
重组人睫状神经营养因子干预家兔视神经损伤动物模型视神经蠕变特性的分析
2016-10-18 10:14:54王玲吕雪漫姜琦袁毅
生物医学工程研究 2016年2期
王玲,吕雪漫,姜琦,袁毅
(吉林大学中日联谊医院眼科,吉林 长春 130033)
1 引 言
视神经由视网膜神经节细胞的轴突和胶质等组成, 其损伤可以发生在视神经的每个部位, 引起视功能部分或全部丧失, 视神经损伤的预防和治疗是视觉科学领域研究的热点及难点[1]。 国内外学者对视神经损伤诊断和治疗进行了大量的研究[2-5]。贺致礼[6]对自发性分化的特点、外源性和内源性睫状神经营养因子对于细胞分化结果的影响进行了研究。结果表明, 睫状神经营养因子能够起到部分介导神经祖细胞自发性分化的作用。并且自发性分化是在培养基中去除有丝分裂原之后发生的。 Birch等[7]以CNTF-4对 3例患者纵向性研究表明,根据自适应光学激光扫描眼底镜提供的高分辨率图像,与治疗侧眼睛相比较,假手术侧眼睛的锥间距增大,锥密度降低,提示,CNTF 具有视锥细胞保护作用。
关于视神经损伤模型以药物干预后视神经生物力学特性研究,Jiang等[8]对视神经损伤家兔模型通过以重组人睫状神经营养因子和中药复光颗粒治疗视神经损伤家兔后,眶内段视神经单向拉伸力学性质进行了研究。得出了视神经损伤家兔模型通过以重组人睫状神经营养因子和中药复光颗粒干预治疗后,眶内段视神经拉伸力学性能指标有一定恢复的结论。以蠕变特性指标探讨重组人睫状神经营养因子干预视神经损伤效果罕见报道。鉴于此,实验以视神经损伤兔为动物模型,以重组人睫状神经营养因子进行干预治疗,以蠕变特性研判重组人睫状神经营养因子干预治疗视神经损伤的效果,为临床视神经损伤治疗提供蠕变特性指标。
2 材料与方法
2.1 实验动物
6月龄健康日本大耳白家兔33只, 体重2.7~2.9 kg, 均为雌性,购于长春高新医学动物实验中心,许可证号SCXK(吉)2003-0004。实验前以 10%(35 mg/Kg)水合氯醛注射家兔腹腔麻醉,检查家兔外眼及眼底无病变的纳入实验。家兔在兔笼中饲养,家兔饲养室自然光照,室内温度23℃-25℃, 家兔饲养室内空气流通,家兔饲养室相对湿度55%~70%,家兔食用饲料为兔颗全价粒料,家兔自由饮水、摄食。
2.2 实验动物分组
33只日本大耳白家兔随机分为正常对照组11只、视神经损伤动物以重组人睫状神经营养因子干预组11只、视神经损伤动物模型组11只。
2.3 家兔视神经损伤模型制备
按文献[9]的方法复制家兔视神经损伤模型,将家兔俯卧固定于动物手术台上。以浓度为1 g /L的乌拉坦( 5 mg /kg)兔耳缘静脉注射麻醉,以手术刀切开兔右侧眼顶侧眶上皮肤, 暴露眶壁、眶上缘切迹, 在眶上缘切迹处向视神经管方向咬除两侧部分眶壁骨板(宽6 mm,深7~8 mm ), 剪开眼球后球筋膜, 沿上直肌向球后分离, 使球后视神经暴露。视神经游离约5 mm, 离后极部约3 mm,以上海医疗器械厂W40160(3. 7 cm )反向血管夹钳夹同一部位视神经5 s。术野用庆大霉素液冲洗两次后,10~0号尼龙线分层缝合。术后在家兔眼结膜囊内涂红霉素眼膏。
2.4 造模成功标准
家兔损伤伤眼直接对光反应消失,瞳孔固定、散大;以检眼镜观察眼底,视网膜血管无梗塞与出血为造模成功标准,达到以上标准的家兔作为视神经损伤模型纳入实验。
2.5 以重组人睫状神经营养因子干预视神经损伤动物方法
视神经损伤动物以重组人睫状神经营养因子干预组家兔于手术(造模)后立即向球后一次性注射 CNTF 50 ng/ml[9],视神经损伤动物模型组不给予治疗视神经损伤的药物。
2.6 各组家兔视神经标本取样方法
造模30 d后, 处死重组人睫状神经营养因子干预组、视神经损伤模型组、正常对照组家兔,在镜下取出眶内段视神经标本各11个,置于生理盐水槽中,在4℃环境下保存备用。
2.7 各组动物视觉电生理检测
对视神经损伤动物模型以重组人睫状神经营养因子干预组、视神经损伤动物模型组动物于造模30 d后分别对动物损伤眼(右眼)和正常眼(左眼)以德国罗兰REFZ-POZE21sompace型视觉电生理仪进行闪光视觉诱发电位(F-VEP) 检测。记录电极放于家兔枕骨结节上1.5~2.0 cm处,参考电极置于家兔额部正中,接地电极置于家兔耳垂;将角膜接触镜电极放好,接地皮肤电极置于耳垂,参考皮肤电极置于眼眶颞侧,。电极放好后,以全视域闪光刺激器, 白色闪光, 闪光照度2 挡, 60 lx, 时间频率2 Hz进行刺激。分析时间250 ms, 数据经由算机进行采集。
1.若等腰三角形的一边长为6cm,另一边长为9cm,求此等腰三角形的周长。(6cm是腰长还是底边长?)
