5盐和重金属胁迫对绿金合果芋SOD活性的影响
SOD作为植物抗氧化保护系统的第一道防线,主要功能是清除活性氧,是防护氧自由基对细胞膜伤害的一种重要保护酶,在适度的条件下,提高活性以增加植物的抗逆能力[13]。图4-A所示,在Pb,Cd单一胁迫下,绿金合果芋叶的SOD活性随着浓度的升高逐渐降低,均在处理水平4处(Pb 2 000 mg·L-1和Cd 200 mg·L-1)SOD活性最低,分别为对照的63.23%和61.84%;在NaCl单一胁迫下,绿金合果芋叶的SOD活性随着盐浓度的升高先增加后降低,200 mmol·L-1处理时SOD活性最高,为对照的118.23%,之后逐渐减弱,并都低于对照,NaCl浓度为400 mmol·L-1时活性最低,为对照的76.50%。所以NaCl胁迫强度不同,其酶活性水平也不相同。低浓度的NaCl胁迫,具有较强的消除氧自由基的功能,但在较高浓度胁迫下却表现酶活性降低,并低于对照,说明绿金合果芋通过自我调节SOD活性来适应盐胁迫的能力是有限的。
重金属与盐复合胁迫条件下,SOD活性随着处理浓度的增加与对照相比呈现显著降低趋势,在处理水平4处达到最低值,其中Pb4+Cd4处理组SOD含量仅为对照的42.22%,NaCl4+Pb4+Cd4处理组降幅更大,SOD含量仅为对照的23.35%,说明随着复合胁迫程度的加剧,绿金合果芋的生长受到了显著的抑制。
2.6盐和重金属胁迫对绿金合果芋POD活性的影响
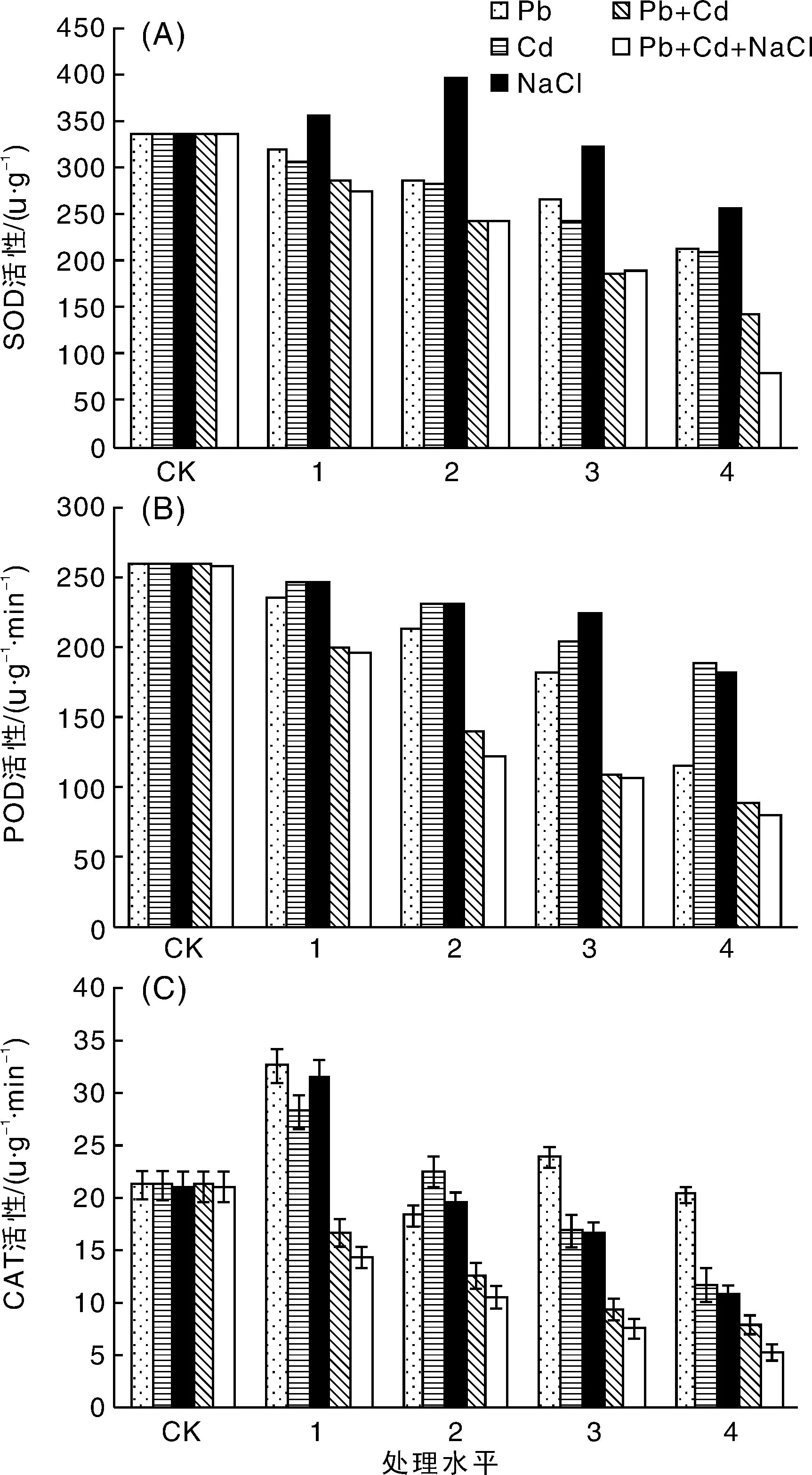
图4 盐及重金属胁迫处理对绿金合果芋SOD(A),POD(B)和CAT(C)活性的影响Fig.4 Effects of salt and heavy metals stress on SOD(A), POD (B)and CAT(C) activity of Syngonium podophyllum Schott
POD是植物体内的另外一种保护酶,有较高的活性,它的作用是清除胞浆或线粒体中产生的低浓度的H2O2,提高植物的抗逆能力,是一种对环境因子十分敏感的酶[13]。由图4-B所示,在重金属Pb的胁迫下,随着浓度的增加其活性逐渐降低,且均小于对照,在重度胁迫(Pb4)下达到最小值,为对照的44.43%;在重金属Cd的胁迫下,随着浓度的增加和重金属Pb表现为同样的趋势,这一结果表明在重度重金属胁迫(处理水平4)下绿金合果芋的生理机能受到了显著的抑制;在NaCl单一胁迫下,绿金合果芋POD活性随着盐浓度的升高逐渐降低,在100 mmol·L-1处理时SOD活性最高。
