
杂拟谷盗触角感器的扫描电镜观察
2016-09-14 06:14唐培安吴海晶孔德英
植物保护 2016年1期
唐培安, 吴海晶, 孔德英, 薛 昊, 宋 伟
(1. 南京财经大学食品科学与工程学院, 江苏省现代粮食流通与安全协同创新中心,江苏高校粮油质量安全控制及深加工重点实验室, 南京 210023; 2. 重庆出入境检验检疫局, 重庆 400020)
杂拟谷盗触角感器的扫描电镜观察
唐培安1*,吴海晶1,孔德英2,薛昊1,宋伟1
(1. 南京财经大学食品科学与工程学院, 江苏省现代粮食流通与安全协同创新中心,江苏高校粮油质量安全控制及深加工重点实验室, 南京210023; 2. 重庆出入境检验检疫局, 重庆400020)
利用扫描电镜对杂拟谷盗成虫触角形态及感器进行了观察,结果表明,杂拟谷盗成虫触角为棒形,由柄节、梗节和鞭节组成,其中鞭节又分为9个亚节,不存在雌雄二型现象。雌、雄成虫触角上均存在以下6类14种感器,包括Böhm氏鬃毛、3种毛形感器、1种栓锥形感器、6种锥形感器、1种刺形感器、2种指形感器。雌雄个体之间触角感器的类型、分布、长度、基部直径没有明显差异。结合感器的形态、分布和已报道的触角电位反应数据,对杂拟谷盗触角各类感器的功能进行了推测。
杂拟谷盗;触角;感器;扫描电镜
触角是昆虫感觉系统的重要组成部分,通过触角上不同类型的感器可行使嗅觉、味觉及感受气流、二氧化碳、湿度和温度等功能,从而调节昆虫的行为适应当下的环境条件[1-2]。感器是昆虫感受器官最基本的结构单元,由触角表皮特化而形成,是昆虫机体感知内外环境,进行化学通讯的接受体[3]。昆虫感器种类众多,不同昆虫其感器的分布、形态和功能也不同。研究昆虫的感器对于探讨昆虫感受外界信息的机制具有重要意义[4]。
杂拟谷盗(Tribolium confusumJacquelinduVal),又称“杂拟谷甲”、“广颊谷蛀”,隶属于鞘翅目(Coleoptera),拟步甲科(Tenebrionidae),是一种全世界分布的重要储藏物害虫,国内各地均有发生。杂拟谷盗食性复杂,主要为害各种粮食、油料、豆类、糠麸、毛皮、药材及其他储藏物品,其中以面粉、玉米、油料受害最严重。成虫有臭腺,能分泌臭液,大量发生时,易使被害物发生霉臭味。被害面粉结块、色污、味臭、不能食用,严重威胁储藏物的安全[5-6]。近年来,扫描电镜技术(SEM)在昆虫学研究领域备受关注,并在昆虫触角感器研究中得到了广泛应用[7]。因此本研究应用扫描电镜观察,鉴定杂拟谷盗触角上的感器种类、形状和分布密度,为进一步运用电生理技术研究触角感器系统在化学感受机理中的作用奠定基础。
1 材料与方法
1.1供试昆虫
供试昆虫杂拟谷盗采自中央储备粮三明直属库,并在养虫室内人工饲养数代。用全麦粉∶干酵母粉=20∶1(质量比)混合而成的饲料,在温度为(28±1)℃、湿度为(75±5)%、24h无光照条件的恒温培养箱中饲养。
1.2试验试剂
戊二醛(C5H8O2),国药集团化学试剂有限公司;叔丁醇(C4H16O),中国医药集团上海化学试剂公司;乙腈(CH3CN),天津市大贸化学试剂厂。
1.3试验仪器
超声波清洗器:KH5200B型,昆山禾创超声仪器有限公司;扫描电镜:TM3000,日本株式会社日立那珂有限公司。
