
被子植物和它们的昆虫伙伴
2016-08-30 17:12杨斧
大自然探索 2016年8期
当我们在森林、原野中或公园里漫步时,见到最多的生物就是能够开花结果的被子植物和与它们如影相随的昆虫了。它们不但种类繁多而且种群庞大,令人目不暇接。在本刊2016年6月刊,我们已经见识了具有远古历史和庞大群体的昆虫帝国,下面,让我们再跟随植物学家杨斧老师来了解一下植物与昆虫的协同进化。
迄今为止,植物分类学家已经记录在案的被子植物有近30万种,昆虫分类学家已记录到的昆虫多达近100万种,而据保守的估计,地球上现有的昆虫应该超过300万种。但是,关于这两大生物类群之间的关系,我们知道的还十分有限。近些年来,生态环境和生物多样性保护已成为人类的共识,然而,追溯漫长的自然演化历史和面对复杂纷纭的物种多样性,我们的知识仅仅是“沧海一粟”。尤其是由于博物学知识的苍白,当我们置身于森林或原野,面对众多生灵所编织的“生命之网”时,往往不知所措。下面,让我们来见识几种植物和昆虫之间有趣的案例。
互惠互利的丝兰和丝兰蛾
在有花植物和昆虫协同进化时,双方均受益的互惠互利关系屡见不鲜。
丝兰是一类原产于美国西南部沙漠中的百合科(或龙舌兰科)植物,现在世界各地的园林、绿地中已普遍栽培,因此并不罕见。但它们与鳞翅目昆虫丝兰蛾之间上演的共同繁荣的一幕,只有在太平洋彼岸才能见到。
每年当洁白的丝兰花盛开时,丝兰蛾便在夜晚悄然而至。雌蛾先在一朵花上将散出的花粉聚在一起成圆球状,然后用前腿与头的下部配合,将花粉球抱走,安置在另一朵花的柱头凹陷处,这就完成了丝兰花的异花传粉过程。然后这只雌蛾将腹端对准接受花粉的雌蕊下部,用产卵器刺入子房,向其中产卵。在柱头上萌发的花粉管通过花柱分别进入子房中的胚珠,在胚囊中精子和卵细胞结合,完成授精过程。受精以后丝兰的子房发育:胚珠发育成种子,子房发育为果实。
与此同时,被雌蛾产入子房的卵,也孵化出幼虫,营养丰富的丝兰幼嫩种子,正好成为丝兰蛾幼虫的美味佳肴,而曾经作为产房的丝兰花子房,又成了丝兰蛾幼虫包吃包住的“保育院”。尽管丝兰蛾幼虫的食量很大,但每一朵丝兰花的子房中有200个胚珠,如果都能受精的话,就有同样多的种子供丝兰蛾幼虫食用,而丝兰蛾在每一个子房中产卵4~5枚,幼虫只能吃掉30%~50%的种子,剩下的种子足够丝兰繁殖后代的需要。
丝兰蛾为丝兰异花传粉,丝兰为丝兰蛾幼虫提供产房、保育院和食物,这种互惠互利,成为植物与昆虫专性共生的典范,常见于博物学书刊之中。但毕竟丝兰属的种类不多,原产地范围有限,比起分布范围广泛、种类繁多的榕树和授粉昆虫的共生关系,可谓小巫见大巫了。
榕树离不开虫瘿
榕树和丝兰相似,也以果实为传粉昆虫提供住所,但在榕树的“虫房”中,昆虫不但生儿育女,而且幼虫变为成虫后,还要在榕树提供的“虫房”中谈婚论嫁。而榕树这个“房东”也因此得到繁衍后代的机会,在生物演化的历程中与共生昆虫携手并进。
在亚洲南部,包括我国华东南部经华南到西南的广大地区,榕树是一类常见又异乎寻常的树种。在较空旷的地面上,榕树以庞大的树冠和独木成林的独特景观闻名遐迩。正如古人所说,榕树因为能够“容人”而得名。据说,印度加尔各答植物园中有一棵大榕树,可以同时为2万多人遮荫挡雨。榕树是桑科榕属树种,有1000余种,主要生长在热带雨林中,那里树种繁多,生存空间有限,榕树独自容人不那么容易了,但“容虫”却独具特色。
榕属树种虽然种类多,但共性明显,一般只要抓住三点就可以了:一是木本;二是具有乳汁(这也是桑科的共性);三是结“无花果”。其中前两点并非榕属特有,但第三点在自然界中却鲜有雷同者。
榕树都是靠昆虫授粉的植物,然而,在年雨量2000~4000毫米甚至高达5000毫米的环境下,昆虫要露天工作就会遇到很大麻烦。面对这一困境,在适者生存的演化过程中,榕树独辟蹊径,在花序上“大作文章”,形成了隐头花序——花序中的所有小花都被包藏于近于球形的花序托中,在“球”的前端仅留有一个小孔,供传粉昆虫进入。这样的花序结出的果实(其实是个果序),就是人们俗称的“无花果”。那么,榕树的授粉昆虫是谁?它们又是如何在隐头花序中工作和生活的?