重金属与盐复合胁迫条件下,与SOD活性表达趋势一样,POD活性也是随着处理浓度的增加与对照相比呈现显著降低趋势,在处理水平4处达到最低值, Pb4+Cd4处理组和NaCl4+Pb4+Cd4处理组POD含量分别为对照的33.98%和30.74%,说明随着复合胁迫程度的加剧,绿金合果芋的生长受到了显著的抑制。
2.7盐和重金属胁迫对绿金合果芋CAT活性的影响
植物在逆境条件下能否正常生长,CAT是酶促防御系统的关键酶之一,它与SOD和POD协同配合,清除体内过剩的自由基,提高了植物的抗逆能力[13]。由图4-C所示,在单一的Pb,Cd和NaCl下,随着浓度的升高CAT活性均先升高再降低,在处理水平1处达到最大值,分别是对照的1.55,1.34和1.50倍。即在低浓度时,绿金合果芋叶片CAT活性增加,显示了其清除氧自由基能力增强,但在较高浓度时,其活性开始下降,说明其抗性保护能力是有一定限度的。
复合胁迫下,CAT活性随着处理浓度的增加而逐渐降低,在处理水平4时,活性最低,其中Pb4+Cd4复合胁迫为对照的36.75%,NaCl4+Pb4+Cd4复合胁迫为对照的24.93%。说明其清除自由基的能力在浓度较低时就表现出抑制性,随着浓度的增加抑制性就更加明显。
2.8盐和重金属胁迫对绿金合果芋MDA含量的影响
MDA含量表示的是植物膜脂的过氧化水平,反映植物受伤害的程度。植物叶片内MDA含量越多,说明受损害程度越大[14]。关于植物在盐碱及重金属胁迫处理下MDA含量的增加报道很多,如沈高峰[15]在高羊茅、张永峰等[16]在紫花苜蓿上的研究,本试验中随着重金属和盐浓度的增大,绿金合果芋叶片的MDA含量均呈现先降后升的趋势,即随着胁迫浓度的增加,绿金合果芋受到的胁迫伤害也加大。无论是单一胁迫还是复合胁迫,在轻度重金属和盐胁迫下,叶片MDA含量均略低于对照,而在中度和重度胁迫下,叶片MDA含量明显高于对照,这说明绿金合果芋对中度和重度重金属及盐胁迫更敏感。其中,增幅最为明显的是在重度复合胁迫处理组,Pb4+Cd4处理水平处其MDA含量是对照的2.74倍,NaCl4+Pb4+Cd4处理水平处是对照的4.53倍(图5)。
2.9盐和铅胁迫对绿金合果芋相对电导率的影响
