
坛紫菜种质品系表型性状的变异规律分析
2016-08-24 06:39仵燕青谢潮添谢小雪陈昌生纪德华
渔业研究 2016年3期
仵燕青,谢潮添,谢小雪,陈昌生,纪德华,许 凯,徐 燕
(集美大学水产学院,福建 厦门 361021)
坛紫菜种质品系表型性状的变异规律分析
仵燕青,谢潮添,谢小雪,陈昌生,纪德华,许凯,徐燕*
(集美大学水产学院,福建 厦门 361021)
坛紫菜(Pyropiahaitanensis)的表型性状是遗传育种的主要考察指标。研究以福建省坛紫菜种质资源库保存的73份种质品系为实验材料,对藻体的5个形态特征和14个数量性状数据进行了统计分析,以探讨坛紫菜表型性状的遗传变异规律。结果表明,坛紫菜藻体的形状、颜色、锯齿大小和扭曲程度4个形态特征与藻体长度、宽度、重量和厚度4个数量性状极显著相关(P<0.01),叶片基部形状与藻体长度、宽度、鲜重和厚度显著相关(P<0.05);比较不同性状均值发现,线形藻体的平均长度增长速度最快,基部为心脏形的藻体平均重量增长速度最快,无锯齿的藻体厚度最薄。巢式方差分析结果显示,坛紫菜品系间的遗传变异(72.79%)远大于品系内(21.46%),说明品系间的变异是坛紫菜表型变异的主要来源。坛紫菜藻体各表型性状的变异系数为7.14%~35.99%,其中藻体厚度变异系数最低,而单株藻体重量的变异系数最高,说明藻体厚度的选择潜力相对较小,而重量的选择潜力则较大。这些数据为后续坛紫菜的遗传育种提供了基本资料。
坛紫菜;表型性状;方差分析;表型变异系数
紫菜(Pyropia/Porphyra)广泛分布于寒带至亚热带潮间带的岩石上,是世界上可人工栽培产值最高的大型海藻[1],目前世界上已经记录的134个种中,我国分布有22个种[2]。坛紫菜(Pyropiahaitanensis)是我国特有的暖温带性种类,在福建、浙江南部和广东北部沿海被广泛栽培。由于坛紫菜在群体栽培的情况下很容易实现同质化,在栽培生产中就会逐渐呈现出品质和抗逆能力的退化,影响坛紫菜栽培的效益。为解决坛紫菜种质资源存在的问题,研究坛紫菜的遗传多样性就显得非常必要。
遗传多样性是生物多样性的一个重要反映,它代表着一个居群对环境的适应能力[3-4],一个物种遗传多样性越高,对环境变化的适应能力就越强[5],它决定了物种的稳定性和进化潜力[6]。遗传多样性可以表现在多个层次上,如分子、细胞、个体等,表型性状的差异通常是个体遗传多样性最直观的体现[5-6]。表型性状在植物中通常是指植株形态和其它可观察或测量的生长性状[7]。藻类方面,我国藻类学家很早就注意到了海带表型数量性状的变化。曾呈奎等[8]在海带幼苗的栽培研究中发现海带的均重随着其藻体长度和宽度的增加而增加。20世纪60年代,方宗熙等[9-13]在海带遗传育种的研究中发现海带亲代藻体长度和子代藻体长度的正相关关系,同时也证明了海带藻体厚度是一个重要的数量性状。张景镛等[14]通过将海带连续自交和定向选育,测得其子代平均藻体厚度都比母本的藻体厚。在紫菜方面,童冠文等[15]在条斑紫菜经济性状的研究中发现,条斑紫菜单孢子子代群体可以很好地保持亲本的优良性状,可从中选育到具特定藻体形状的品系。陈昌生等[16]开展了坛紫菜品系间杂交研究,并对杂交子代和亲本的经济性状进行了比较分析。徐燕等[17-18]对坛紫菜6个表型性状(长度、宽度、厚度、重量、长度日增长率和重量日增长率)的遗传特征进行了分析,并结合坛紫菜高密度遗传连锁图谱实现了这6个性状的QTL定位。