2.8 光学显微镜观察兔视神经纵断面的组织结构
取各组家兔眶内视神经各一条,标本固定于10%中性福尔马林液 24 h。常规梯度乙醇脱水,二甲苯透明,石蜡包埋;纵向切片,厚度为 3 μm,蜡带裱于涂有蛋白甘油的载玻片上;温箱烤片,切片常规脱腊;苏木精染色3 min,自来水冲洗;1%盐酸分化,自来水冲洗;弱氨水返蓝,自来水冲洗;伊红染色2 min;常规梯度乙醇脱水,二甲苯透明,中性树胶封片;以日本东京Olympus BX51光学显微镜分别观察每个视神经的形态。
2.9 蠕变实验方法
实验设备为长春试验机研究所集团生产的电子万能材料试验机,以长春市第三光学仪器厂生产的读数显微镜测量各组家兔视神经试样的长度和直径,各组家兔视神经拉伸试样长度为10 mm,正常对照组家兔视神经试样直径1.01~1.03 mm,视神经损伤模型组家兔视神经试样直径0.97~0.99 mm,视神经损伤以重组人睫状神经营养因子干预组家兔视神经试样直径1.00~1.02 mm。按文献[10-12]的方法分别对每个家兔视神经试样进行预调处理。视神经拉伸实验温度为36.5±1.0℃,分别将3组家兔视神经试样装夹在试验机夹头内,以0.2 Mpa/s的速度对试样施加应力,当正常对照组视神经试样应变达到3.98%、视神经损伤模型以重组人睫状神经营养因子干预组试样应变达到3.85%、视神经损伤模型组试样应变达到3.78%时,各组视神经试样应力达到0.454 Mpa时,保持应力恒定,设定视神经试样蠕变实验时间为7 200 s。观察视神经试样应力与时间的变化关系。为在实验中保持试样的湿度,在实验中向视神经试样喷洒生理盐水。
2.10 统计学分析
计量资料以mean±SD表示,用SPSS16.0 软件包(SPSS, Chicago, IL, USA)进行数据分析,组间数据差异的比较采用单因素方差分析法和Sceffe法,P<0.05 为差异有显著性意义。
3 结果
3.1 各组视神经电生理检测结果
F- VEP 记录 图形分析标准: P1 波幅值为自N1 波谷底至P1 波峰顶, 潜伏期为自触发时至P1峰为止, 由计算机自动记录, 各组动物电生理检测结果见表1。
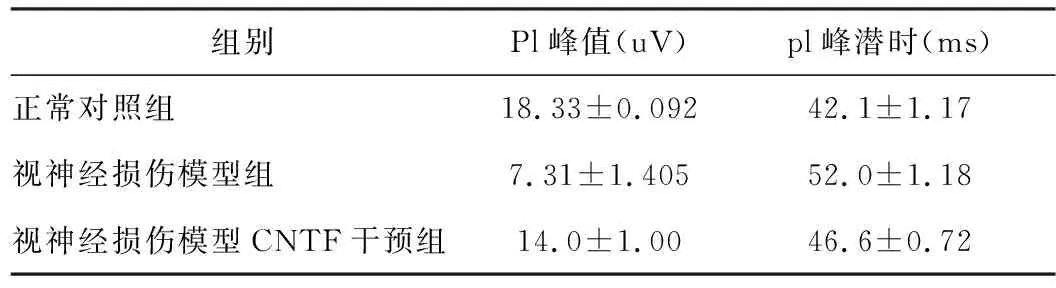
表1 各组视神经电生理检测实验结果