现在,人们已经搞清榕树的授粉昆虫是一类被称为“榕小蜂”的膜翅目昆虫。一些学者还认为:榕树的隐头花序和“无花果”是一种特殊的“虫瘿”。
在观察自然的活动中,很多人都关注过虫瘿。这些在植物身上长出的怪模怪样的突起,往往让人感到莫名奇妙。其实虫瘿是植物与昆虫之间特殊作用的产物,是昆虫取食时所分泌或形成的某些促植物生长的化学成分引起的植物组织畸变。多数情况下,昆虫是得益者,它们从寄主植物身上获得丰富的食物和有良好保护作用的居室;而植物则消耗物质畸形生长,影响了正常的机能。但也有双方都受益的例外,榕树依靠造瘿昆虫——榕小蜂传授花粉,就是典型事例。
榕树是雌雄异花植物,其中雌花有两种类型:短花柱花和长花柱花,它们的花柱都不分裂;雄花只有一种类型:花柱较长,且一分为二。雌花一般位于隐头花序的下方。在两种雌花中只有长花柱者能够接受花粉,形成可以繁殖后代的果实;短花柱花是供榕小蜂产卵用的瘿花,由此类花中分别繁育出有翅膀的雌蜂和无翅膀的雄蜂,它们从短花柱花变成的虫瘿中出来后即进行交配。受精的雌蜂开始向花序的上部爬,途径雄花的领地时,收集成熟的花粉,并将其贮藏在胸部的囊中。
然后,雌蜂开始在花序接近顶部的地方,将花序的厚壁开出一条隧道,由此爬出花序。见到光明的雌蜂很快物色到一个没有被开出隧道的年轻花序,于是从花序顶部留出的小孔中爬进去。这个花序中雄花还没有发育,但下部的雌花已发育成熟,雌蜂在寻找短花柱花产卵时,用前足将胸囊中的花粉拨出送给长花柱花的柱头,完成了为榕树异花传粉的工作。同时用腹部长长的产卵器将卵产在短花柱花的子房中。完成交配的雄蜂和完成异花传粉及产卵的雌蜂分别结束生活史,死在不同的花序中。已受精的榕树雌花将发育成果实和种子,繁育出新的植株;榕小蜂则一代代地在榕树的隐头花序和“无花果”中繁衍生息。
榕树和榕小蜂的共生关系是被子植物与传粉昆虫协同进化共同繁荣的范例。目前已知世界上的每一种榕树都有一种特异的榕小蜂为其传粉,因此榕小蜂也成了膜翅目中的著名类群,有1000余种。
野牡丹为木蜂量体裁衣
榕树和榕小蜂之间配合默契的传粉适应,是在隐头花序中进行的,外界难以窥视。然而,在虫媒花和传粉昆虫的协同进化中,多数传粉伙伴间的交易,是开诚布公,显而易见的。
在我国南方较为常见的野牡丹科植物中,许多都能开出艳丽而奇特的花朵。说它们“艳丽”,是因为许多花的花冠呈现出艳红、粉红、紫红的色彩;而“奇特”,则因为它们的雄蕊不仅颜色有变化,而且形状也变得异乎寻常:在花中直立的黄色花药外,又出现了一种向四周伸展弯如镰刀头的紫色花药,而花的雌蕊花柱也追随镰刀型雄蕊向外伸出,例如野牡丹属的野牡丹、多花野牡丹、地菍,异药花属的异药花、肥肉草,野海棠属的野海棠、红毛野海棠等等。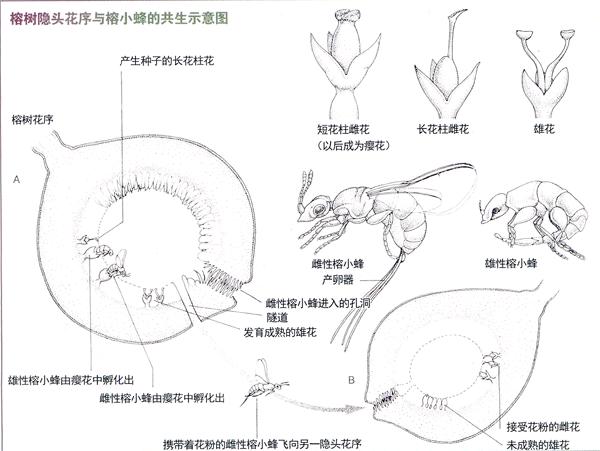
