
南麂岛潮间带软体动物多样性与群落结构
2016-08-29 01:57毕耜瑶蔡厚才陈万东俞存根夏陆军CheikhSarr
渔业研究 2016年2期
毕耜瑶,蔡厚才,陈万东,林 利,俞存根*,夏陆军,Cheikh Sarr,谢 旭
(1.浙江海洋大学水产学院,浙江 舟山 316024;2.南麂列岛国家海洋自然保护区管理局,浙江 平阳 325401)
南麂岛潮间带软体动物多样性与群落结构
毕耜瑶1,蔡厚才2,陈万东2,林利2,俞存根1*,夏陆军1,Cheikh Sarr1,谢旭1
(1.浙江海洋大学水产学院,浙江 舟山 316024;2.南麂列岛国家海洋自然保护区管理局,浙江 平阳 325401)
为揭示南麂岛潮间带软体动物多样性及其群落结构的现状,于2013年11月、2014年2月、5月、8月分四次对南麂岛的潮间带7个断面进行了定量和定性调查,采用传统多样性分析、群落聚类分析和非度量多维测度、ABC曲线相结合的方法,对软体动物的群落结构及其多样性进行研究。结果显示,(1)共鉴定出软体动物种类63种,隶属于3纲13目34科52属,不同季节的优势种及其优势度都有较大的差别,春、夏、秋、冬四个季节分别有4种、4种、5种和3种,条纹隔贻贝(Septifervirgatus)是四个季节共有且优势度最大的优势种;(2)软体动物群落结构组成的季节变化差异显著,春季、夏季、秋季、冬季组内相似性分别为30.77%、24.66%、19.58%、39.01%,春季-夏季、夏季-秋季、秋季-冬季、冬季-春季平均相异性分别为39.82%、39.00%、47.64%、48.02%;(3)软体动物Shannon-Wiener多样性指数(H′)四个季度平均值为冬季(1.73)>春季(1.69)>夏季(1.58)>秋季(1.40);(4)经聚类分析和非度量多维标度(NMDS)分析,该潮间带的软体动物群落可分为两组,主要是受生境不同的影响;(5)ABC曲线表明,春季、秋季、冬季软体动物群落未受到较大干扰,夏季受到中等强度的干扰。
软体动物;潮间带;多样性;群落结构;南麂岛
潮间带处于特殊的海陆交叉带,是海陆两大生态系统的生态交错带,可以提供无法替代的生态服务功能,是海洋生态系统生产力较高的区域[1]。潮间带海洋生物资源问题不仅是国外学者的研究重点[2-4],也是国内众多学者非常重视的领域。软体动物是潮间带一大生物群落,其生物多样性和群落结构的构成与潮间带生态系统息息相关,是整个海洋生态系统重要的一个环节。近年来,针对潮间带生物的研究有较多的报道,邹莉等[5]报道了关于浙江沿岸软体动物的种类分布情况,纪莹璐等[6]报道了乳山湾潮间带底栖动物群落结构,柯盛等[7]研究了雷州半岛流沙湾贝类的多样性,冷宇等[8]分析了黄河三角洲南部潮间带底栖生物的群落结构及多样性;有关软体动物的生态学方面,蔡林婷等[9]报道了舟山东极岛潮间带贝类的生态学状况,李荣冠等[10]主要针对鼓浪屿软相潮间带进行生态研究。
南麂列岛被誉为“贝藻王国”,是1990年经国务院批准建立的我国首批5个国家级海洋自然保护区之一。历史资料表明,南麂岛的气候比较适宜海洋生物的生长,四季分明,具有独特的地理位置和丰富的自然资源,使南麂列岛潮间带海洋生物颇具代表性和特殊性,又由于其受保护较好,成为研究潮间带生物的天然实验室,也成为许多学者的研究热点,如高爱根等[11-14]主要研究了自南麂列岛海洋自然保护区建区以来贝类的种类、数量、结构等变化,尤仲杰等[15-17]主要是在自己研究的基础上总结前人研究报道了南麂列岛贝类的种类及其群落结构,汤雁滨等[18-19]主要是针对南麂列岛底栖生物的生态位进行了研究,彭欣等[20]主要针对南麂列岛底栖动物群落结构进行了分析,高爱根等[21]还对人类活动与自然的关系进行了探讨。
此次对软体动物多样性和群落结构的研究,可摸清南麂岛软体动物的群落现状,不仅有利于软体动物的保护和开发,也可为管理部门制订有关南麂岛保护和开发的相关法规提供技术性支撑,因而在经济、科研以及管理方面都具有重要意义。
1 调查区域与方法
1.1调查区域

