
小麦籽粒WSC含量QTL的整合与元分析
2016-09-21 09:46胡雅君栗孟飞杨德龙程洪波柴守玺
麦类作物学报 2016年8期
胡雅君,栗孟飞,杨德龙,刘 媛,程洪波,常 磊,柴守玺
(1.甘肃省干旱生境作物学重点实验室/甘肃农业大学生命科学技术学院,甘肃兰州 730070;2.甘肃农业大学农学院,甘肃兰州 730070)
小麦籽粒WSC含量QTL的整合与元分析
胡雅君1,栗孟飞1,杨德龙1,刘 媛1,程洪波1,常 磊2,柴守玺2
(1.甘肃省干旱生境作物学重点实验室/甘肃农业大学生命科学技术学院,甘肃兰州 730070;2.甘肃农业大学农学院,甘肃兰州 730070)
在干旱胁迫条件下,小麦营养器官暂贮性可溶性碳水化合物(Water-soluble carbohydrates,WSC)是小麦籽粒灌浆所需的重要碳源。为发掘控制小麦籽粒WSC含量的真实主效数量性状位点(Quantitative trait loci,QTL),利用生物信息学方法,借助小麦高密度分子标记遗传图谱,对来自不同遗传作图群体的控制小麦籽粒WSC含量的168个QTL位点进行图谱映射和元分析。结果发现,142个QTL定位区间与参考图谱有共有标记,其中92个QTL对籽粒WSC含量的表型变异具有增效效应,50个QTL具有减效效应。建立控制小麦籽粒WSC含量的QTL一致性图谱,获得16个“一致性”QTL (Meta quantitative trait loci,MQTL)位点及其连锁标记,MQTL置信区间最小达到0.77 cM。
小麦;可溶性碳水化合物;一致性数量性状位点;元分析
干旱是制约小麦稳产增产的重要环境限制因子[1]。随着干旱胁迫的季节性频发,每年因干旱缺水造成的小麦减产在30%以上,甚至绝产[2]。研究表明,小麦能否正常灌浆对最终产量形成至关重要,小麦灌浆期籽粒物质的来源,除源于小麦植株绿色部分光合产物外,还与小麦各营养器官花前暂贮存性可溶性碳水化合物(Water soluble carbohydrates,WSC)密切相关,且后者对籽粒产量的贡献更大[3]。因此,小麦生育后期营养器官WSC向籽粒的高效转运和再分配对小麦产量形成至关重要[4]。尤其是在干旱胁迫条件下,当小麦灌浆期旗叶光合产物无法满足冠层呼吸消耗和保持籽粒正常灌浆需要时,最终产量形成主要依赖于花前贮存在茎秆中的WSC[4]。
研究表明,小麦籽粒WSC是典型的微效多基因控制的复杂数量性状,其遗传机理复杂,易与环境发生显著互作[4]。目前,研究者利用不同遗传背景材料和遗传图谱,对小麦籽粒WSC含量进行了数量性状位点(Quantitative trait loci,QTL)定位和遗传分析[4-11]。如Huynh等[5]利用小麦双单倍体群体(Double haploid lines,DH)定位了5个控制籽粒果聚糖性状QTL,分别位于2B、3B、6D和7A染色体上,可解释4%~27%的表型变异。付夕阳等[6]利用小麦重组自交系(Recombinant inbred line,RIL)群体,定位了13个控制籽粒WSC含量的QTL,位于1B、1D、2B、2D、3B、4A、4B、5A、5D、6B和7B等11条染色体上,可解释4.97%~53.57%的表型变异。但是,由于前人采用了不同的组配亲本、作图群体、实验设计、统计方法、标记类型和群体大小等,导致即使是同一性状,所定位出的QTL位点数目、染色体位置和基因效应差异较大,极难获得真实、稳定控制小麦WSC含量的QTL区段或位点,无法对这些QTL区段或位点在育种实践中应用[4]。