为进一步系统地研究坛紫菜种质品系表型的变异规律,本研究以福建省坛紫菜种质资源库保存的73个坛紫菜种质品系为实验材料,分析了19个表型性状的特征,并基于各表型性状数据进行了遗传变异规律分析,为坛紫菜的种质资源保护、核心种质库构建及良种选育提供基础资料。
1 材料与方法
1.1材料及处理
研究所采用的73份坛紫菜种质品系均取自福建省坛紫菜种质资源库(其中40份为诱变或杂交所选育,33份为采自福建沿海各地,并经纯化的野生坛紫菜种质)。
坛紫菜种质丝状体的促熟和促放:取一定量种质丝状体进行40 d左右高温促熟(27~29℃),当丝状体形成壳孢子囊枝时,采用降温和充气刺激使成熟的种质丝状体放散壳孢子。
叶状体的培养:壳孢子充气培养至小苗后,每个品系随机挑选生长正常,形状完整,长度4.0~4.2 cm的叶状体15株,测量后单株置于500 mL锥形瓶中充气培养,期间每隔3天更换一次新鲜培养液,每隔5天测量一次藻体长度、宽度和鲜重,培养期间每天观察并比较不同品系藻体的生长、发育及颜色等的变化情况。培养条件为:温度(21±1)℃,光强2 000~3 000 lx,光周期L∶D=12 h∶12 h,培养时间15 d,恒温培养室中充气培养。
1.2实验方法
1.2.1表型性状记录及计算
各种质品系的表型性状采用5个形态特征和14个数量性状进行评价。形态特征包括藻体形状、藻体颜色、基部形状、锯齿大小、扭曲程度。数量性状包括藻体长、藻体宽、藻体长宽比、藻体厚度(基部、中部、尖端)、藻体胶质膜厚度(基部、中部、尖端)、重量、长度日生长速率、重量日生长速率、长度绝对生长速率、重量绝对生长速率。计算公式如下:
叶片厚度:随机取12株(25±5)cm的藻体,分别测量藻体1/4、1/2、3/4处的厚度及胶质膜厚度,取平均值。
叶片宽度:随机取12株(25±5)cm的藻体,分别测量藻体1/4、1/2、3/4处的宽度,取平均值。
式中Ln为第n天的藻体长度(cm);L0为初始藻体长度(cm);Wn为第n天的藻体重量(g);W0为初始藻体重量(g)。
1.2.2表型性状的统计分析
对各种质品系的表型特征中的形态特征进行统计,计算各性状的频率,并进行方差分析,利用巢式设计模型对藻体长、藻体宽、藻体厚度、重量、长度日生长速率、重量日生长速率、长度绝对生长速率、重量绝对生长速率8个数量性状进行方差分析。
1)各性状方差分析的线性模型参照李斌[19]等的方法,计算公式为:
Yijk=μ+Si+T(i)j+e(ij)k
式中,Yijk为第i个群体(品系)第j个藻体第k个观测值;μ为总均值;Si为群体效应(固定);T(i)j为群体内品系效应(随机);e(ij)k为实验误差。