南麂岛作为南麂列岛的主岛,中心位置经纬度为27°27′N、121°05′E,西距大陆最近点约45 km,该海域属亚热带季风气候区,但具有明显的海洋性特征。潮间带生物调查分别于2013年11月(秋季)、2014年2月(冬季)、2014年5月(春季)、2014年8月(夏季)在当地大潮汛低潮期间进行,选择位于南麂岛的大山脚、大山脚砾石滩、国姓岙、火昆岙、马祖岙、三盘尾、山角寮7个断面取样,分别记为P1、P2、P3、P4、P5、P6、P7(图1),其中,除P2生态类型为砾石滩外,其余皆为岩礁。每条断面设置15个站位,分别为高潮区3个、中潮区9个、低潮区3个。1.2调查方法
在每个断面一般采集15个定量样方(高潮3个、中潮9个、低潮3个);采样框大小为0.25 m×0.25 m,采集样框内所有的生物样,并在每站附近区域广泛采集定性样品。部分区域因种类疏密程度及地势所迫,取样数量及采样框大小会有所调整。所有样品均放置于样品袋中并集中带回实验室进行分析鉴定,实验室中样品的采集、分析等均按照《GB/T12763.6-2007 海洋调查规范 第六部分:海洋生物调查》[22]进行。
数据处理及分析方法采用下列指数对南麂岛潮间带软体动物群落的优势种、物种多样性及结构进行分析:
1)物种优势度
物种的优势度(Y)计算公式[23]:
Y=(Ni/N)fi
(1)
式(1)中,Ni为第i种的个数(ind),fi为该种在各采样点出现的频率,N为所有物种出现的总个数(ind)。Y大于0.02的种类视为优势种。
2)多样性指数
采用Shannon-Wiener多样性指数(H′)、Pielou均匀度指数(J′)及Margalef物种丰富度指数(D)对软体动物群落进行多样性分析[24]。
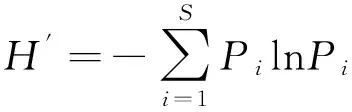
(2)
J′=H′/lnS;
(3)
D=(S-1)/lnM
(4)
式(2)(3)(4)中,S为总种数;M为所有种类的总生物量(g);Pi为第i种的生物量占总生物量的比例。
3)群落结构分析
对软体动物生物量采用对数转化[lg(x+1)],以降低少数优势种权重对数据分析的影响,使它们的分布更接近正态分布[25]。在Bray-Curtis相似性测量的基础上,也采用聚类和非度量多维标度(NMDS)进行群落结构分析,对这两种方法结果的准确性进行验证。采用相似性百分比(SIMPER)分析软体动物对组内相似性和组间相异性的贡献率。
4)ABC曲线
ABC(Abundance biomass comparison curve)曲线即丰度生物量曲线,在同一坐标系中,通过比较生物量优势度曲线和数量优势度曲线的走势情况来分析某一群落受到的干扰。如果生物量优势度曲线在数量优势度曲线之上,则表示群落稳定;若两条曲线相交,则表示群落受到中等程度干扰;若数量优势度曲线在生物量曲线之上,则表明群落受到严重干扰。W逐渐接近1时,表示群落生物量优势逐渐由单一物种所统治,但各物种的丰富度趋于相同;当W逐渐趋近于-1时,情况恰好相反[26]。
本文多样性指数、聚类分析、SIMPER、ABC曲线均用PRIMER 5.0软件绘制。
2 结果
2.1种类组成、优势种
根据本次调查所得的样品,经分析共鉴定出软体动物种类63种,隶属于3纲13目34科52属。各季节物种数分布不均,春季共鉴定出48种软体动物,夏季48种,秋季45种,冬季41种。依据此次调查结果得出,种类最多的出现在春、夏季,最少的出现在冬季,而且中、低潮区的物种数明显多于高潮区。根据相关的研究[27-28],将软体动物区系按温度性质可划分为广温广布种、温带种、亚热带种和热带种。其中亚热带种数量最多,达到30种;广温广布种次之,有29种;温带种和热带种都较少,都仅为2种。