Arcade等[12]开发出用于QTL元分析的BioMercator软件,能将与某一性状相关的所有QTL定位数据映射到一张遗传图谱上,对多个独立试验的QTL数据比对分析,预测一致性真实QTL位点,并进一步缩小其置信区间,提高定位精确度[7]。在玉米上,Chardon等[13]利用元分析对玉米进行研究,通过收集313个与玉米花期相关的QTL,发现62个一致性QTL(Meta quantitative trait loci,MQTL),发现利用元分析方法,其定位准确度是相应区域定位最准确的QTL的两倍;方永丰等[14]对来自7个作图群体的173个控制玉米持绿相关相关性状的QTL进行元分析,共获得5个控制玉米持绿性的MQTL。这些MQTL主要分布于1、4、5和9号染色体上。在大豆上,吴 琼等[15]利用RIL群体发掘出大豆2个重要生育时期共 9个MQTL,主要分布在大豆C2、M、L、F、J、D1a和D1b连锁群上。但迄今为止,有关小麦相关性状的QTL整合和元分析研究甚少,尤其是关于小麦籽粒WSC含量的QTL元分析研究未见报道。鉴于此,本研究利用生物信息学方法,将已报道的控制小麦籽粒WSC含量的QTL数目和位点进行收集整理,以小麦高密度遗传图谱作为参考图谱,利用BioMercator 4.2软件(http://moulon.inra.fr/index.Php/fr/component/docman/cat_view/21-logiciels/101-abi-project-and-software/104-biomercator-v21)将QTL映射到该参考图谱上,构建小麦籽粒WSC的QTL整合图谱,通过元分析的方法进行MQTL区间的鉴定,以期为进一步研究小麦籽粒WSC含量的QTL精细定位以及图位克隆提供理论依据。
1 材料与方法
1.1材 料
以美国农业部小麦公共数据库(http://wheat.pw.usda.gov/)和已发表的文献中控制小麦籽粒WSC含量的QTL信息[4-11]为研究对象。
1.2试验方法
1.2.1小麦籽粒WSC含量QTL数据的收集
对已报道的小麦籽粒WSC含量QTL定位信息进行收集,将收集到的每一个QTL数据按BioMercator 4.2软件的要求整理,包括QTL名称、染色体位置、置信区间、连锁系数、贡献率和群体大小等,其中,QTL位置(置信区间和QTL的最大可能位置)和贡献率作为影响QTL元分析的两个重要因素[12]。
1.2.2小麦籽粒WSC含量QTL信息的整合
根据收集到的小麦籽粒WSC含量QTL信息确定其所涉及的染色体,以Wheat composite 2004 (http//wheat.pw.usda.gov/ggpages/mapse shortlist.html.)高密度遗传图谱作为参考图谱,将控制小麦籽粒WSC含量QTL的最大可能性位置及置信区间两端坐标按比例标注到参考图谱上,对比原始图谱(即所收集的遗传图谱)与参考图谱。如果没有公共标记,不能直接映射到参考图谱上,为了获得精确的映射结果,先映射到Somers等[16]绘制的小麦整合图谱(即中介图谱)上,再利用图谱上的公共标记将QTL映射到参考图谱上。并将原始图谱与参考图谱上相关标记载入BioMercator 4.2软件中,建立图谱信息库。基于图谱间的共有分子标记,利用齐序函数[13]将控制小麦籽粒WSC含量QTL的最大可能性位置和置信区间两端坐标按比例标注到参考图谱上,即映射[13],并将在原始图谱和参考图谱间有争议的标记剔除。
1.2.3小麦籽粒WSC含量QTL的元分析
利用BioMercator 4.2软件进行小麦籽粒WSC含量QTL的元分析。将位于同一连锁群相同位点附近的N个独立存在的与目标性状相关的QTL进行运算,对独立来源的同一性状且位于同一座位或有重叠座位的QTL计算出一个MQTL,该QTL会给出5个模型(模型1、2、3、4和N),其中可靠性检验值(Akaike-type criteria values,AIC)最小的模型为最优模型,即真实QTL模型。如果某一QTL置信区间未知,可通过Darvasi和Soller[19]的公式计算95%的置信区间。