2)表型分化系数计算方法参照葛颂等[20]方法,公式为:
式中,δ2t/s为群体间方差分量;δ2s为群体内方差分量;Vst为表型分化系数,表示群体间变异占遗传总变异的百分比。δ2t/s的计算公式为:
式中,MSt为群体间均方;MSs为群体内均方;j为群体内样本数;k为观测重复数[18]。
3)采用变异特征系数(Cv)表示性状的离散程度,用相对极差R'i表示性状的极差异程度,Cv的计算公式为:
Cv=S/X
式中,S=标准差;X=平均值。
相对极差R'i的计算公式为:
R'i=Ri/Ro×100%
式中,Ri表示局群内的极差;Ro表示总极差。
2 结果
2.1藻体形态特征的变异
坛紫菜73份种质品系叶状体的形态特征记录结果如表1所示。坛紫菜形态特征比较简单,从表1中可以看出:坛紫菜的藻体形状以披针形为主,其次为线性披针,分别占63.38%和32.87%;基部形状以圆形为主,其次为楔形,所占比例分别为68.49%和23.28%;藻体颜色以基部绿色少于1/4的野生色为主,所占比例为32.88%;锯齿程度主要以1~3 mm为主,所占比例为41.89%;扭曲程度以小于藻体1/4为主,所占比例为68.49%。
根据单因素方差分析结果可知,藻体形状、颜色、锯齿和扭曲程度与藻体的长度、宽度、鲜重和厚度极其显著相关(P<0.01)。从藻体形状上看,线形叶片培养15 d后的平均长度最长(61.33 cm),宽线形叶片培养15 d后的平均鲜重最重(0.150 2 g),且平均厚度最薄(26.67 μm);9种颜色藻体中,棕红色藻体平均生长速度最快,培养15 d后长度35.02 cm、宽度0.93 cm、鲜重0.158 6 g,深绿色藻体平均厚度最薄为25.73 μm;具波浪状锯齿的藻体比其他类型藻体生长速度更快(培养15 d后,长度为27.71 cm),无锯齿的藻体相对较重和薄;平整无扭曲的藻体长度和重量增长明显,培养15 d后平均长度和重量分别为30.24 cm 和0.127 1 g。叶片基部形状与藻体长度、宽度、鲜重和厚度显著相关(P<0.05),基部为心脏形的藻体增重明显(培养15 d后鲜重为0.157 6 g),且平均厚度最薄(22.50 μm)。

表1 坛紫菜叶状体不同表型性状的分布频率及均值

续表1
2.2各种质品系表型性状品系间方差分析结果
对各品系表型性状进行单因素方差分析,结果显示品系间各性状呈极显著差异(P<0.01)(表2)。进一步根据巢式方差分析结果,得出坛紫菜藻体长、宽、长宽比、重量等表型性状的方差分量、方差分量百分比,以及表型分化系数,结果如表3所示。从表中可以看出,各性状品系间的方差分量均大于品系内的方差分量,品系间方差分量均值为72.79%,品系内方差分量均值为21.46%,说明品系间变异是坛紫菜表型变异的主要部分。各性状的表型分化系数为64.24%~99.9%,表型分化系数最小的性状是长宽比,最大的性状是基部的胶质膜厚度,平均表型分化系数为92.96%。