我国的园林工作者,在研究野牡丹科花卉多花野牡丹等的传粉生物学时发现:这类花的传粉者主要是一类以钻木为“专长”的木蜂,它们也喜食花粉以补充蛋白质等营养物质。当一只寻觅花粉的木蜂,飞临多花野牡丹上空时,位于紫红色花冠中部的黄色花药十分显眼,于是平落到花上,用口器对准花药取食花粉。由于木蜂体长达20多毫米,身体自然向外伸,正巧与向上翘起的镰刀形紫色花药的“镰刀头”接触,这时已经成熟的紫色花药端部孔裂,散出的花粉粘在了木蜂腹部。当这只木蜂再到另一朵多花野牡丹花上食黄色花药的花粉时,就将腹部刚“俘获”的紫色花药散出的花粉传给了雌蕊的柱头。
多花野牡丹花与木蜂的传粉适应,犹如量体裁衣,恰到好处。在野牡丹科中,其他具有长、短两型雄蕊的种类,与传粉昆虫是否也具有类似“量体裁衣”的适应方式,还有待进一步的观察和研究。
在自然界中,像多花野牡丹那样与传粉昆虫配合默契的被子植物不在少数,尤其是花的形态由较原始的辐射对称演化为较进化的两侧对称后,由于身体两侧对称的昆虫必须使身体和花朵的对称轴保持一致,才能钻进花朵将口器深入花的底部或距中吸食花蜜,这种默契变得更精彩。在双子叶植物中的唇形科和单子叶植物中的兰科,都不乏这种花与传粉昆虫协同进化的精彩表现。
唇形科鼠尾草属有著名的观赏花卉一串红和药用植物丹参等,我们可能并不陌生。这类两侧对称花的花冠上唇向上方拱起,下唇向前方平展,像一张企图吞食猎物的蟒蛇嘴。它的雌蕊花柱长长的成拱形,向上几乎贴着上唇伸出花冠,柱头二裂;两枚能育雄蕊的花丝,在下部具有一个位于花冠内壁上的支点,形成具有长臂和短臂的杠杆结构。
当一只与鼠尾草类花体形大小相适合的膜翅目蜜蜂科的蜂类,发现了鼠尾草花向它张开的花唇时,便向花冠口部飞来,落在平展的下唇上。再当它爬进花中吸食底部的花蜜时,就会触动雄蕊的下部,由于杠杆作用,长长的花丝向下弯垂,将花药触到拜访者多毛的背部,散出花粉。尝到甜食后,这只蜂退出这朵花,带着花托付给它的花粉再去另一朵同类花中解馋,这时在花冠上唇外等候的雌蕊柱头,刚好接收其背上携带的异花花粉。这只帮助鼠尾草花完成异花传粉任务的昆虫,犹如递送邮包的快递员,精准无误,但它并不白干,报酬是它渴望的甜食——蜜汁。
囚禁“媒人”的马兜铃花朵
北京西山的山谷中,经常可以见到一种长相有些古怪的草。它的身体柔软,不能独自站立,靠缠绕在别的草或小灌木上生长。在它开花时,见不到鲜艳的花瓣,也没有芳香的气味。一个黄绿色的口部偏斜的小喇叭样的东西,就是它的花。有时几只小喇叭凑在一起,像是鼓号队的号手相聚。这种草结出的果子有些像吊在马脖子上的铃铛。原来它是一种生活在北方的马兜铃家族成员——北马兜铃。
我们知道,北马兜铃有颇为奇特的外表,可能有人会问,这种异乎寻常的花朵如何接受“媒人”的访问,它有媒人吗?如果有,“媒人”又是谁呢?