(1) C.I=530/(N×R2)
(2) C.I=163/(N×R2)
公式中C.I为QTL 95%的置信区间,N为研究群体的大小,R为目标性状QTL的贡献率。公式(1)适用于F2群体和BC群体,公式(2)适用于RIL、DH以及近等基因系群体(Near isogenic lines,NIL)。若已知置信区间,也可以应用该公式估算未知QTL贡献率。
2 结果与分析
2.1小麦籽粒WSC含量QTL信息的收集和图谱整合结果
从小麦公共数据库和已发表的文献中收集到来源于Hanxuan 10×Lumai 14、Berkut×Krichauff、R97× R146、PH82/2×Neixiang 188、Chuan 35050×Shannong 483、Cranbrook×Halberd、Suco×Tasman 和CD87×Katepwa等8个作图群体,共涉及小麦籽粒WSC含量的168个QTL(表1)。利用BioMercator 4.2软件中的QTL projection功能将收集到的QTL通过中介图谱映射到参考图谱上,成功映射到142个QTL定位区间与参考图谱有共有标记(表2),包括92个对小麦籽粒WSC含量的表型变异具有增效效应的QTL和50个具有减效效应的QTL。这些QTL在各自的连锁图谱定位中分布比较分散,LOD值在2.0~30.0之间,各位点遗传贡献率在2.6%~26.5%之间。
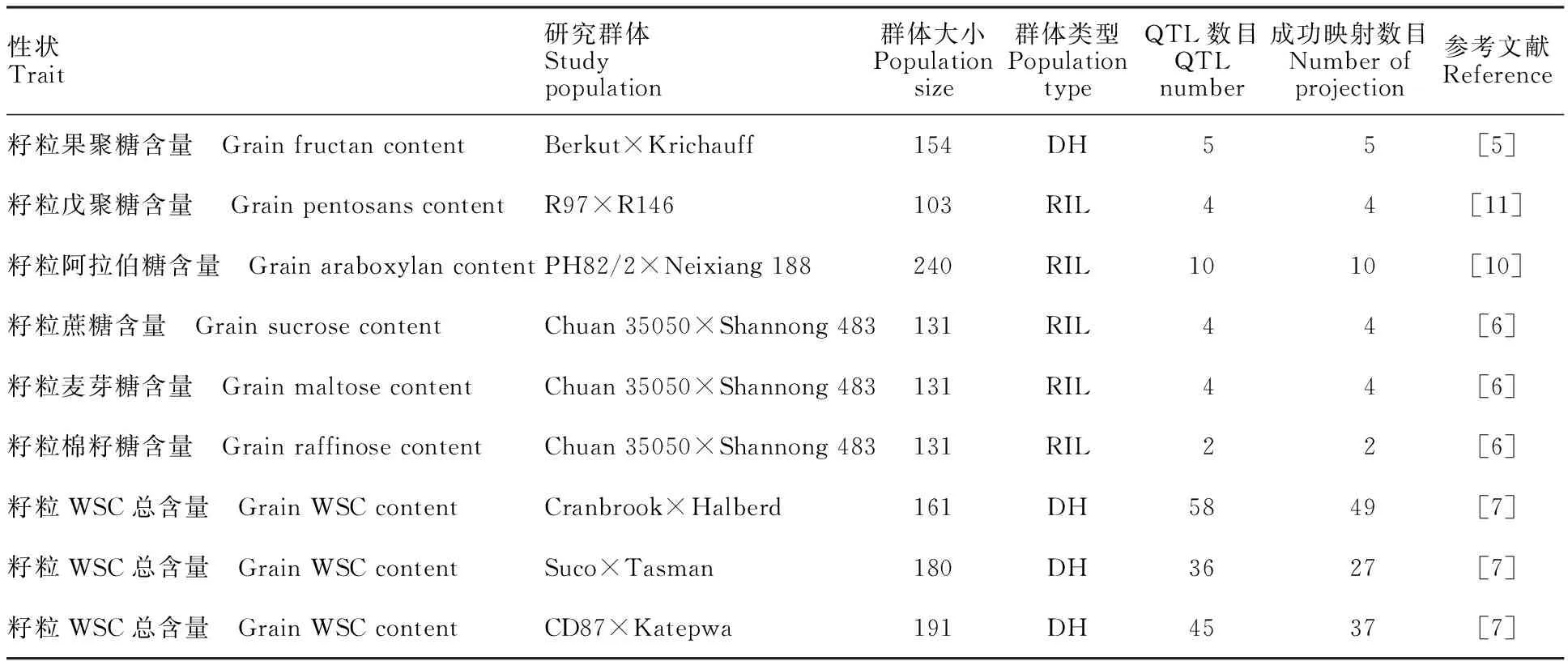
表1 小麦籽粒WSC含量QTL数据的整合
利用图谱整合的方法,将所收集的QTL整合到参考图谱上。如图1所示,被整合的QTL分布在小麦染色体1A、1B、3B、4B、4D、5B、6D、7A和7D上,但各染色体上分布不均匀,在6D染色体上最少(1个QTL),7A染色体上最多(7个QTL),其他染色体上分布2~5个。
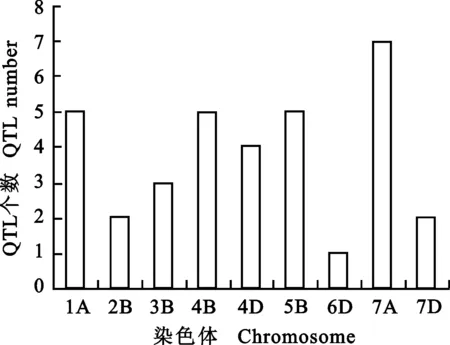
图1 小麦籽粒WSC含量的QTL在不同染色体上的分布
其中,某些QTL在同一连锁群上成簇存在,共有10个QTL簇,在4B、4D和5B连锁群上各发现2个QTL簇,在1A、3B、7A和7D连锁群上各发现1个QTL簇。在4B、4D和5B连锁群上的1个QTL簇均包含2个QTL,在1A连锁群上的1个QTL簇包含4个QTL,在3B连锁群上的1个QTL簇包含3个QTL,在7A连锁群上的1个QTL簇包含6个QTL,在7D连锁群上的1个QTL簇包含2个QTL。这些QTL彼此间有区间的重叠,大都出现在一段区间内,如7A连锁群的20 cM左右有6个QTL。同一区间内不同研究都发现了相同QTL,说明这个QTL区间存在更为真实重要的QTL,可能含有大量控制小麦籽粒WSC含量的基因(图2)。
2.2小麦籽粒WSC含量QTL的元分析
结合小麦已定位的QTL信息,利用Biomercator 4.2软件中元分析程序分析各连锁群上的QTL。由于分析模型不同,以每次元分析中AIC值最小的区间为最优,确定1个真实QTL,最终得到16个控制小麦籽粒WSC含量的MQTL(表2),分布于小麦1A(2个)、1B(2个)、3B(2个)、4B(2个)、4D(2个)、5B(1个)、6D(1个)、7A(2个)和7D(2个)染色体上,平均每条染色体上含有1.8个MQTL。在1A染色体上确定了2个控制籽粒WSC总含量的MQTL,分别位于59.13 cM和96.48 cM处,置信区间为57.59~60.66 cM和96.48~106.58 cM,分别在标记P39~Xgbx3076和Xcfa2056b~Xpsr967.1之间,图距为3.07 cM和10.10 cM。