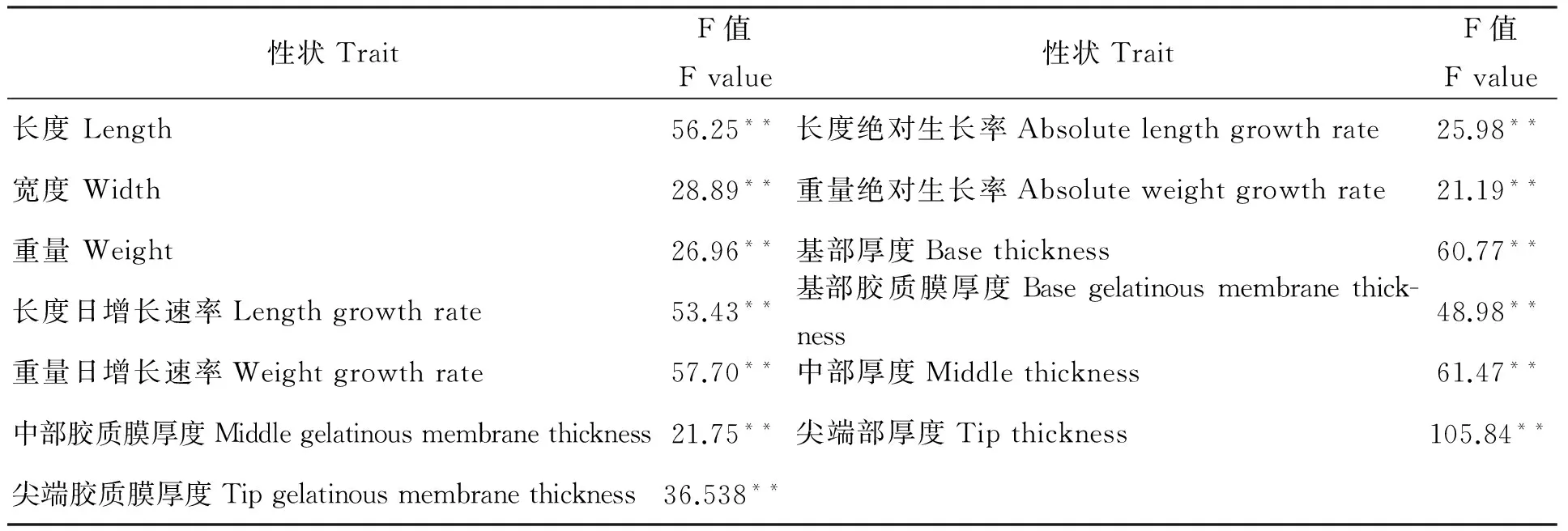
表2 坛紫菜品系间表型性状方差分析

表3 坛紫菜品系间与品系内表型性状方差分析
2.3各表型性状的表型变异系数、极差、变异幅度和分化系数
各种质品系各表型性状的变异系数、极差、变异幅度、表型分化系数的统计结果如表4所示。从表中可以看出,14个数量性状的平均变异系数存在一定差异,其中重量最大,尖端厚度最小。变异系数表示性状值的离散性特征,变异系数大则性状离散程度大。本实验结果说明坛紫菜藻体的尖端厚度性状比其他性状更为稳定,而重量性状则变化相对较大,变异范围最宽,变异系数最大,遗传多样性最为丰富,变异系数为35.99%,变异幅度为0.002~0.443 9 g。表型性状的相对极差以中部厚度、宽度及重量性状较大,长度绝对生长速率、重量绝对生长速率较小。

表4 坛紫菜品系间变异系数、相对极差、变异幅度的统计结果
CV:变异系数;R’:相对极差。
Notes:CV indicated variation;R’ indicated relative range.
3 讨论
表型性状是研究遗传多样性在形态学水平的基本手段,可以反映生物居群遗传因素及其生活环境互作的复杂性及其适应环境的能力[21]。紫菜的表型性状比较简单[22],但在紫菜遗传育种中,表型性状是其中主要的选育指标,如陈昌生等[23]进行坛紫菜品系间杂交选育时就着重考察了藻体的长度、宽度、颜色、鲜重、平均日增长率、平均日增重率等指标;严兴洪等[24]在筛选坛紫菜优良品系时,也把藻体的颜色、体型、生长速度等性状作为筛选的指标;纪德华[25]等在野生坛紫菜进行优良品系选育时选取了颜色、形状、长宽、厚度、锯齿等作为考察指标;梁艳等[26]在研究坛紫菜新品系的优良性状时选取了藻体的日增长量、日增重率、厚度、色素含量、蛋白含量、氨基酸含量等作为考察指标。相比陆生经济作物以及水产动物而言,大型经济海藻的遗传育种研究起步较晚,经济性状的判断标准还不是很完善,目前仅海带的经济性状标准较为完善,但学者们对紫菜的表型性状研究正在不断的完善中[27]。总体来看,目前对紫菜表型性状的研究主要针对的形态特征包括:藻体形状、颜色、锯齿及基部形状等;数量性状包括:藻体长度、宽度、重量、厚度以及重量日增长速率、长度日增长速率等。如徐燕[17]等对坛紫菜DH群体的数量性状进行统计分析时选取了长度、宽度、厚度、鲜重、长度增长率和重量增长率6个表型性状进行统计分析。但已有的研究都是从单一种质或是单一群体入手,并未涉及大量来源于不同产地的坛紫菜种质的研究,本研究通过对坛紫菜73份种质品系14个数量性状和5个形态特征进行统计分析,试图从中找到坛紫菜表型性状的变异规律,以指导坛紫菜的遗传育种工作。