马兜铃属是马兜铃科中的望族,有350多种,它们的花朵没有花萼和花冠的分化,仅有一个口部敞开的或弯或直的长管,雌蕊和雄蕊就深藏在长管的底部;尽管许多种类在管口处有较鲜艳的色彩,但花朵开放时不但不香,反而有股腐肉的臭味。一般来说,散发臭味的花朵中,没有引诱蝴蝶和蜜蜂的蜜汁,深藏的雄蕊又令喜食花粉的昆虫难以企及。看来,上门的“媒人”只能是既逐臭而来,又能钻进长管中的小虫了。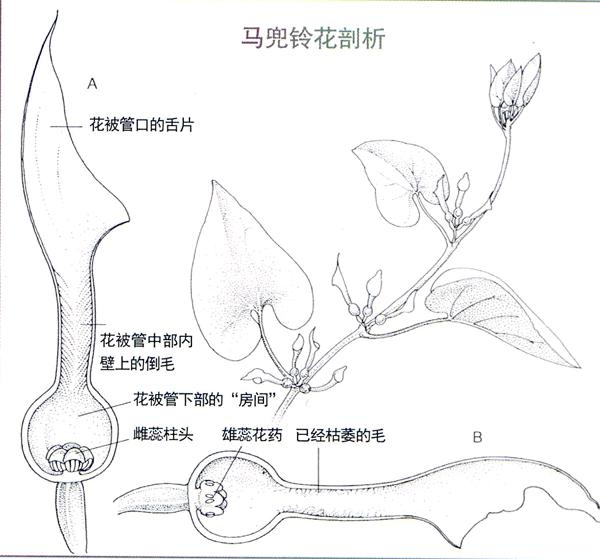
欧洲人自古就有喜好博物学的传统,他们不仅创建了生物分类学,而且对各类生物间的关系十分好奇。关于马兜铃属植物是如何传粉的,西方博物学家通过一种出产于欧洲等地的马兜铃——欧马兜铃,剖析了这类植物异花传粉的奥秘。
欧马兜铃的花和北马兜铃的花长得很像,花管口处有一片向一侧偏斜的长舌;花管的中部狭长,内壁上长满了尖头指向斜下方的刺毛;下部膨大成球形,中间生长有雄蕊和雌蕊。当欧马兜铃花开放时,一种喜欢臭味的双翅目蠓科小虫,逐臭而至。
在油滑的花管口内壁上,小蠓虫还没站稳,就滑了下去。在狭窄的花管内倒毛的领地,小虫更是像坐滑梯一样,懵懵懂懂地掉到了花管的底部。这里的内壁上没有毛,房间也较宽敞,但蠓虫没有找到发臭的腐烂物,在房子的中间却与聚在一起的雄蕊和雌蕊不期而遇。
马兜铃的花总是雌蕊比雄蕊先成熟。如果掉进来的小虫已经拜访过另一朵马兜铃花,身上一定粘有那朵花的花粉。当它失望地企图爬出花管时,管壁上的倒毛成了拦路虎。当它一次次向上爬,又一次次地被倒毛挡了回来时,身上的异花花粉就会不经意地传给雌蕊柱头,充当了传粉的“媒人”。
但已受精的马兜铃花仍不肯放走蠓虫,“拘留室”的“门”还要再关闭两三天,待自己的雄蕊成熟,花粉散出粘在小虫身体上后,那些把守细管通道的硬毛便枯萎了,小虫才能从容地爬出马兜铃的花朵。当这只“获释”的小蠓虫不记幽禁之苦,又被另一朵刚开放的马兜铃花释放的气味引诱,再遭拘留时,就又一次充当了马兜铃异花授粉的“媒人”。
博物学家指出,发生在欧马兜铃花内的异花传粉过程,在北马兜铃、马兜铃、管花马兜铃等同属植物中普遍存在,这也是被子植物与昆虫协同进化的事例之一。但是,这种关系看上去是不平等的,或者说是非互惠互利的关系。与此有些相似例子,在自然界中还有不少,其中既有比马兜铃更甚的“臭花”和“怪花”,也有令人愉悦的“香花”和“美花”。
聪明的模仿者——眉兰和火烧兰
春天,万物复苏。一只刚刚出世的雄角蜂,在地中海沿岸的草地上飞来飞去,寻找着自己的配偶。很快,在不远处一只雌角蜂出现了。它正一动不动地落在草丛上,似乎在静候雄蜂的到来。“就是它!”急于交配的雄蜂兴冲冲地落在了这只雌蜂的身上,用腿紧紧地抱住雌蜂那圆滚滚、毛绒绒、具有棕黄相间花纹的身躯,企图一同飞翔,完成空中交配的“婚飞”任务。但不知怎么回事儿,试了几次就是飞不起来,而自己的头上还粘上了一块黏乎乎的东西。这只雄蜂只得扫兴而去。很快,它又锁定了新的目标。但第二只雌蜂依然不配合,“婚飞”再次失败;头上那块黏东西却留在了第二只雌蜂那里。以后,这只不走运的雄蜂又在这片草地上如出一辙地经历了多次失败的求偶过程,最终筋疲力尽地飞走了。那些拒绝“婚飞”的“雌蜂”们却大功告成。