1.4试验方法
昆虫触角的处理参考孙红霞等[7]的方法,并稍作修改。随机选取雌、雄杂拟谷盗各5头,浸泡在0.1mol/L磷酸缓冲液中超声波处理30s,再用超纯水清洗3次。立即放入2.5%戊二醛中固定2~3h,再用0.1mol/L磷酸缓冲液和超纯水各清洗3次。然后依次用30%、50%、60%、70%、80%、90%乙醇和无水乙醇进行梯度脱水。最后以50%叔丁醇、70%叔丁醇、100%叔丁醇、叔丁醇-乙腈(2∶1)、叔丁醇-乙腈(1∶1)、100%乙腈进行梯度干燥。
将触角用导电双面胶背向粘在样品台上,采用离子溅射器喷金,应用TM3000扫描电镜在15kV加速电压下观察和摄影。触角感器类型的形态学鉴定参照Schneider[4]的方法。长度测量采用显微图像分析测量软件Digimizer3.2。同一类型的感器至少选择15张照片测量其长度和基部直径,取其平均数。
1.5数据处理
所有数据采用SPSS16.0进行计算和统计分析。采用独立样本t检验法比较雌雄间触角各节长度、直径的差异。
2 结果与分析
2.1触角的一般形态
杂拟谷盗雌雄成虫触角形态相似,均呈棒状(图1a~b),共11节,柄节(scape,Sc)圆柱形,较粗壮;梗节(pedicel,Pe)较柄节细短;鞭节(flagellum,F)由9 个亚节组成。除雌虫触角鞭节第2、6、8亚节长度、第1、7亚节的端部直径显著大于雄虫外,雌雄成虫触角其余各节长度及端部直径和基部直径、总长度均无显著性差异(P>0.05)。杂拟谷盗与赤拟谷盗形态相似,两者的区别为:杂拟谷盗鞭节末端3亚节直径、长度逐渐膨大成棒形,而赤拟谷盗末端3节突然膨大成锤状[8]。鞭节第9亚节似球形,为鞭节中最粗壮亚节。触角表面除鞭节末端端部光滑外其余各节表面均呈鳞片状。杂拟谷盗雌雄成虫触角各节长度、端部、基部直径数据见表1~3。
表1杂拟谷盗成虫触角各节平均长度1)
Table1MeanlengthofantennalsegmentsofTribolium confusum
μm
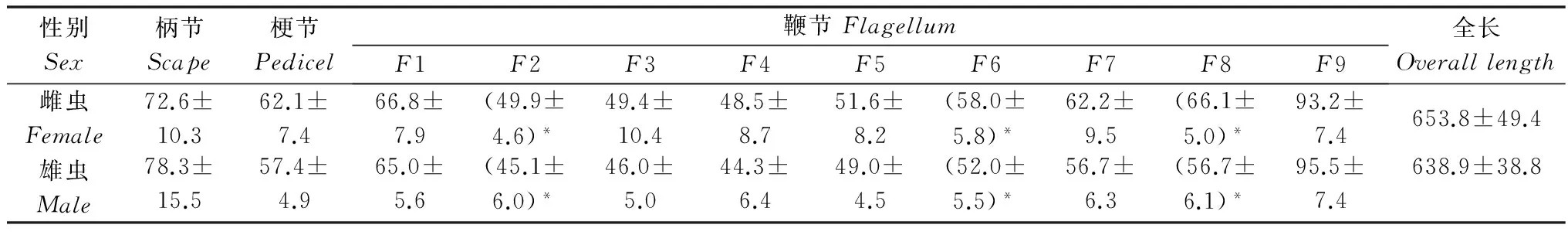
性别Sex柄节Scape梗节Pedicel鞭节FlagellumF1F2F3F4F5F6F7F8F9全长Overalllength雌虫Female72.6±10.362.1±7.466.8±7.9(49.9±4.6)*49.4±10.448.5±8.751.6±8.2(58.0±5.8)*62.2±9.5(66.1±5.0)*93.2±7.4653.8±49.4雄虫Male78.3±15.557.4±4.965.0±5.6(45.1±6.0)*46.0±5.044.3±6.449.0±4.5(52.0±5.5)*56.7±6.3(56.7±6.1)*95.5±7.4638.9±38.8
1) 表中数据为平均值±标准误,*表示雌雄虫触角该节长度有显著差异(P<0.05)。下同。
Datainthistablearemean±SE.Theasteriskindicatesignificantdifferenceinthevaluesofantennalsegmentsbetweensexesbasedont-testat0.05level.Thesamebelow.