在1B染色体上确定了2个MQTL,均是控制籽粒WSC总含量的MQTL,分别位于48.19 cM和86.64 cM处,置信区间为45.19~51.39 cM和84.64~91.64 cM,在标记Xabc155c~Xwmc320和Xmwg69.1~Xcdo1189之间,图距为6.20 cM和7.00 cM。在3B染色体上确定了2个MQTL,均为控制籽粒WSC总含量的MQTL,分别位于44.42 cM和62.77 cM处,置信区间为39.94~48.9 cM和49.77~63.67 cM,在标记Xfbb24~XksuH7和XksuH7~Xbarc7之间,图距为8.96 cM和13.90 cM。在4B染色体上确定了2个MQTL,一个控制籽粒蔗糖含量,另一个控制籽粒阿拉伯木聚糖含量,分别位于11.45 cM和29.25 cM,置信区间为10.40~12.49 cM和21.44~37.07 cM,在标记Xmwg2025a~Xgwm495a和标记XgbxG057~Xcdo1401之间,图距分别为2.09 cM和15.63 cM。在4D染色体上确定了2个MQTL,一个控制籽粒WSC总含量,另一个控制籽粒阿拉伯木聚糖含量,分别位于18.25 cM和46.83 cM,置信区间为12.99~23.52 cM和38.93~54.73 cM,在标记Xbcd1930b~Xwmc89b和标记XgbxG102~Xfba211之间,图距为10.53 cM和15.80 cM。在5B染色体上确定了1个控制籽粒戊聚糖含量的MQTL,位于31.40 cM处,置信区间为17.17~45.64 cM,在标记Xfba232a~Xpsr326b之间,图距分别为28.47 cM和3.89 cM。在6D染色体上确定了1个控制籽粒果聚糖含量的MQTL,位于65.51 cM处,置信区间为59.51~73.51 cM,在标记Xfba187~Xcfd80之间,图距为14.00 cM。在7A连锁群上确定了2个MQTL,一个控制籽粒果聚糖含量,位于0.88 cM,置信区间为0.29~1.06 cM,图距为0.77 cM,在标记Xgwm344~Xfba93之间;另一个控制籽粒棉籽糖含量,位于72.47 cM处,置信区间为71.47~77.67 cM,图距为6.20 cM,在标记Xwmc83a~XksuH9之间。在7D连锁群上确定了2个MQTL,一个控制籽粒麦芽糖含量,另一个控制籽粒WSC总含量,分别位于30.03 cM和52.29 cM处,置信区间为29.03~33.63 cM和48.29~61.19 cM,在标记Xcdo1400~Xcfd41和Xgpw1142a~Yr18之间,图距分别为4.60 cM和12.90 cM。
16个控制小麦籽粒WSC含量的MQTL按所在染色体的位置依次排序为MQTL1~MQTL16,其中5个MQTL的置信区间小于5 cM,分别是MQTL1(3.07 cM)、MQTL7(2.09 cM)、MQTL12(3.89 cM)、MQTL14(0.77 cM)、MQTL16(4.60 cM)。经过元分析后,这些检测到的MQTL都缩小了原QTL的置信区间,使MQTL的区间更精细,置信区间也更窄,很大程度上减小了由于不同试验所得到的QTL位置差异而造成的误差,提高了MQTL定位的准确度和有效性。

染色体左侧“点”至“横线”表示QTL所在位点的遗传贡献率大小的连续变化;“竖线”表示QTL的所在置信区间。
The “dot” to “transverse line” on the left side of chromosome mean the successive changes in genetic contribution rates of QTL; “Vertical line” means the confidence interval of QTL.