原来,草地上一只只飞不起来的“雌蜂”都是超级“模仿秀”,它们的真实身份是一种被称为“角蜂眉兰”的兰花的花朵,其多毛而胖胖的唇瓣酷似雌蜂的身体。角蜂眉兰乔装成雌蜂的目的就是诱骗雄蜂来为自己传播花粉,那块曾粘在雄蜂头上的东西,实际上是由许多角蜂眉兰花粉粘在一起形成的“花粉块”。
早在1862年出版的《兰花的传粉》一书中,博物学家达尔文就揭示了多种眉兰靠拟态实现异花传粉的过程。目前,博物学家已经发现从北非、西亚到南欧、西欧和中欧,分布着30多种靠拟态获得传粉机会的兰科眉兰属植物。这些眉兰像其他兰科植物一样,每一种都有自己特定的传粉者。其中,有膜翅目的土蜂、黄蜂、蜜蜂、角蜂家的成员,也有蝇类甚至蛛形纲的某种蜘蛛。科学家们新的研究表明,这些眉兰不仅在外形上模仿传粉者追逐的对象,而且还能通过释放化学物质,模拟特异传粉者的雌性个体向雄性发出的求爱信号——性信息素,以引诱其传粉者。而这种吸引传粉昆虫的化学信息的模拟,在兰科植物中颇为普遍。
与眉兰的传粉者不同,多数蜂类和蝴蝶迷恋芳香的花朵,是为了采集或吸食营养价值高的花粉和香甜的花蜜。但吸食蜜汁并非蜂、蝶的“专利”,许多另类昆虫也是甜食爱好者,具有刺吸式口器的同翅目蚜虫科的昆虫就是其中之一。食蚜蝇是蚜虫的天敌,它的幼虫专门以蚜虫为食。为了使自己的后代一出生就有充足的食物来源,“聪明”的雌性食蚜蝇根据一些种类的蚜虫有喜食蜜汁的习性,专门选择产蜜汁的花朵产卵。但有时“聪明反被聪明误”,当食蚜蝇遇到兰科中的气味模仿者——火烧兰属植物时,不但新出生的幼虫得不到美食,自己还要无偿地“为他人做嫁衣”。
高大火烧兰生长在北美西部的湿草地上,这里也是食蚜蝇活动的区域。像许多“吝啬”的兰花一样,高大火烧兰为了节约能源,并无蜜腺分泌蜜汁。为了同样得到传粉者的惠顾,模拟蜜汁的气味是这种兰花的拿手好戏。结果气味一出,为后代着想的食蚜蝇就立即响应,飞赴自以为是的幼虫“美食排档”,稳稳地降落在高大火烧兰唇瓣向前伸出的“平台”上。在兰花的唇瓣上转了几下后,食蚜蝇果断地产下一枚卵。与此同时,伸展在唇瓣上方的蕊柱也回赠给食蚜蝇一份对其毫无价值的“厚礼”——将花粉块粘到它的背上。食蚜蝇很有“生态学观念”,知道自己的后代应有合理的生存空间,不要让它们为争夺食物而进行生存竞争,于是在产下一枚卵后就飞走了。这也正是高大火烧兰求之不得的。只有当另一朵高大火烧兰的花让这只食蚜蝇再次上当而产下第二枚卵时,异花传粉过程才有可能完成。
在食蚜蝇与高大火烧兰之间因繁殖后代而结成的生态关系中,食蚜蝇是牺牲者,高大火烧兰是受益者。因为后者只是模拟了蜜汁的气味,并没有真的蜜汁分泌,自然也引不来喜食蜜汁的蚜虫。食蚜蝇的幼虫出生后就会遇到饥荒客死它乡,高大火烧兰的后代则因前辈拟态骗取的异花传粉机会而茁壮成长。
金合欢树和它的蚂蚁卫队
蚂蚁是地球上最牛的昆虫之一,这类膜翅目蚁科昆虫有1.4万种,别看它们的体形小,可数量极大,总重量不逊于人类的总重量!虽然多数种类的蚂蚁是肉食者,但在长期的进化历史中也与植物发生了千丝万缕的的联系,关乎许多植物类群的演化过程,并为陆生植物的扩散作出了不小的贡献。