2.2触角感器的种类、形态和分布
利用扫描电镜观察杂拟谷盗触角感器,发现有14种感器,包括Böhm氏鬃毛、毛形感器(Ⅰ、Ⅱ、Ⅲ)、栓锥形感器、锥形感器(Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅴ、Ⅵ)、刺形感器和指形感器(Ⅰ、Ⅱ)。感器呈环状排列,柄节存在于触角窝中,其上感器分布较少,其余节上触角感器的种类和数量从柄节到鞭节逐步增加,主要集中在鞭节第6~9亚节上。触角末节端部感器呈菊花状分布,囊括了除Böhm氏鬃毛外的所有感器类型,排列紧密,密度较大。
表2杂拟谷盗成虫触角各节端部直径
Table2DiameteroftheendregionsofantennalsegmentsofTribolium confusum
μm
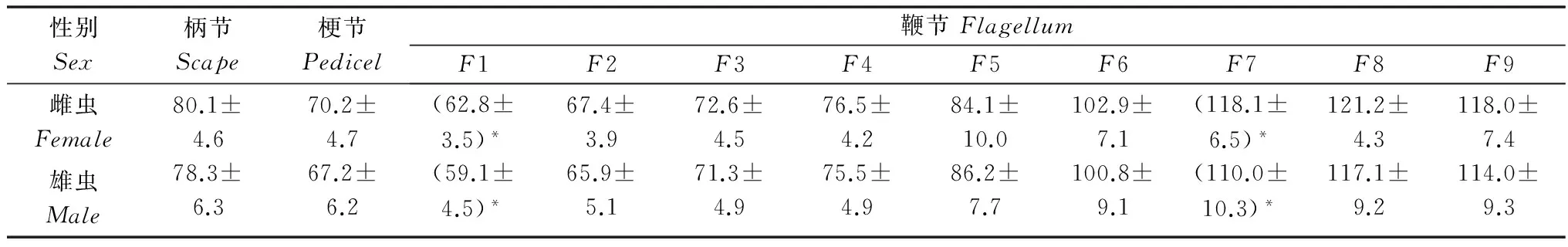
性别Sex柄节Scape梗节Pedicel鞭节FlagellumF1F2F3F4F5F6F7F8F9雌虫Female80.1±4.670.2±4.7(62.8±3.5)*67.4±3.972.6±4.576.5±4.284.1±10.0102.9±7.1(118.1±6.5)*121.2±4.3118.0±7.4雄虫Male78.3±6.367.2±6.2(59.1±4.5)*65.9±5.171.3±4.975.5±4.986.2±7.7100.8±9.1(110.0±10.3)*117.1±9.2114.0±9.3
表3杂拟谷盗成虫触角各节基部直径
Table3DiameteratthebaseofantennalsegmentsofTribolium confusum
μm

性别Sex柄节Scape梗节Pedicel鞭节FlagellumF1F2F3F4F5F6F7F8F9雌虫Female51.2±4.764.0±8.251.2±3.956.2±6.858.9±5.461.6±6.269.7±8.579.9±6.988.7±9.390.7±10.388.6±10.2雄虫Male45.8±3.363.0±5.251.7±4.657.5±1.560.4±3.667.0±5.574.4±7.780.7±8.786.5±14.391.1±14.288.6±11.9
2.2.1Böhm氏鬃毛
外形类似短刺,比刺形感器更短而尖,表面光滑,不具基窝(BB,图1c~d)。成簇分布于杂拟谷盗触角的柄节及柄节与梗节节间周围,长短不一。侧面和背面分布较多,鞭节上无此感器。基部直径1.1~1.7μm,长度7.5~11.5μm,数量较少。
2.2.2毛形感器
该类感觉器是杂拟谷盗触角上分布最多的感器,在柄节、梗节、鞭节均有分布。直立或略弯曲,端部尖细,着生部凹陷,密集着生于整个触角。根据其长度和形状,又可分为Ⅰ型、Ⅱ型和Ⅲ型。
毛形感器 Ⅰ 型(STⅠ,图1e~f):着生于凹陷的窝内,窝口平展。毛状体外形纤长,顶端最为尖细,特点是与触角方向平行,端部指向触角末端,感器上有明显的纵纹。基部直径 0.8~2.4μm,长度23.7~33.8μm。分布于杂拟谷盗触角每一节上。数量较多。