图2小麦籽粒WSC含量的QTL的一致性图谱
Fig.2Consensus map of QTL for grain WSC content in wheat
3 讨 论
近几年,利用元分析方法对玉米等性状QTL的遗传改良已经取得了一定进展。栗文娟等[17]整合了360个玉米抗旱相关的QTL,得到79个MQTL及其连锁标记,并通过生物信息学分析,获得43个区间内包含抗旱相关的基因信息。将4个玉米抗旱相关的基因家族定位到玉米18个MQTL区段内。王帮太等[18]整合 400 个玉米产量及产量相关性状 QTL 的基础上,采用元分析方法发掘了96个“一致性”QTL,缩小了原QTL的置信区间,发现关联性较强的产量性状的 QTL常集中在相同或相近的座位上。在玉米上,基因组测序基本完成,玉米数量性状基因(Quantitative trait gene,QTG)的克隆进展很快,当大量的控制玉米目标性状的基因或 QTL 被整合到一致性图谱上后,控制相同和不同的目标性状的 QTL 分布特征将更加明显。然而利用元分析技术对小麦数量性状进行QTL整合研究的报道较少,尤其是关于小麦籽粒WSC含量的元分析还处在起始阶段。但随着小麦基因组信息的日渐完善以及物理图谱的成功构建,对于成簇分布的 QTL 富集区域,以及被前人反复证实的小麦目标性状 QTL 的核心染色体区段,将是发掘小麦目标性状关键基因的重点研究部位,最终可以为小麦目标性状 QTG 的克隆提供大量候选基因,进而提高小麦目标性状基因的克隆效率。
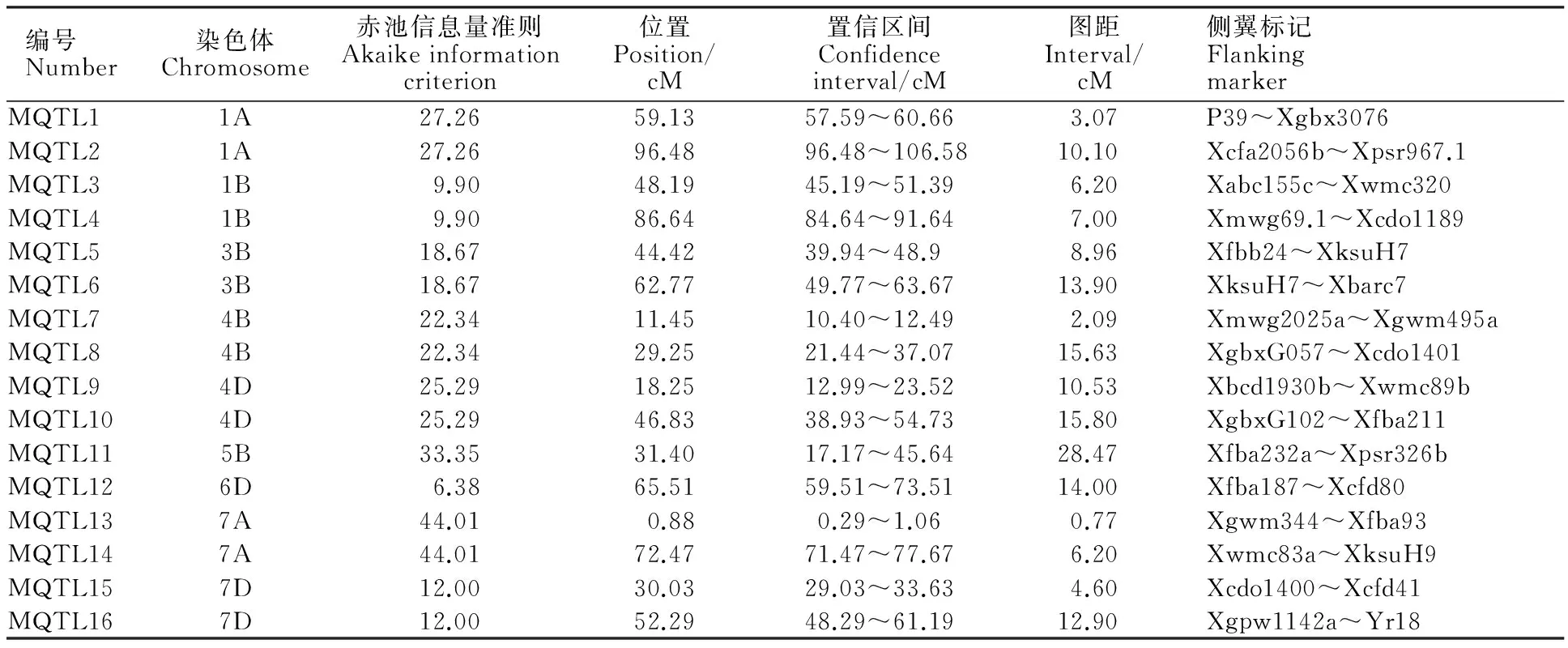
表2 小麦籽粒WSC含量QTL的元分析
在小麦上,王健维等[19]通过元分析技术,检测到264个与小麦品质相关的MQTL,最小置信区间仅有0.04 cM,被整合的QTL在小麦各条染色体的不同区段分布不均,并有集簇现象,其中,1A染色体中Xgdm136~Xgbx3076区段与本研究1A染色体中集簇区段Glu-A3~Xgbx3076重合,4D染色体中Xwmc89~Xwmc473区段与本研究4D染色体中集簇区段Xwmc825~Xwmc473重合,Tuberosa等[20]对这种簇集分布的特征做了多种预测,其中一因多效,或者控制不同性状的基因紧密连锁可能是其主要原因。说明在这些区段上可能存在“一因多效”的基因,同时控制小麦品质性状和小麦籽粒WSC含量。