条纹隔贻贝(Septifervirgatus)是各季度的第一优势种,春季、夏季、秋季、冬季的优势度分别达到0.31、0.29、0.17和0.42,但其他优势种的变化较大(表1)。其中,春季和夏季的优势种相同,都有4种,分别是条纹隔贻贝、短石蛏(Lithophagacurta)、疣荔枝螺(Thaisclavigera)和带偏顶蛤(Modioluscomptus);秋季优势种达到5种,除了条纹隔贻贝外,还有短滨螺(Littorinabrevicula)、日本菊花螺(Siphonariajaponica)、青蚶(Barbatiaobliquata)、疣荔枝螺;冬季优势种仅有3种,除条纹隔贻贝,还有疣荔枝螺和青蚶。四个季度共有的优势种是条纹隔贻贝和疣荔枝螺。

表1 不同季节软体动物优势种与优势度Tab.1 Dominance and dominant species of molluscs in different seasons
2.2时空分布
在季节变化上,从平均丰度来看,春季(676.0 ind/m2)>秋季(596.4 ind/m2)>冬季(435.2 ind/m2)>夏季(318.3 ind/m2);从平均生物量来看,春季(16 123.32 g/m2)>秋季(866.32 g/m2)>夏季(771.03 g/m2)>冬季(555.42 g/m2)。在空间分布上,P6不管在丰度还是生物量上都占有极大优势,而P3和P2都一直处在最末端。
2.3物种多样性
根据软体动物调查结果,做出四个季度Shannon-Wiener多样性指数(H′)及其偏差变化图(图2)。

春季Shannon-Wiener多样性指数(H′)平均值为1.69,最大值出现在P7和P3,达到2.24,最小值出现在P6,仅为0.40;夏季平均值为1.58,最大值出现在P7,达到2.37,最小值出现在P2,仅为0.86;秋季平均值为1.40,最大值出现在P4,达到2.08,最小值出现在P1,仅为0.46,;冬季平均值为1.73,最大值出现在P7,达到2.15,最小值出现在P3,仅为1.15。从Shannon-Wiener多样性指数(H′)平均值来看,冬季>春季>夏季>秋季。
2.4群落结构特征的季节变化
由表2可知春季、夏季、秋季、冬季四个季节组内相似性,其中春季组内平均相似性为30.77%,夏季为24.66%,秋季为19.58%,冬季为39.01%,冬季组内相似性最高,而秋季最低,春季、夏季、秋季均是以条纹隔贻贝贡献率最高,分别达到45.30%、50.54%、54.24%,冬季则是以短滨螺贡献率最高,达到35.42%。

表2 南麂岛潮间带四季组内平均相似性贡献率之和占90%以上的特征种Tab.2 The characteristic species for average similarity contribution rate that accounted for over 90% in the intertidal zone of Nanji Island for four seasons %
注:Av.Sim表示平均相似性,contrib表示贡献率。
Note:Av.Sim=average similarity.Contrib=contribution.
由表3可知春季-夏季、夏季-秋季、秋季-冬季、冬季-春季组间相异性。其中春季-夏季组间平均相异性为39.82%,夏季-秋季组间平均相异性为39.00%,秋季-冬季组间平均相异性为47.64%,冬季-春季组间平均相异性为48.02%,组间相异性最大的是冬季-春季,最小的是夏季-秋季。在四组组间相异性贡献率最大的种均是条纹隔贻贝,其贡献率分别为56.36%、68.54%、57.16%、68.54%。

表3 南麂岛潮间带四季组间平均相异性贡献率之和占90%以上的分歧种Tab.3 The discriminating species for average dissimilarity contribution rate that accounted for over 90% in the intertidal zone of Nanji Island between four seasons %
注:Av.Diss表示平均差异性.Contrib表示贡献率。
Note:Av.Diss=average dissimilarity. Contrib=contribution.