毛形感器Ⅱ型(STⅡ,图1g):着生于凹陷的窝内。直立或略弯曲,外形与STⅠ相似,顶端尖细,表面具明显纵纹,但比STⅠ短且细。基部直径0.6~1.3μm,长度9.0~18.1μm。普遍存在于杂拟谷盗触角鞭节的末端,在鞭节第6~8亚节近端部两侧也少量存在。
毛形感器Ⅲ型(STⅢ,图1h):着生于凹陷的窝内,窝口平展,端部略向外伸展,不与触角平行。外形与STⅠ相似,顶端尖细,表面具明显纵纹,但比STⅠ明显粗且长。基部直径 1.41~2.77μm,长度 35.4~40.0μm。仅分布于杂拟谷盗鞭节第5~8亚节近端部的两侧,数量较少。
2.2.3栓锥形感器
呈拇指状,着生于凹窝内。其表皮突起成栓状,端部有一锥状突起,表面有较浅的纵纹(SS,图1i)。基部直径 0.9~1.6μm,长度3.0~5.9μm。分布于鞭节末端的前缘位置,数量较多。
2.2.4锥形感器
末端钝圆,依据其形状差异分为6种:
锥形感器Ⅰ型(SBⅠ,图1j):着生于宽阔的凹穴中。圆台底座上的小锥分为2~4个分叉,基部联合,锥体光滑。底座直径3.4~5.5μm,底座部分至锥体顶端的长度9.0~12.2μm。主要集中在鞭节末端,鞭节第6~8亚节近端部的两侧也有分布,数量较少。
锥形感器Ⅱ型(SBⅡ,图1k):着生于较浅的凹穴内。类似于锥形感器Ⅰ上的分支小锥,上下粗细均匀,近端部有小锥,端部圆润,表面光滑。为锥形感器中最为粗壮的一种。基部直径1.5~3.0μm,长度7.0~15.6μm。集中分布于鞭节末端,少量发现于鞭节第6~8亚节近端部的两侧。
锥形感器Ⅲ型(SBⅢ,图1l):着生于较窄的凹窝内。直立细长,从基部到端部逐渐变细如锥状,端部较其他锥形感器略尖细,表面光滑。基部直径0.8~1.6μm,长度4.5~7.0μm。仅分布于鞭节末端,数量较少。
锥形感器Ⅳ型(SBⅣ,图1m):着生于较浅的凹陷窝内。短锥形感器,锥体很小,表面光滑。端部圆润,为所有感器中最短的一种,紧贴于表皮,不易被发现。基部直径0.8~1.6μm,长度1.2~2.4μm。分布于鞭节末端前缘上,数量较少。
锥形感器Ⅴ型(SBⅤ,图1n):着生于较深的凹穴内。短粗形圆锥体,可分为两部分,椎体下半部分为粗壮直立的柱体,端部钝圆为小锥体,呈乳头状突起,感器表面具较浅的纵纹。基部直径0.6~1.4μm,长度2.0~4.5μm。分布于鞭节末端前缘上,数量较少。
锥形感器Ⅵ型(SBⅥ,图1m):着生于较深的凹穴内。外形类似于锥形感器Ⅳ型,但比其短且极细,端部圆润,表面光滑。基部直径0.7~1.0μm,长度2.8~4.5μm。分布于鞭节末端,数量较少。
2.2.5刺形感器
刚直如刺,较为粗大,顶端钝圆。直立于触角表面,比其他感器明显高出许多,表面有明显的纵纹(SC,图1o~p)。基部直径 1.4~2.2μm,长度15.0~21.0μm。集中分布在鞭节的最后一节上。同时发现于鞭节第7亚节近端部的侧面,两侧各一根。
2.2.6指形感器
指形感器Ⅰ型(SFⅠ,图1q):着生于较深的凹穴内。长指形,高度与毛形感器Ⅱ相似,但端部直径比毛形感器明显粗,并呈钝圆状,与毛形感器区分明显。整体形似手指,上下粗细均匀,表面具较浅纵纹。基部直径0.7~1.4μm,长度5.5~9.5μm。集中分布在鞭节末端的前缘,数量较多。