生物膜结构和功能的稳定性与植物的抗逆性密切相关。重金属及盐胁迫对细胞膜造成了伤害,导致渗透物质大量外流,引起相对电导率升高。图6所示:在胁迫下,绿金合果芋叶片的相对电导率呈现上升的趋势,这与秦锋梅等[17]在黄菖蒲,熊春晖等[18]在莲藕,李学强等[12]在欧李,原海燕等[11]在喜盐鸢尾上的研究结论相似,即从轻度胁迫到重度胁迫,绿金合果芋叶片的相对电导率逐渐上升,均在处理水平4处达到最大值。即在处理30 d时,NaCl4,Pb4,Cd4,Pb4+Cd4,NaCl4+ Pb4+Cd4处理组的相对电导率分别是对照的1.46倍、1.42倍、1.34倍、1.55倍和1.74倍。
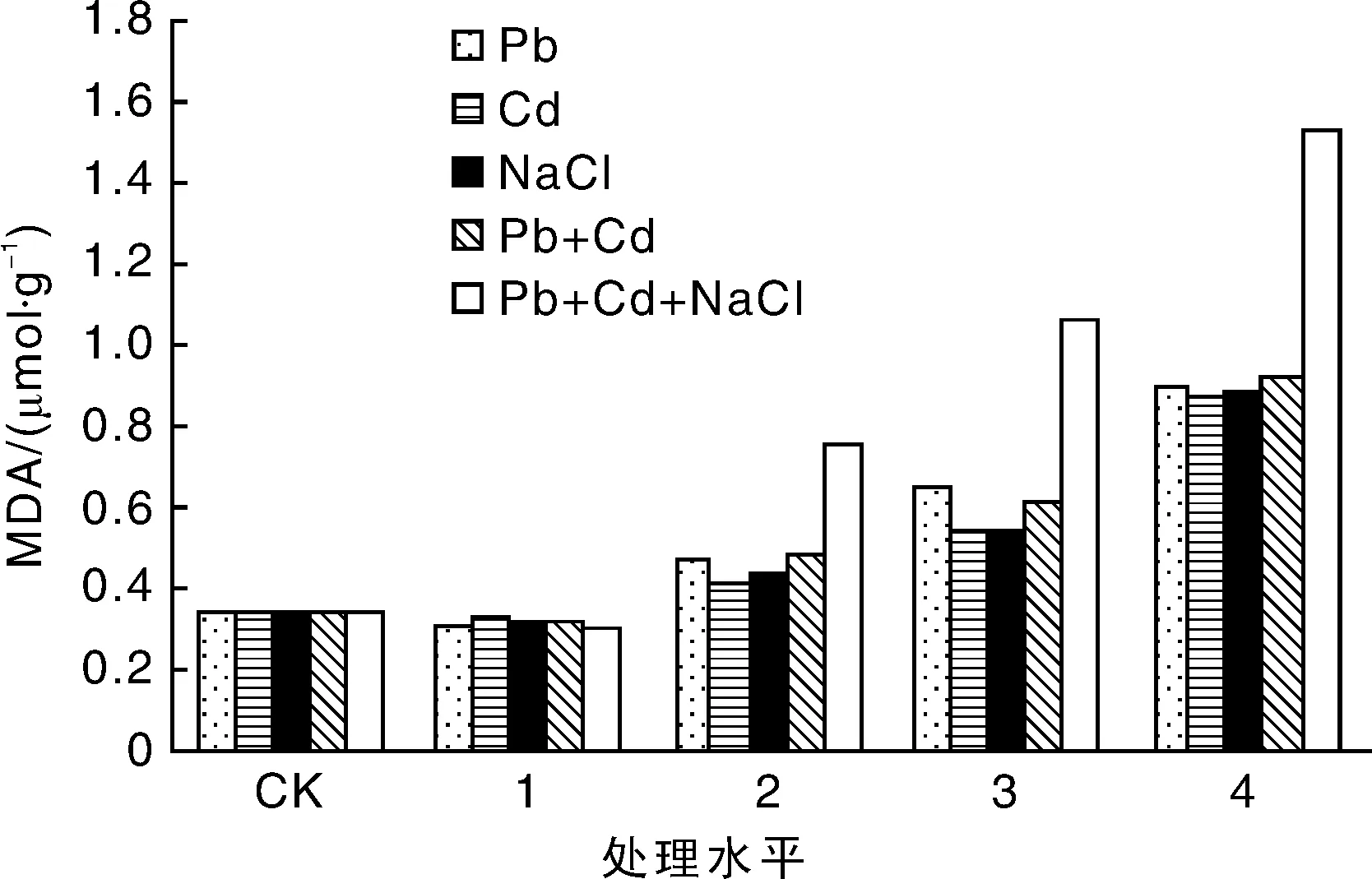
图5 盐及重金属胁迫处理对绿金合果芋丙二醛含量的影响Fig.5 Effects of salt and heavy metals stress treatment on malondialdehyde content of Syngonium podophyllum Schott

图6 盐及重金属胁迫处理对绿金合果芋相对电导率的影响Fig.6 Effects of salt and heavy metals stress treatment on relative electrical conductivity of Syngonium podophyllum Schott
3 讨论
本试验研究结果表明,不同于单一重金属及盐胁迫,在复合胁迫下,绿金合果芋对于Pb,Cd,NaCl的吸收与富集机制更加复杂,这一结果与熊春晖等[18]研究结果相一致。原因可能是绿金合果芋对重金属及盐的吸收不仅与外施浓度有关,还与土壤酸碱度、有机质含量、重金属元素的生物有效性以及在土壤中的迁移力等因素有关;同时,相比单一元素胁迫,复合胁迫下元素之间也可能存在一定的相互作用,即一种元素的存在能够影响绿金合果芋对另外一种或几种元素的吸收和富集。
叶绿素和类胡萝卜素是植物光合作用的主要色素,其含量高低直接反映植物光合作用能力的强弱,当植物受到逆境胁迫后可导致光合色素含量下降。大量研究表明植物在盐及重金属胁迫下光合色素含量呈下降趋势,如夏方山等[19]对碱地风毛菊的研究。Alberte[20]认为,逆境胁迫下叶绿素含量下降的主要原因是叶绿体片层中捕光Chlalb-Pro复合体合成受到抑制。植物体内的重金属离子可能与叶绿素合成的几种酶(原叶绿素脂还原酶、δ-氨基乙酰丙酸合成酶和胆色素原脱氨酶)的肽链富含-SH的部分结合, 抑制了酶活性从而阻碍了叶绿素的合成。