2.5群落划分
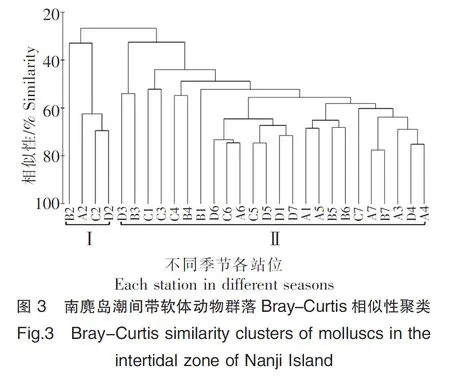
对每个季度的7个断面进行系统聚类和群落的非度量多维标度的二维分析,如图3和图4,聚类分析和MDS在群落生态研究中常用以评价群落结构[29-30]。其中字母A表示春季,B表示夏季,C表示秋季,D表示冬季,字母后数字代表该季节断面。对软体动物群落的聚类分析得出非常明显的分组,A2、B2、C2、D2在相似度为32.89%聚为一组,其余各点在相似度为32.49%聚为一组,两组在相似度为26.69%处相聚,两组的差异非常明显,这表明在四个季度中,P2和其余断面的软体动物群落差异都较大。

2.6不同季节ABC曲线的特征
根据南麂岛潮间带种类的丰度和生物量调查结果做出ABC曲线(图5),全年ABC曲线图显示,其W<0(W=-0.014),软体动物的群落结构受到较小程度的干扰。其中春季、秋季、冬季的ABC曲线显示都是生物量曲线位于丰度曲线之上,其W值分别为0.038、0.044、0.130(均大于0),表示春季、秋季、冬季的群落尚未受到较大的干扰,且这三个季度的生物量的优势度累积曲线起点均较高(在60%以上),显示出一种软体动物优势较为明显[31]。夏季的ABC曲线显示丰度曲线起点高于生物量曲线,然后相交,且W=-0.132,表示夏季潮间带软体动物群落受到中等程度的扰动。

3 讨论
3.1种类数量及生物量变化
基于本次南麂岛调查的实验结果,共鉴定出软体动物种类63种,对比高爱根、彭欣等在20世纪90年代和本世纪初对南麂列岛软体动物的调查结果,差异较大。根据资料显示,1992—1993年南麂列岛岩礁潮间带发现103种软体动物,2003年发现93种,2006—2007年发现84种,此次调查仅发现63种,总体来讲物种数递减且速度较快,此次调查的软体动物的平均丰度(506 ind/m2)、平均生物量(801.54 g/m2)低于1992年调查结果(1 771.42 ind/m2和2 179.22 g/m2)。这可能主要是由于近年来对南麂列岛的开发不当和环境污染、生态破坏等因素造成的。从本世纪初开始,每年有大量游客进入南麂岛旅游,岛上经济发展迅猛,旅游资源开发迅速,为此大量的服务设施开始兴建,比如新建码头、海景宾馆、酒店等,产生大量的建筑、生活垃圾,破坏了当地的生态环境,并且有部分渔民为了谋取经济利益,铤而走险盗采经济价值较高的野生软体动物(如牡蛎),使软体动物的生存危机进一步加大。历次调查结果的差异也有可能与不同断面范围采样有关(此次调查点仅位于南麂岛,未包括周边岛屿,且样方大小和样方数存在差异),故难以反映南麂岛潮间带软体动物的全貌,但能客观反映南麂岛不同时段潮间带采样断面的软体动物分布状况。
四个季节之间的显著差异还体现在软体动物的平均丰度和平均生物量上,春季是各类软体动物的主要繁殖期,所以春季的丰度和生物量都较大,夏季天气较为炎热,许多软体动物都隐藏在岩石缝隙中,且夏季人为的采捕活动会增多,上岛人数逐年增多,夏季每天最多可达上千人次,游客品尝海鲜大多是经济贝类[21],所以夏季的量都比较小,但由于处在大多数软体动物的繁殖发育期,个体相较其他季节大。秋季大量新的补充个体出现,使丰度和生物量再次增加,但平均个体较小。冬季大多数广温广布种和温带种从低潮区向浅水区下移,使丰度和生物量再次减少,同时,江浙沿岸流的加强以及台湾暖流的变弱,使得新个体成长较慢,直到冬末春初,个体才慢慢长大。
3.2多样性分析
Shannon-Wiener多样性指数(H′)各季节平均差异较大,且冬季和春季的多样性指数明显高于夏、秋季节,这可能与当地的渔民活动和旅游业发展有关,南麂岛每年4—11月是旅游旺季,游客与当地居民的相关活动势必会影响到软体动物的多样性;夏季的高温,使得许多温带种和广温广布种受到影响,迁移到潮下区或者消失,也对其多样性产生一定的影响。P7的多样性指数在四个季节中都是各断面数值最高或次高,这可能是由于P7位于南麂岛的南端,在大沙岙的岙口附近,且岙口朝南,有利于台湾暖流的进入和冲击,浪高且急,拍打范围大,将更多的海水营养盐类输送到更高更广的潮间带,使得P7软体动物生存的范围扩大,减少因潮间带生物内部争夺生存空间的机会,且岙口的存在有利于规避冬季的江浙沿岸流,利于此处多样化的软体动物的繁殖发育。