本研究整合了近年发表的168个小麦籽粒WSC含量的QTL,构建了小麦QTL一致性图谱,具簇集分布特征,其中共有10个QTL簇。并通过元分析,得到16个MQTL及其连锁标记,其置信区间最小可缩小到 0.77 cM,大大减少了 QTL的误差,使结果更趋准确可靠。建立小麦籽粒WSC含量QTL的一致性图谱是对不同遗传背景、不同环境条件和使用不同类型标记条件下定位结果的总结。该图谱可应用于对小麦籽粒WSC含量的进一步精细定位和图位克隆,为深入理解小麦籽粒WSC含量的遗传控制机制、发掘可供超高产小麦新品种选育中利用的功能分子标记以及创建小麦目标性状MQTL的近等基因系群体奠定了基础。本研究所发掘的MQTL是基于不同作图群体以及不同自交系整合的,对后续的研究奠定了基础,可利用分子标记辅助选择技术(Marker assisted selection,MAS),对不同群体的后代进行基因型预测,再结合表型数据,初步完成分子辅助选择,提高 MAS 的实际效率[21]。此外,一些QTL密集的热点区域可以为物理图谱的构建提供切入点[22],用来寻找和定位同源基因[23],还可以针对热点区域中的基因或QTL开展分子设计育种,为候选基因挖掘和QTL到QTG 的进程提供有利依据。
[1]赵紫平,邓西平,刘立生,等.灌浆期干旱对不同倍性小麦光合和产量的影响[J].麦类作物学报,2009,29(3):470-475.
ZHAN Z P,DENG X P,LIU L S,etal.Effect of water deficit on photosynthetic capacity and yield of different ploidy wheat during grain filling stage [J].JournalofTriticeaeCrops,2009,29(3):470-475.
[2]MUGO S N,BANZIGER G O.Edmeades prospects of using ABA in selection for drought tolerance in cereal crops [J].PlantPhysiology,1992,4:112-116.
[3]胡继超,曹卫星,姜 东,等.小麦水分胁迫影响因子的定量研究Ⅰ.干旱和渍水胁迫对光合、蒸腾及干物质积累与分配的影响[J].作物学报,2004,30(4):315-320.