叶绿素含量降低必然影响植物的光合作用,本试验结果显示:在Pb,Cd,NaCl单一及复合胁迫下,绿金合果芋叶片光合色素含量呈下降趋势,在低浓度的单一胁迫及复合胁迫下,光合色素含量与对照相比变化不大,这说明绿金合果芋对轻度胁迫有一定的耐性,而中度和重度胁迫处理对光合色素的合成具有一定的抑制作用,其中叶绿素a表现得更加敏感;Pb,Cd与NaCl复合胁迫下绿金合果芋叶片光合色素含量呈快速下降趋势,即随着胁迫浓度的增加,叶绿素a、叶绿素b和类胡萝卜素的减少量更大。和单一胁迫一样,其中以叶绿素a的下降幅度最大,这表明该处理对光合色素尤其是叶绿素a的合成具有明显的破坏作用。
可溶性糖、可溶性蛋白、脯氨酸是植物体内重要的渗透调节物质。本研究结果显示:在Pb,Cd,NaCl单一及复合胁迫下,可溶性糖和可溶性蛋白随着胁迫浓度的升高呈下降的趋势,说明已经不能通过可溶性糖和可溶性蛋白的渗透调节作用使合果芋免受伤害。而在低浓度胁迫下,脯氨酸的含量上升,合果芋通过不断积累脯氨酸增强渗透调节能力,但不能有效缓解叶片膜脂过氧化进程,MDA含量呈上升趋势,当胁迫浓度较高时脯氨酸含量下降,叶片细胞膜受到较大伤害,MDA含量进一步升高。
脂质过氧化作用中的自由基反应能使细胞膜系统发生变化,导致细胞的损害甚至死亡。SOD,POD和CAT是植物体内抗氧化系统中的主要保护酶,它们协同作用减少活性氧自由基对细胞膜系统的伤害,本研究表明在Pb,Cd单一和复合胁迫及Pb,Cd与NaCl复合胁迫下,随着胁迫浓度的升高,合果芋叶片的SOD活性呈下降趋势,NaCl单一胁迫时表现为先升高后降低的趋势。POD活性则随着胁迫浓度的升高逐渐降低。CAT活性的变化与二者有所差异。在Pb,Cd单一胁迫和NaCl单一胁迫下,随着胁迫浓度的升高CAT活性先升高后下降;Pb,Cd复合胁迫和Pb,Cd与NaCl复合胁迫下,CAT活性呈下降趋势。这说明在胁迫过程中SOD,POD合成体系遭到破坏,从而减弱了这两种酶对合果芋的保护作用。在处理水平1时,CAT具有较高的活性,从而能够清除合果芋体内有毒害作用的过氧化氢;但从处理水平2开始,随着胁迫浓度的升高该酶活性也随之下降,说明高浓度胁迫下合果芋受到的伤害最为严重。从三种酶的变化趋势可以推测,在重金属和盐的胁迫过程中,起主要保护作用的酶是CAT;并且可以看出,NaCl单一胁迫对合果芋的毒害较其他几种胁迫相对较弱。