指形感器Ⅱ型(SFⅡ,图1r):着生于凹穴内。短指形,与长指形形状相近,但大小差异显著。短指形感器粗壮且短小,端部圆润,表面具浅纵纹。基部直径1.1~1.6μm,长度2.8~3.8μm。仅在触角鞭节末端发现两三根,数量极少。
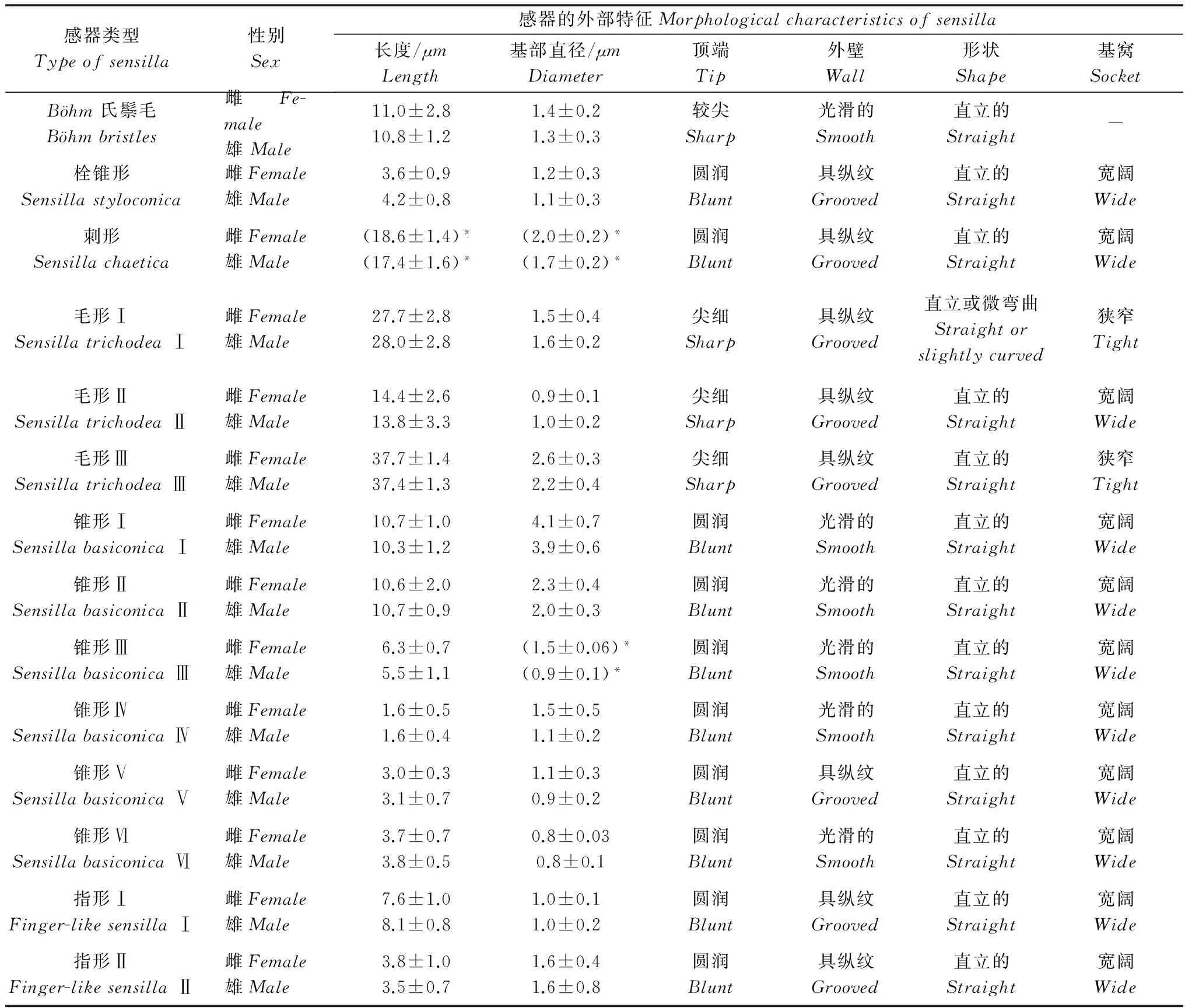
表4 杂拟谷盗触角感器的外部特征Table 4 Morphological characteristics of antennal sensilla of Tribolium confusum

图1 杂拟谷盗触角形态及感器类型Fig.1 Shapes and types of antenna and antennal sensilla of Tribolium confusum
2.3雌、雄虫触角感器比较
杂拟谷盗雌雄个体之间触角感器的类型、分布均不存在二型性。均具有Böhm氏鬃毛,毛形感器(Ⅰ、Ⅱ、Ⅲ)、栓锥形感器、锥形感器(Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅴ、Ⅵ)、刺形感器、指形感器(Ⅰ、Ⅱ)。触角各节表面均密布感器,但梗节和鞭节之间的节间膜没有感器。从柄节到鞭节的各亚节,感器类型越来越丰富,尤以鞭节末端节上的感器类型最多。虽然各类感器的分布位置在雌雄间没有差异,但各类感器的数量在雌雄间是否存在差异有待进一步观察。
对雌、雄成虫各类感器的基部直径和长度进行测量,结果表明,除雌虫刺形感器的基部直径和长度,锥形感器Ⅲ的基部直径显著大于雄虫外,其余各类感器的基部直径和长度均不存在显著性差异(P>0.05)。雌雄虫间比较数据见表4。
2.4触角感器的分布和数量
研究发现,杂拟谷盗每一种感器都有其特定的数量和分布位置。数量较多的是毛形感器,其次是锥形感器。毛形感器Ⅰ广泛分布于触角的每一节上。毛形感器Ⅱ主要分布鞭节末端,也发现于触角鞭节第6~8亚节近端部的两侧,数量较少。毛形感器Ⅲ数量为毛形感器中最少,仅在鞭节第5~8亚节端部的两侧发现几根。栓锥形感器仅分布于触角鞭节末端,数量较多。锥形感器(Ⅲ、Ⅳ、Ⅴ、Ⅵ)散布在触角鞭节的末端,其中锥形感器Ⅲ数量最多,其余类型数量较少。而锥形感器Ⅰ、Ⅱ分布在鞭节末端和第6~8亚节近端部的两侧,主要集中在鞭节末端,两侧数量极少。刺形感器集中分布在触角鞭节的末端,有10根左右,同时在鞭节的第7亚节近端部两侧各发现1根。指形感器分布于鞭节末端,Ⅰ型数量远远大于Ⅱ型。Böhm氏鬃毛仅成簇分布于柄节及柄节和梗节连接处。
3 讨论
昆虫触角感器在昆虫感受外界环境、实现信息交流方面具有重要意义。环境选择压力的变化及相互作用能够导致感器的数目及分布发生变化,其中重要的影响因素包括个体的大小、性别、食性、习性和栖境等[9]。