在俗人眼里,踩死几只蚂蚁是小事一桩,但在热带地区,你要损害一棵与蚂蚁共生的树,可能会惹大麻烦。最典型的与蚂蚁结缘的树是金合欢。这是一类含羞草科金合欢属树种,有1200多种,广泛分布在热带及亚热带地区。它们只有几米高,树冠开张,具有二回羽状复叶,在茎上常有由托叶变成的成对的锐刺;在头状花序上有许多黄色小花,多数种的花有香味。
在非洲和热带美洲,金合欢树深受当地的蚂蚁青睐,是许多种蚂蚁栖身的“宝地”。蚂蚁们在树干上钻洞或将粗刺中心掏空变为自己的住所;而金合欢树羽状复叶基部分泌的蜜汁凝结成的蜜露和嫩叶尖上长出的富有营养的小颗粒,都是金合欢“房客”的美食。但这些蚂蚁房客并不白住、白吃,它们自觉地充当“房东”的卫士。当损害树木的切叶蚁或危害金合欢的其他昆虫来犯时,它们就会倾巢而出,将金合欢的敌人赶走或杀死,甚至一些较大型的食草动物也不例外。
不仅危害金合欢的动物会遭到蚂蚁卫士的凶猛攻击,就连妨碍金合欢树生长的别的植物,也会受到蚂蚁的攻击。这些蚂蚁房客会将金合欢周围距树干40厘米半径内的异种植物的枝叶咬碎,直到致其死亡。
早在100多年前博物学家就推断:这些在金合欢树上生活的蚂蚁,是被金合欢树当作一支“常备军”来供养的。以后博物学者的观察和试验也证实了这一观点。他们还观察到:不论白天、黑夜,蚂蚁卫队中都有四分之一左右的成员在“室外”执勤。而且除金合欢属树种外,还有大戟科、茜草科、野牡丹科和兰科的40多种植物都和蚂蚁有类似的共生关系,而所涉及的蚂蚁有蚁科中5个亚科的数百个蚁种。
在我国古代,南方的果农就已观察到:与柑橘树共生的黄猄蚁能大量捕食柑橘树上的害虫,护卫这类芸香科树木健康生长,被称为“橘园卫士”。另外,有一种体形较小的红蚂蚁,能捕食多种农业害虫,尤其喜欢消灭甘蔗螟虫。目前这两种蚂蚁均已被列为我国的天敌昆虫,受到保护。
科学家在总结植物和昆虫协同进化的关系时,认为不外乎以下三种:一是对双方都有利的互惠共生关系;二是对一方有利,对另一方有害的关系;三是对一方有利,对另一方无利,但也无损的关系。第一种关系无疑促进了双方的共同繁荣。第二、第三种关系,虽然看上去利益的天平是不平衡的,但在长期的演化过程中,不利方或无利方会通过变异来寻求无害和有利而得到发展。
以上虽然仅是被子植物和昆虫协同进化的几个事例,却也能让我们感知到保护生物多样性和生态环境的深远意义。
猜你喜欢
中国科学探险(2022年8期)2022-12-31
疯狂英语·新悦读(2022年12期)2022-03-08
昆虫学报(2021年5期)2021-07-07
新疆大学学报(自然科学版)(中英文)(2020年2期)2020-07-25
世界科学技术-中医药现代化(2019年7期)2019-10-22
浙江大学学报(农业与生命科学版)(2019年4期)2019-09-19
癌变·畸变·突变(2016年5期)2016-08-22
——凹脉马兜铃
广西植物(2016年4期)2016-05-27
中成药(2016年3期)2016-04-06
生态学报(2013年14期)2013-09-07