HU J C,CAO W X,JIANG D,etal.Water stress impact factor of quantitative research of wheatⅠ.Dry and wet stress on photosynthesis,transpiration and the influence of dry matter accumulation and distribution [J].ActaAgronomicaSinica,2004,30(4):315-320.
[4]YANG D L,JING R,CHANG X P,etal.Identification of quantitative trait loci and environmental interactions for accumulation and remobilization of water-soluble carbohydrates in wheat (TriticumaestivumL.) stems [J].Genetics,2007,176:571-584.
[5]HUYNH B,JAMES C R,KERRIE L.Quantitative trait loci for grain fructan concentration in wheat (TriticumaestivumL.) [J].TheoreticalandAppliedGenetics,2008,117:701-709.
[6]FU X Y,QI Z L,LI S S.QTL detection for water-soluble oligosaccharide content of grain in common wheat (TriticumaestivumL.) [J].MolecularPlantBreeding,2011,2(10):68-74.
[7]REBETZKE G J,VAN HERWAARDEN A F,JENKINS C,etal.Quantitative trait loci for water-soluble carbohydrates and associations with agronomic traits in wheat (TriticumaestivumL.) [J].AustralianJournalofAgriculturalResearch,2008,59:891-905.
[8]LEHMENSIEK A,ECKERMANN P J,VERBYLA A P,etal.Flour yield QTL in three Australian doubled haploid wheat populations (TriticumaestivumL.) [J].AustralianJournalofAgriculturalResearch,2006,57:1115-1122.
[9]LEHMENSIEK A,ECKERMANN P J,VERBYLA A P,etal.Curation of wheat maps to improve map accuracy and QTL detection(TriticumaestivumL.) [J].AustralianJournalofAgriculturalResearch,2005,56:1347-1354.
[10]杨 莉,黄玉莲,常 萍,等.小麦阿拉伯木聚糖含量的QTL分析及其与品质性状的关系[J].作物学报,2014,40(9):1695-1701.
YANG L,HUANG Y L,CHANG P,etal.QTL mapping for arabinoxylans content and its relationship with processing quality in common wheat [J].ActaAgronomicaSinica,2014,40(9):1695-1701.
[11]梁 恒.小麦戊聚糖含量的QTL定位及迟熟α-淀粉酶鉴定[D].雅安:四川农业大学,2011:21-25.
LIANG H.QTL mapping for pentosan content and identification of late maturity α-amylase in common wheat [D].Ya’an:Sichuan Agricultural Uniersity,2011:21-25.
[12]ARCADE A,LABOURDETTE A,FALQUE M,etal.BioMercator:Integrating genetic maps and QTL towards discovery of candidate genes [J].Bioinformatics,2004,20(14):2324-2326.
[13]CHARDON F,VIRLON B,MOREAU L,etal.Genetic architecture of flowering time in maize as inferred from quantitative trait loci meta-analysis and synteny conservation with the rice genome [J].Genetics,2004,168:2169-2185.
[14]方永丰,李永生,白江平,等.玉米持绿相关QTL整合图谱构建及一致性QTL区域内候选基因发掘[J].草业学报,2012,21(4):175-185.
FANG Y F,LI Y S,BAI J P,etal.Integration map construction and identification candidate gene in consensus QTL for chlorophyll factors in maize based on bioinformatics [J].ActaPrataculturaeSinica,2012,21(4):175-185.
[15]吴 琼,齐照明,陈庆山,等.基于元分析的大豆生育期QTL的整合[J].作物学报,2009,35(8):1418-1424.
WU Q,QI Z M,CHEN Q S,etal.An integrated QTL map of growth stage in soybean constructed through meta-analysis [J].ActaAgronomicaSinica,2009,35(8):1418-1424.
[16]SOMERS D J,FEDAK G,SAVARD M.Molecular mapping of novel genes controllingFusariumhead blight resistance and deoxynivalenol accumulation in spring wheat [J].JournalofGeneticsandGenomics,2003,46(4):555-564.
[17]栗文娟,刘志斋,石云素,等.基于元分析和生物信息学分析的玉米抗旱相关性状QTL一致性区间定位[J].作物学报,2010,36(9):1457-1467.
LI W J,LIU Z Z,SHI Y S,etal.Detection of consensus genomic region of QTL relevant to drought-tolerance in maize by QTL meta-analysis and bioinformatics approach [J].ActaAgronomicaSinica,2010,36(9):1457-1467.