各组视神经电生理检测实验结果表明,正常对照组Pl峰值大于视神经损伤模型组和视神经损伤动物模型以人睫状神经营养因子干预组,差异显著(P<0.05),正常对照组pl峰潜时值小于视神经损伤模型组和视神经损伤模型以重组人睫状神经营养因子干预组,差异显著(P<0.05)。视神经损伤模型以人睫状神经营养因子组Pl峰值大于视神经损伤模型组,差异显著(P<0.05),视神经损伤模型以重组人睫状神经营养因子干预组组pl峰潜时值小于视神经损伤模型组,差异显著(P<0.05)。
3.2 各组视神经的超微结构
正常对照组视神经纵断面的组织形态(HE染色,×400)纤维呈密集的平行排列,结构排列整齐规则,染色均匀,胶质细胞大小、排列均匀见图1(a),视神经损伤模型组兔视神经纵断面的病理学改变(HE染色,×400)神经纤维束结构消失,神经纤维和胶质细胞变性、坏死,偶见溶解的细胞核见图1(b),视神经损伤模型以重组人睫状神经营养因子干预组的兔损伤视神经纵断面的病理学改变(HE染色,×400)视神经纤维结构尚存,较少部分视神经纤维排列不规则,视神经无明显变细,见图1(c)。

图1 各组视神经的超微结构照片
3.3 各组视神经试样蠕变曲线
由各组试样的蠕变试验数据以计算机进行曲线拟合,蠕变曲线见图2。
3.4 各组视神经试样的归一化蠕变函数曲线
对各组视神经试样归一化蠕变函数数据以计算机进行曲线拟合,各组视神经试样归一化蠕变函数曲线见图3。

图2 各组视神经试样蠕变曲线
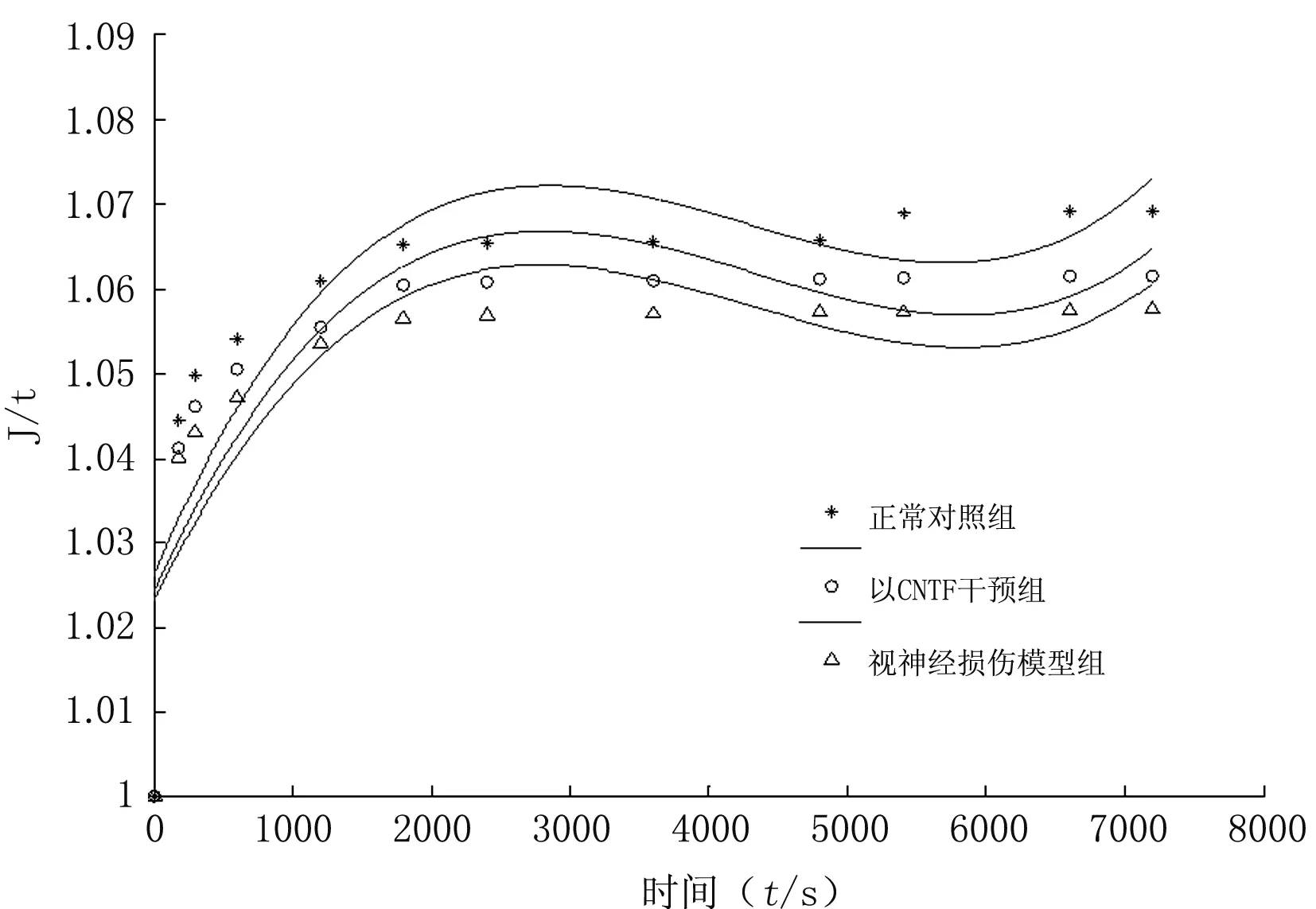
图3 各组视神经试样归一化蠕变函数曲线
3.5 各组视神经试样归一化蠕变函数方程的建立
J(t)=a+be-t
(3)
令:
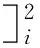
正则方程:
(4)
分别将各组视神经蠕变实验数据代入(4)式,解出各组试样的a、b值,将各组试样的a、b值分别代入(3)式得出两组试样的归一化蠕变函数方程如下:
正常对照组:
视神经损伤模型组:
视神经损伤模型以重组人睫状神经营养因子干预组:
4 讨论
各组家兔视神经电生理检测结果表明,视神经损伤模型以重组人睫状神经营养因子干预组Pl峰值大于视神经损伤模型组,差异显著(P<0.05),视神经损伤模型以重组人睫状神经营养因子干预组pl峰潜时值小于视神经损伤模型组,差异显著(P<0.05),说明重组人睫状神经营养因子对视神经损伤动物模型的视神经具有一定的疗效。