MDA含量可以反映细胞膜脂过氧化程度和植物对逆境条件反映的强弱。张凤琴等[21]指出重金属是脂质过氧化的诱变剂,浓度越高脂质过氧化产物 MDA积累越多,两者关系密切。重金属胁迫使得植物组织内MDA含量增加,表明膜脂过氧化程度加剧,透性增大,膜的稳定性降低,细胞内含物被动外渗,影响植物的正常代谢,对植物的毒害作用就越大。本试验结果表明随着重金属和盐胁迫浓度的增大,无论是Pb,Cd,NaCl单一胁迫或复合胁迫,绿金合果芋叶片的MDA含量均呈现上升的趋势,这与本试验中相对电导率的变化一致。说明随着胁迫浓度的增加,绿金合果芋叶片细胞膜的稳定性和结构遭到破坏,膜透性增大,细胞内电解质渗漏加剧,叶片受到的伤害随之加大。
本试验设计都是在盆栽环境下进行的,在自然环境下的重金属及盐胁迫对绿金合果芋生长发育的影响将更加复杂。此外,土壤是一个比较复杂的系统,Pb2+,Cd2+,NaCl进入土壤后,部分离子可能与土壤中的有机物或阴离子形成络合物,胁迫浓度相对减小,同时不同离子间也可能存在一定的协同或拮抗作用。在这些复杂因素影响下,重金属及盐胁迫对绿金合果芋的生理特性有哪些影响,体内的富集、运输和分配情况如何,以及机体如何在胁迫下进行调控,还有待进一步的研究,以便为胁迫污染修复生态工程植物种类的选择提供更科学的依据。
[1]王学礼,马祥庆.重金属污染植物修复技术的研究进展[J].亚热带农业研究,2008, 4(1):44-48.
[2]蒋亚莲,桂敏,黎霞,等. 合果芋组织培养快繁技术研究[J]. 北方园艺,2009 (3):62-65.
[3]叶飞,建德锋. 合果芋组培苗的生根与移栽技术研究[J]. 江苏农业科学,2012, 40(11):196-197.
[4]金琎. 合果芋试管微繁殖技术的研究[J]. 江苏林业科技,2000 (5): 35-37.
[5]任小林,李嘉瑞. 植物生理学通讯[J]. 1991,27(1):34-36.
[6]李合生,孙群,赵世杰. 植物生理生化实验原理和技术[M]. 北京:高等教育出版社,2000.
[7]中国科学院上海植物生理研究所,上海市植物生理学会. 现代植物生理学实验指南[M]. 北京:科学出版社,1999: 303.
[8]张志良. 植物生理学实验指导[M].北京:高等教育出版社,2004.
[9]刘周莉,何兴元,陈祎,等. 镉胁迫下金银花的生长反应及积累特性[J]. 生态学杂志,2009,28(8):1579-1583.
[10]张华新,宋丹,刘正祥. 盐胁迫下11个树种生理特性及其耐盐性研究[J].林业科学研究,2008,21(2):168-175.