3.3群落结构特征分析
Bray-Curtis相似性聚类分析和MDS分析结果表明,四个季节中,P2为单独一个群落,其余各点为一个群落。软体动物群落的空间差异主要是不同断面的环境存在差异,导致其生境不同,不同生境导致栖息的种类及优势种的软体动物也不同,形成了不同群落在空间排序上的群聚[32]。这可能是因为P2断面设在大山脚砾石滩,生态类型为砾石,冲浪作用使砾石滩的石面变得更加平滑,蓄水性差,多数软体动物难以附着生长,甚至优势种条纹隔贻贝在这个断面都极少分布,而其余断面基本以适合软体动物繁殖的岩礁潮间带为主,致使P2与其他断面的软体动物群落差异很大。
经SIMPER分析,软体动物的组内相似性冬季最高,秋季最低,冬季南麂岛海域受到江浙沿岸流和台湾暖流的双重影响,但是台湾暖流较弱,所以冬季南麂岛海域总体来讲低温低盐,温带种和广温广布种发育较好,而亚热带种和热带种较为稀少,群落整体较为稳定。秋季,台湾暖流较为强烈,且过了炎热的夏季,气温变冷,许多热带种和亚热带种开始了秋季的繁殖,许多新物种开始显现,此时,在夏季藏于潮下带的物种开始向潮间带迁移,整个群落在相互补充,软体动物在地形开阔、宽敞、有较大岩礁面的断面繁殖发育较快(如P7、P6),但是在较为封闭或者生境较差的断面如(P2和P3)繁殖发育较为迟缓,导致秋季各断面差异较大的情形。在组间相异性分析中得出,差异最大的是冬季-春季,最小的是夏季-秋季,对其差异贡献率最大的是条纹隔贻贝,由于冬季受江浙沿岸流影响,水温气温较低,而条纹隔贻贝属于亚热带种,小个体的条纹隔贻贝会自然死亡,春、夏季是条纹隔贻贝生殖腺成熟期[33],且春季气温回暖,海水营养盐丰富,小个体条纹隔贻贝会大量涌现。夏、秋两季整体的水文条件变化不大,都是受到台湾暖流的影响,且当地居民和游客的采捕活动也集中在这两季,所以这两季差异和变化不大。
3.4ABC曲线特征分析
根据调查结果显示,春季、秋季、冬季这三个季节的软体动物群落尚未受到较大的干扰,且显示出条纹隔贻贝在群落中的优势较为明显。从全年来看,软体动物群落仍然受到一定程度的干扰,尤其是夏季软体动物群落受到了中等程度的扰动,春季和秋季一些机会物种的数量开始增加,这也预示着潮间带生境出现了污染或干扰的趋势[25]。夏季群落受到波动可能是由于南麂岛渔民和游客大量采捕当地软体动物造成的,且气温较高,许多软体动物藏于岩缝中,或者迁移到潮下带,从而也对于全年的群落结构产生了重大的影响。近年来南麂岛基础设施的建设、旅游产业的大力发展、岛屿周边海洋经济生物养殖业的兴盛、海水污染等,或许这些才是影响南麂岛潮间带软体动物群落结构变动最主要的因素。
[1]Cushing D H.The flow of energy in marine ecosystemswith special reference to the continental shelf[C]//Postma J.Ecosystems of the World 27,Continental Shelves.Amsterdam:Elsevier,1988:203-230.
[2]Fernández C.Current status and multidecadal biogeographical changes in rocky intertidal algal assemblages:The northern Spanish coast[J].Estuarine,Coastal and Shelf Science,2016,171:35-40.
[3]Wheat E,Ruesink J L.Commercially-cultured oysters(Crassostreagigas)exert top-down control on intertidal pelagic resources in Willapa Bay,Washington,USA[J].Journal of sea research,2013,81:33-39.