Böhm氏鬃毛在几乎所有的鞘翅目昆虫中均有分布,它是感受重力的机械感器,当遇到机械刺激时,能够缓冲重力的作用力,从而控制触角位置下降的速度[4]。毛形感器现已被发现具有触觉、嗅觉、味觉、机械感受等功能,是昆虫感受性信息素的主要器官[10-12]。杂拟谷盗的毛形感器Ⅰ型外部形态与许多报道的昆虫中毛形感器类似,如绿盲蝽、白背飞虱等[7,13]。诸多文献中膜翅目昆虫的毛形感器Ⅰ型被认为具有感受机械刺激的功能[14-15]。但毛形感器Ⅰ型的功能是嗅觉感受器已经在鞘翅目小蠹科[16]、象甲科[17]、天牛科[18]和其他科甲虫被证实或推测。因此,推测杂拟谷盗触角上的毛形感器Ⅰ型具有嗅觉感受功能。毛形感器Ⅱ型,与Bembidion properans[19]的毛形感器Ⅱ型外部形态很相似,被认为很可能是聚集信息素的接收器。毛形感器Ⅲ型分布在鞭节末端4节近端部两侧,接触不到表面,其功能可能为负责与其他昆虫的接触,有利于其防御天敌的进攻,因此推测毛形感器Ⅲ型具机械功能。
刺形感器底部较大的凹陷是其典型特点,其在触角上着生特点是直立于触角表面,并且比其他感器明显高出许多。刺形感器的这一形态特点,利于最先接触外界,因而具有感受机械刺激的功能,也可能在对寄主植物和异性近距离识别中发挥作用[17]。此外,刺形感器壁上无孔,且具有纵脊,是机械和味觉感器的特点[20-21]。
杂拟谷盗栓锥形感器与油菜蚤跳甲(Psylliodes chrysocephala)的栓锥形感器外部形态很相似,Bartlet等通过超微结构研究表明,其内部有丰富的神经细胞,且是触角内容物质的延伸,具有感受湿度、味觉和嗅觉功能[20],据此推测杂拟谷盗栓锥形感器具有与此相似的功能。
锥形感器被推测具有嗅觉、触觉功能[22-23]。杂拟谷盗锥形感器Ⅰ型为分叉锥形感器,与青杨脊虎天牛(Xylotrechus rusticus)的“锥形感器Ⅵ”[24]外形相近,推测为嗅觉感器。杂拟谷盗锥形感器Ⅱ型较为粗壮,且着生于末端前缘,易与外界接触,推断其可能为触觉感器。杂拟谷盗锥形感器Ⅲ型外部形态和桉嗜木天牛(Phoracantha semipunctata)的锥形感器Ⅱ[18]和沟眶象(Eucryptorrhynchus chinensis)触角感器的锥形感器b1[25]相似。Lopes[18]等通过对桉嗜木天牛电生理试验证明,锥形感受器具有识别植物气味分子的功能。据此推断,杂拟谷盗锥形感器Ⅲ型可能具有嗅觉感受功能。杂拟谷盗锥形感器Ⅴ型与红缘吉丁(Buprestis fairmairei)的锥形感器Ⅳ[26]形态相似,杂拟谷盗锥形感器Ⅵ型和Ⅳ型分别与东北大黑鳃金龟(Holotrichia diomphalia)锥形感器ba1和ba2[27]类似,研究者通过透射电镜扫描发现这些感器表皮为单壁,表皮上有许多微孔,分布在锥形感器的锥体部分表面有孔道贯穿于表皮和体腔之间,符合嗅觉感器的特征。因此推断杂拟谷盗锥形感器Ⅴ、锥形感器Ⅳ、锥形感器Ⅵ也为嗅觉感器。
有关指形感器的报道很少,在松褐天牛(Monochamus alternatus)触角上分布两种指形感器(为直形和弯形)[28],而在中华微蛾(Sinopticula sinica)触角上的指形感器为长指形和短指形[29]。指形感器具有明显的端部单孔,壁上无深刻纹。
鉴于不同感器的功能只在少数虫种中得以证实,并且感器类型和功能依据外部形态来划分并非总是一致,因此,对于杂拟谷盗触角上各类感器的功能,需进一步采用单细胞记录、触角电位等技术给予明确。
[1]王桂荣,郭予元,吴孔明. 棉铃虫触角感器的超微结构观察[J]. 中国农业科学, 2002, 35(12): 1479-1482.