各组视神经蠕变实验结果表明,视神经损伤模型以重组人睫状神经营养因子干预组7 200 s应变上升量大于视神经损伤模型组(P<0.05)。各组家兔视神经蠕变实验最初1 800 s应变上升速度快,之后应变缓慢上升,达到设定的蠕变实验时间7 200 s时,各组家兔视神经试样蠕变曲线基本接近于水平,各组视神经试样蠕变曲线变化趋势呈指数函数关系。蠕变是视神经对生理应力适应性的反应,视神经生理范围内的蠕变特性有利于抵抗应力,视神经的生理功能要求视神经纤维的结构完整,结构完整的视神经才具有良好的蠕变特性。分析认为视神经损伤模型家兔视神经受到钳夹伤后,使视神经结构丧失了完整性,将会使视神经全部和部分丧失抵抗应力的能力,所以,视神经损伤模型组的蠕变特性发生了改变。
本实验结果显示,CNTF组视神经纤维排列较紊乱,胶质细胞核增多,部分排列不规则,可见少量细胞空泡,胞核溶视神经纤维迂曲、偶见胞核碎片,但视神经无明显变细,较模型组损伤程度轻,通过以CNTF对视神经损伤动物进行治疗,对破坏的视神经组织形态有所改变,在长时间恒应力作用下的应变上升量有所提高,说明视神经损伤以重组人睫状神经营养因子干预后蠕变特性具有一定的恢复,说明CNTF对治疗动物视神经损伤具有一定的作用。
视神经的损伤发生机制与视神经的生物力学性质密切相关。我们研究了以重组人睫状神经营养因子干预家兔视神经损伤动物模型视神经的蠕变特性。以回归分析的方法根据已得的蠕变实验结果以及以往的经验建立统计模型构建了各组坐骨神的蠕变函数方程;研究了各组视神经应变与时间之间的相关关系,建立起应变与时间变量之间的函数关系。各组视神经蠕变函数方程的建立能更好的阐明视神经的蠕变特性。从而给临床中相同发病机制而造成视神经损伤提供粘弹性力学方面的理论依据。
以往对动物视神经损伤模型药物干预效果分析,以生物学、组织形态学观察等居多。本研究是以蠕变指标研判重组人睫状神经营养因子干预视神经损伤的效果,具有创新性。
猜你喜欢
山东冶金(2022年3期)2022-07-19 03:25:22
今日农业(2021年17期)2021-11-26 23:38:44
基层中医药(2021年8期)2021-11-02 06:24:54
科学(2020年3期)2020-11-26 08:18:30
Annals of Applied Mathematics(2020年3期)2020-09-14 10:51:02
中州大学学报(2019年3期)2019-07-17 06:45:12
中医眼耳鼻喉杂志(2019年3期)2019-04-13 05:26:50
武汉工程职业技术学院学报(2017年1期)2017-04-24 08:34:24
兽医导刊(2015年9期)2016-01-04 12:00:06
兽医导刊(2015年7期)2016-01-04 11:59:58