[4]Misra S,Kundu R.Seasonal variations in population dynamics of key intertidal molluscss at two contrasting locations[J].Aquatic Ecology,2005,39(3):315-324.
[5]邹莉,张龙,张宾,等.浙江省沿岸岛礁区贝类种类组成与分布特征[J].水产学报,2010,34(11):1769-1775.
[6]纪莹璐,赵宁,王振钟,等.乳山湾潮间带春季大型底栖动物群落结构[J].应用生态学报,2015,26(2):609-615.
[7]柯盛,申玉春,谢恩义,等.雷州半岛流沙湾潮间带底栖贝类多样性[J].生物多样性,2013,21(5):547-553.
[8]冷宇,刘一霆,刘霜,等.黄河三角洲南部潮间带大型底栖动物群落结构及多样性[J].生态学杂志,2013,32(11):3054-3062.
[9]蔡林婷,王一农,李祥付,等.舟山东极岛潮间带贝类生态学初步研究[J].海洋科学,2013,(8):47-54.
[10]李荣冠,江锦祥,鲁琳,等.鼓浪屿软相潮间带生态初步研究[J].海洋学报:中文版,1996,(2):123-129.
[11]高爱根,陈国通,杨俊毅.南麂列岛海洋自然保护区潮间带软体动物生态研究[J].东海海洋,1994,12(2):44-61.
[12]高爱根,曾江宁,陈全震,等.南麂列岛海洋自然保护区潮间带贝类资源时空分布[J].海洋学报,2007,29(2):101-111.
[13]高爱根,曾江宁,徐晓群,等.南麂列岛大沙岙沙滩贝类的时空分布[J].海洋学研究,2008,26(2):13-19.
[14]高爱根,董永庭,王慧珍,等.南麂列岛邻近海域贝类生态分布的初步研究[J].东海海洋,1998,16(2):50-54.
[15]尤仲杰,孙建璋,王一农.南麂列岛的贝类[C]//尤仲杰.南麂列岛国家级海洋自然保护区论文选(一).北京:海洋出版社,1992:34-54.
[16]尤仲杰,王一农,南麂列岛岩相潮间带软体动物群落生态研究[C]//中国贝类学会.贝类学论文集第四期.青岛:海洋大学出版社,1993.
[17]尤仲杰,王一农.南麂列岛海产双壳类的补充报道[J].浙江水产学院学报,1989,8(1):17-28.
[18]汤雁滨,廖一波,寿鹿,等.珊瑚藻类对南麂列岛潮间带底栖生物群落多样性的影响[J].生物多样性,2014,22(5):640-648.
[19]汤雁滨,廖一波,寿鹿,等.南麂列岛潮间带大型底栖动物群落优势种生态位[J].生态学报,2016,36(2):489-498.
[20]彭欣,谢起浪,陈少波,等.南麂列岛潮间带底栖生物时空分布及其对人类活动的响应[J].海洋与湖沼,2009,40(5):584-589.
[21]高爱根,陈全震,曾江宁,等.人类活动对南麂列岛海洋自然保护区的影响分析[J].海洋开发与管理,2006,23(5):112-115.
[22]中华人民共和国国家质量监督检验检疫总局,中国国家标准化管理委员会.GB/T12763.6-2007海洋调查规范 第六部分:海洋生物调查[S].北京:中国标准出版社,2007.
[23]陈亚瞿,徐兆礼,王云龙,等.长江口河口锋区浮游动物生态研究Ⅰ生物量及优势种的平面分布[J].中国水产科学,1995,(1):49-58.
[24]Wilhm J L.Use of biomass units in Shannon’s formula[J].Ecology,1968,49(1):153-156.
[25]刘淑德,线薇微,刘栋.春季长江口及其邻近海域鱼类浮游生物群落特征[J].应用生态学报,2008,19(10):2284-2292.
[26]孙莎莎,唐文乔,郭弘艺,等.靖江沿岸秋季鱼类群聚的组成特点及其丰度生物量变化[J].生物多样性,2013,21(6):688-698.
[27]施时迪.大陈岛潮间带软体动物区系研究[J].台州学院学报,2000,3:62-64.
[28]安传光,赵云龙,林凌,等,长江口九段沙潮间带大型底栖动物季节分布特征的初步研究[J].水产学报,2007,31(B09):52-58.