[2]那杰,于维熙,李玉萍,等. 昆虫触角感器的种类及其生理生态学意义[J]. 沈阳师范大学学报(自然科学版), 2008, 26(2): 213-216.
[3]岳璐,刘星月,杨定. 四种鱼蛉触角感器的超微形态[J]. 植物保护, 2014, 40(5): 62-69.
[4]SchneiderD.Insectantennae[J].AnnualReviewofEntomology, 1964, 9(1): 103-122.
[5]王阿旻,程超,潘沈元,等. 体内沃尔巴克氏体(Wolbachia)对寄主杂拟谷盗生殖的调控作用[J]. 江苏农业学报, 2014, 30(1): 47-52.
[6]CampbellJF,ArthurFH,MullenMA.Insectmanagementinfoodprocessingfacilities[J].AdvancesinFood&NutritionResearch, 2004, 48: 239-295.
[7]孙虹霞,胡新军,舒迎花,等. 白背飞虱触角感器的扫描电镜观察[J]. 昆虫学报, 2006, 49(2): 349-354.
[8]胡飞. 十种储藏物害虫触角感器的超微结构研究[D]. 重庆: 西南大学, 2009.
[9]李竹,陈力. 触角感器特征应用于昆虫分类的研究进展[J]. 昆虫分类学报, 2010, 32(1): 113-118.
[10]CastrejónGómezVR,NietoG,ValdesJ,etal.TheantennalsensillaofZamagiria dixolophellaDyar(Lepidoptera:Pyralidae)[J].AnnalsoftheEntomologicalSocietyofAmerica, 2003, 96(5): 672-678.
[11]陈湖海,康乐. 蝗虫触角感受器及其生态学意义[J]. 动物学杂志, 1998, 33(3): 46-49.
[12]DolzerJ,FischerK,StenglM.Adaptationinpheromone-sensitivetrichoidsensillaofthehawkmothManduca sexta [J].JournalofExperimentalBiology, 2003, 206(9): 1575-1588.
[13]陆宴辉,仝亚娟,吴孔明. 绿盲蝽触角感器的扫描电镜观察[J]. 昆虫学报, 2007, 50(8): 863-867.
[14]OnagbolaEO,FadamiroHY.ScanningelectronmicroscopystudiesofantennalsensillaofPteromalus cerealellae (Hymenoptera:Pteromalidae)[J].Micron, 2008, 39(5): 526-535.
[15]PetterssonEM,HallbergE,BiggerssonG.Evidencefortheimportanceofodour-receptionintheparasitoidRhopalicus tutela (Walker) (Hymenoptera:Pteromalidae)[J].JournalofAppliedEntomology, 2001, 125(6): 293-301.
[16]FaucheuxMJ.Distributionandabundanceofantennalsensillafromtwopopulationsofthepineengraverbeetle, Ips pini (Say) (Coleoptera:Scolytidae)[J].AnnalesdesSciencesNaturelles-ZoologicetBiologieAnimale, 1994, 15(1): 15-31.
[17]SaïdI,TaubanD,RenouM,etal.StructureandfunctionoftheantennalsensillaofthepalmweevilRhynchophorus palmarum (Coleoptera:Curculionidae)[J].JournalofInsectPhysiology, 2003, 49(9): 857-872.
[18]LopesO,BarataEN,MustapartaH,etal.Finestructureofantennalsensillabasiconicaandtheirdetectionofplantvolatilesintheeucalyptuswoodborer, Phoracantha semipunctataFabricius(Coleoptera:Cerambycidae)[J].ArthropodStructure&Development, 2002, 31(1): 1-13.