[11]原海燕,黄苏珍,郭智,等.铅与盐胁迫对喜盐鸢尾生长及生理抗性的影响[J]. 应用生态学报,2007,18(9):2111-2116.
[12]李学强,李秀珍. 盐碱胁迫对欧李叶片部分生理生化指标的影响[J]. 西北植物学报, 2009,29(11):2288-2293.
[13]何冰,叶海波,杨肖娥. 铅胁迫下不同生态型东南景天叶片抗氧化酶活性及叶绿素含量比较[J]. 农业环境科学学报,2003,22(3):274-278.
[14]江行玉,赵可夫.植物重金属伤害及其抗性机理[J]. 应用与环境生物学报,2007,16(7):92-99.
[15]沈高峰. 重金属Pb对高羊茅生长及抗氧化生理特性的影响[J]. 长江大学学报(自然科学版),2011,8(5):248-251.
[16]张永锋,梁正伟,隋丽,等. 盐碱胁迫对苗期紫花苜蓿生理特性的影响[J]. 草业学报, 2009,18(4):230-235.
[17]秦峰梅,张红香,武炜,等. 盐碱胁迫对黄花苜蓿发芽及幼苗生长的影响[J]. 草业学报,2010,19(4):71-78.
[18]熊春晖,许晓光,卢永恩,等. 铅镉复合胁迫下莲藕对铅镉的富集及其生理变化[J]. 园艺学报,2012,39(12):2385-2394.
[19]夏方山,董秋丽,董宽虎. 碱胁迫对碱地风毛菊生理特性的影响[J].中国农学通报,2010,26(21):152-155.
[20]ALBERTE R S, THOMBER J P, FISCUS E L. Water stress effects on the content and organization of chlorophyll in mesophyll and bundle sheach chlorlplasts of maize[J].PlantPhysiology, 1977, 59(3):351-353.
[21]张凤琴,王友邵,董俊德,等. 重金属污水对木榄幼苗几种保护酶及膜质过氧化作用的影响[J]. 热带海洋学报,2006,25(2):66-70.
(责任编辑张韵)
Effects of heavy metals and salt stress on physiological characteristics of Syngonium podophyllum Schott
ZHANG Jia-yang
(SchoolofLifeScienceandTechnology,XinxiangUniversity,Xinxiang453003,China)
The influence of two heavy metals (Pb2+, Cd2+) , salt stress (NaCl) and their mixed solutions on the photosynthetic system,membrane system and antioxidant system ofSyngoniumpodophyllumSchott were investigated. The results showed that with the increasing concentration of single and mixed solution treatment, chlorophyll, carotenoids, soluble protein and soluble sugar content and POD activity inSyngoniumpodophyllumSchott were all inhibited. The content of free proline and CAT activity increased first and then decreased with the increasing concentration of stress, MDA content and relative conductivity increased with increasing concentration of stress. SOD activtly decreased with the increasing concentration of heavy metals, but increased first and then decreased with the increasing concentration of NaCl. This study suggested that, chlorophyll content, membrane system and antioxidant system ofSyngoniumpodophyllumSchott were sensitive to the stress of heavy metals and salt. These parameters could be taken as indicators in evaluating the damage of heavy metals and salt stress onSyngoniumpodophyllumSchott.
heavy metal; salt stress;SyngoniumpodophyllumSchott; physiological characteristics
10.3969/j.issn.1004-1524.2016.04.10
2015-10-09
新乡市重点科技攻关项目(ZG14035)
张家洋(1975—),男,河南信阳人,硕士,讲师,从事污染生态学等研究。E-mail: skxsyszr@163.com
S688
A
1004-1524(2016)04-0601-08
张家洋. 重金属及盐胁迫对绿金合果芋生理特性的影响[J].浙江农业学报,2016,28(4): 601-608.
猜你喜欢
阅读(科学探秘)(2020年8期)2020-11-06
矿产综合利用(2020年1期)2020-07-24
中国果业信息(2019年1期)2019-01-05
中成药(2018年8期)2018-08-29
中成药(2018年8期)2018-08-29
生物学教学(2017年9期)2017-08-20
当代化工研究(2016年6期)2016-03-20
中国资源综合利用(2016年3期)2016-01-22
医学研究杂志(2015年9期)2015-07-01
中国当代医药(2015年9期)2015-03-01