[29]Weis J S,Weis P.Is the invasion of the common reed,Phragmitesaustralis,into tidal marshes of the eastern US an ecological disaster[J].Marine Pollution Bulletin,2003,46(7):816-820.
[30]T Thorin S,Radureau A,Feunteun E,et al.Preliminary results on a high east-west gradient in the macrozoobenthic community structure of the macrotidal Mont Saint-Michel bay[J].Continental Shelf Research,2001,21(18):2167-2183.
[31]田胜艳,于子山,刘晓收,等.丰度/生物量比较曲线法监测大型底栖动物群落受污染扰动的研究[J].海洋通报,2006,25(1):92-96.
[32]胡成业,徐衡,邹莉,等.浙江沿岸岛礁区潮间带软体动物群落结构研究[J].水产学报,2015,39(8):1144-1154.
[33]张永普,郑洁,王一农.浙南岛屿岩相潮间带贻贝类的生态特点[J].海洋湖沼通报,2000,3:24-28.
Community structure and diversity of molluscs in the intertidal zone of Nanji Island
BI Siyao1,CAI Houcai2,CHEN Wandong2,LIN Li2,YU Cungen1*,XIA Lujun1,Cheikh Sarr1,XIE Xu1
(1.Marine Fishery College of Zhejiang Ocean University,Zhoushan 316024,China;2.Nanji Islands National Marine Nature Reserve Administration,Pingyang 325401,China)
To reveal the community structure and diversity of intertidal zone molluscs in the Nanji Islands,the quantitative and qualitative investigation of Nanji Island intertidal seven sections were studied for four times in November 2013 and February,May,August 2014.Traditional diversity analysis,community and non-metric multi-dimensional cluster,Abundance Biomass Comparison curve analysis method of combining measures were used to research community structure and diversity of molluscs.The results showed that:(1)63 molluscss species were identified belonging to 3 classes,13 orders,34 families and 52 genuses.In spring,summer,autumn and winter,48 kinds,48 kinds,45 kinds,41 kinds of molluscs were identified,respectively.Dominance of species in different seasons is different,and there are four kinds in spring,four in summer,five in autumn and three in winter.Septifervirgatusis the largest dominance of the species researched.(2)Seasonal differences in changes in community structure consisting of molluscs is notable,and spring,summer,autumn,winter group within similarity were 30.77%,24.66%,19.58%,39.01%,respectively.In the spring - summer and summer - autumn,autumn-winter,winter - spring,the average dissimilarity were 39.82%,39.00%,47.64%,48.02%,respectively.(3)Molluscs Shannon-Wiener diversity index(H′)in the fourth quarter average was winter(1.73)>spring(1.69)>summer(1.58)>autumn(1.40).(4)With cluster analysis and non-metric multidimensional scaling(NMDS)analysis,the intertidal zone molluscs community can be divided into two groups,mainly due to the different habitats.(5)Abundance Biomass Comparison curve showed that spring,autumn,winter community structure did not suffer significant interference,the summer disturbed moderate intensity.
molluscs;intertidal zone;diversity;community structure;Nanji Island
2016-03-17
国家自然科学基金项目(31270527);南麂列岛国家海洋自然保护区管理局委托项目(NJKJ-2013-003).
毕耜瑶(1989- ),男,山东滨州人,硕士研究生,研究方向:渔业资源. E-mail:bisiyao@163.com
俞存根(1960- ),男,浙江永康人,教授,主要从事渔业资源与渔业生态学研究. E-mail:cgyu@zjou.edu.cn
S932.6
A
1006-5601(2016)02-0102-10
毕耜瑶,蔡厚才,陈万东,等.南麂岛潮间带软体动物多样性与群落结构[J].渔业研究,2016,38(2):102-111.
猜你喜欢
海洋信息技术与应用(2020年4期)2021-01-18
海洋通报(2020年3期)2020-10-20
烹调知识(2020年10期)2020-10-20
环球时报(2019-07-09)2019-07-09
石油化工建设(2018年1期)2018-07-10
东南文化(2016年4期)2016-08-22
浙江大学学报(农业与生命科学版)(2015年4期)2015-12-15
应用海洋学学报(2015年1期)2015-11-22
中学生天地(A版)(2015年7期)2015-08-06
读者(乡土人文版)(2013年10期)2013-04-12