[19]MeriveeE,PloomiA,RahiM,etal.AntennalsensillaofthegroundbeetleBembidion properansSteph. (Coleoptera:Carabidae)[J].Micron, 2002, 33(5): 429-440.
[20]BartletE,RomaniR,WilliamsIH,etal.FunctionalanatomyofsensorystructuresontheantennaeofPsylliodes chrysocephalaL. (Coleoptera:Chrysomelidae)[J].InternationalJournalofInsectMorphologyandEmbryology, 1999, 28(4): 291-300.
[21]王焱,穆兰芳,曾凡荣,等. 樟巢螟成虫触角感器的扫描电镜观察[J]. 应用昆虫学报, 2011, 48(3): 675-679.
[22]李科明,张永军,吴孔明,等. 中红侧沟茧蜂触角超微结构[J]. 中国农业科学, 2012, 45(17): 3522-3530.
[23]陈丽,陈科伟,梁广文. 夜蛾黑卵蜂雌蜂触角感器的扫描电镜观察[J]. 华南农业大学学报, 2013, 34(1): 72-75.
[24]程红,严善春,徐波,等. 青杨脊虎天牛触角主要感器的超微结构及其分布[J]. 昆虫知识, 2008, 45(2): 223-232.
[25]杨贵军,张大治,孙晶莹. 沟眶象触角感器的扫描电镜观察[J]. 昆虫知识, 2008, 45(6): 926-931.
[26]刘玉双,石福明. 红缘吉丁(鞘翅目: 吉丁虫科)触角感器的扫描电镜观察[J]. 昆虫学报, 2005, 48(3): 469-472.
[27]孙凡,胡基华,王广利,等. 东北大黑鳃金龟嗅感器超微结构[J]. 昆虫学报, 2007, 50(7): 675-681.
[28]王四宝,周弘春,苗雪霞,等. 松褐天牛触角感器电镜扫描和触角电位反应[J]. 应用生态学报, 2005, 16(2): 317-322.
[29]高素红,吉志新,王长青,等. 中华微蛾(Sinopticula sinicaYang)触角感器的扫描电镜观察[J]. 安徽农业科学, 2010, 38(7): 3499-3502.
(责任编辑:杨明丽)
ScanningelectronmicroscopicobservationonantennalsensillaofTribolium confusum
TangPeian1,WuHaijing1,KongDeying2,XueHao1,SongWei1
(1.CollegeofFoodScienceandEngineering,JiangsuCollaborativeInnovationCenterforModernGrainCirculationandSafety,KeyLaboratoryofGrainsandOilsQualityControlandProcessing,NanjingUniversityofFinanceandEconomics,Nanjing210023,China; 2.ChongqingEntryExitInspectionandQuarantineBureau,Chongqing400020,China)
AntennalmorphologyandthesensillaofTribolium confusumJacquelinduValwerestudiedbyscanningelectronmicroscopy.Theresultsshowedthattheantennaearemadeupofscape,pedicelandflagellumthatconsistedof9segmentsandtherearenodimorphismsontheantennaeofthemaleandfemaleadults.Furthermore,thereare6kindsand14typesofsensillaontheantennaeofbothadults,includingBöhmbristles, 3typesofsensillatrichodea, 1typeofsensillastyloconica, 6typesofsensillabasiconica, 1typeofsensillachaetica,and2typesoffinger-likesensilla.Nodifferencewasfoundamongthetypes,thedistribution,thenumber,lengthanddiameterofsensillabetweenbothadults.Thefunctionsofsensillawerespeculatedaccordingtotheirmorphology,distribution,andthepreviouslyreportedelectroantennographicresponsesofTribolium confusum.
Tribolium confusum;antenna;sensilla;scanningelectronmicroscopy
2014-11-28
2015-01-21
“十二五”国家科技支撑计划项目(2013BAD17B01-3);粮食公益性行业科研专项(201413007-2);国家自然科学基金(31000828);江苏高校优势学科建设工程项目
E-mail:tangpeian@163.com
Q964
ADOI:10.3969/j.issn.0529-1542.2016.01.017
猜你喜欢
大电机技术(2022年4期)2022-08-30
大电机技术(2022年1期)2022-03-16
新疆钢铁(2021年1期)2021-10-14
生物学杂志(2021年4期)2021-08-24
昆明医科大学学报(2021年1期)2021-02-07
铁道通信信号(2020年10期)2020-02-07
河南农业科学(2019年5期)2019-05-28
植物保护(2018年3期)2018-05-14
中国水稻科学(2017年2期)2017-04-05
中华老年口腔医学杂志(2016年5期)2016-03-01