[18]王帮太,吴建宇,丁俊强,等.玉米产量及产量相关性状QTL的图谱整合[J].作物学报,2009,35(10):1836-1843.
WANG B T,WU J Y,DING J Q,etal.Map integration of QTLs for grain yield and its related traits in maize [J].ActaAgrnomicaSinica,2009,35(10):1836-1843.
[19]王健维,程宇坤,叶雪玲,等.小麦品质相关性状的一致性数量性状位点(MQTL)连锁图谱构建[J].农业生物技术学报,2015,23(5):671-682.
WANG J W,CHENG Y K,YE X L,etal.Construction of linkage map of the meta quantitative trait loci(MQTL) on quality related traits in wheat(TriticumaestivumL.) [J].JournalofAgriculturalBiotechnology,2015,23(5):671-682.
[20]TUBEROSA R,SALVI S,SANGUINETI M C,etal.Mapping QTL regulating morpho-physiological traits and yield:case studies,shortcomings and perspectives in drought-stressed maize [J].AnnalsBotany,2002,89:941-963.
[21]齐照明,孙亚南,陈庆山,等.基于Meta分析的大豆百粒重的QTL定位[J].中国农业科学,2009,42(11):3795-3803.
QI Z M,SUN Y N,CHEN Q S,etal.Meta-analysis of 100-seed weight QTL in soybean [J].ScientiaAgriculturaSinica,2009,42(11):3795-3803.
[22]VPPRRIPS R E.MapChart:Software for the graphical presentation of linkage maps and QTL [J].Heredity,2001,93:77-78.
[23]LI X H,LI X H,HAO Z F,etal.Comparative QTL mapping of resistance to sugarcane mosaic viruses maize based on bioinformatics [J].Hereditas,2008,30(1):101-108.
Integration and Meta-analysis of Quantitative Trait Loci for Water-Soluble Carbohydrate Content in Wheat Grain
HU Yajun1,LI Mengfei1,YANG Delong1,LIU Yuan1,CHENG Hongbo1,CHANG Lei2,CHAI Shouxi2
(1.Gansu Provincial Key Lab of Aridland Crop Science/College of Life Science and Technology,Gansu Agricultural University,Lanzhou,Gansu 730070,China; 2.College of Agronomy,Gansu Agricultural University,Lanzhou,Gansu 730070,China)
Under drought stress conditions,water-soluble carbohydrates (WSC) of vegetative organs in wheat are an important source of carbon required for wheat grain filling. To explore true and major quantitative trait loci (QTL) for WSC content in wheat grain,a total of 168 QTL controlling grain WSC content from different genetic mapping populations in wheat were employed to establish consensus map and to perform meta-analysis by the method of bioinformatics,depending on a high-density genetic map as a reference map. There were 142 QTL overlapped the same marker intervals with the reference map.Of them,ninty-two QTL had positive effects on phenotypic variations of grain WSC content in wheat,whereas 50 QTL showed negative effects. A consensus genetic map for QTL controlling grain WSC content was established. Furthermore,sixteen meta QTL (MQTL) and their tightly linked markers were confirmed,with the minimum confidence interval of MQTL reaching to 0.77 cM.The findings might provide theoretical basis for fine mapping and map cloning of QTL controlling grain WSC content in wheat.
Wheat; Water-soluble carbohydrates; Meta quantitative trait loci; Meta-analysis
2016-03-09
2016-04-21
国家自然科学基金项目(31460348;30960195);陇原青年创新人才扶持计划项目;甘肃省农业生物技术研究与应用开发项目(GNSW-2015-18);甘肃农业大学“伏羲人才”计划项目(FXRC20130102)
E-mail:hyj9800@163.com
杨德龙(E-mail:yangdl@gsau.edu.cn)
S512.1;S330
A
1009-1041(2016)08-0989-07
网络出版时间:2016-08-01
网络出版地址:http://www.cnki.net/kcms/detail/61.1359.S.20160801.1120.006.html
猜你喜欢
内江师范学院学报(2022年4期)2022-04-27
现代畜牧科技(2021年4期)2021-12-05
现代畜牧科技(2021年10期)2021-11-19
湖北师范大学学报(自然科学版)(2021年3期)2021-09-08
数学物理学报(2021年1期)2021-03-29
现代装饰(2020年7期)2020-07-27
铁道通信信号(2018年9期)2018-11-10
河北农业科学(2018年2期)2018-07-26
NBA特刊(2018年7期)2018-06-08
中国粮油学报(2016年5期)2016-01-23