紫菜藻体的长度、宽度、厚度、重量、生长速度等表型性状是决定产量和质量的主要性状,这些性状受多基因控制,易受环境条件影响,这就使得在遗传育种中很难直接通过表现型选择来达到基因型选择的目的。本研究结合前人的经验,通过比较不同表型性状的均值,并进行方差分析和性状相关性分析,以期揭示各数量性状和形态特征的相关性,从而可以从藻体的形态特征预测各性状育种潜力的目的,以达到缩短育种周期、提高育种效率的目的。如需要选择长度生长速度快的品系,可以在线形和宽线型藻体中选择;需要选育重量增长速度快的品系,可以从宽线型,基部形状为心脏形,颜色为棕红色的藻体中选择;选育薄叶型品系,可以从基部为心脏形,无锯齿的藻体中选择。巢式方差分析结果显示,坛紫菜各表型性状品系间的方差分量均远大于品系内的方差分量,说明品系间的变异是坛紫菜表型变异的主要来源。变异系数是衡量某种性状变异程度大小的一个重要指标,大的变异系数意味着从群体中选择获得这一性状优良个体的概率也相对较高。本研究发现坛紫菜表型性状的变异系数为7.14%~35.99%,其中藻体厚度的变异系数最低,基部、中部和尖端的变异系数均小于9%,而单株藻体重量的变异系数则最高为35.45%,说明藻体厚度的选择潜力相对较小,而重量的选择潜力则较大,这与徐燕等[17]和王清印[12]的研究结果基本一致。因此,在今后坛紫菜的遗传育种中,可以参照表型遗传变异的分析数据及数量性状与形态特征的相关性进行优良品系的选育。
[1]李晓川.我国紫菜产业发展、标准状况及安全性评价[J].中国渔业质量与标准,2011,1(3):9-12.
[2]曾呈奎,相建海.海洋生物技术[M].济南:山东科学技术出版社,1998:231-232.
[3]王清印.海水养殖生物的细胞工程育种[M].北京:海洋出版社,2007:314.
[4]郗荣庭,张毅萍.中国核桃[M].北京:中国林业出版社,1992:121-122.
[5]卢欣石,何琪.种群遗传变异及基因多样度分析[J].草业学报,1999,(3):76-82.
[6]王洪新,胡志昂.植物的繁育系统、遗传结构和遗传多样性保护[J].生物多样性,1996,(2):32-36.
[7]顾万春.统计遗传学[M].北京:科学出版社,2004:1-422.
[8]曾呈奎,孙国玉,吴超元.海带的幼苗低温度夏栽培实验报告[J].植物学报,1955,4:255-264.
[9]方宗熙,蒋本禹,李家俊.海带遗传和育种的研究[J].高等学校自然科学学报(生物学版),1965,3:92-40.
[10]方宗熙,蒋本禹,李家俊.海带叶片长度遗传的进一步研究[J].海洋与湖沼,1965,7(1):59-66.
[11]方宗熙,蒋本禹.海带叶片长度的遗传[J].海洋与湖沼,1963,(2):172-182.
[12]王清印.海带几个经济性状遗传力和遗传相关的研究[J].山东海洋学院学报,1984,14(3):65-76.
[13]Miura W,Fujio Y,Suto S.Genetic differentiation between the wild and cultured populations ofPorphyrayezoensis(algae)[J].Tohoku Journal of Agricultural Research,1980,30(3):114-125.
[14]张景镛,方宗熙.海带叶片厚度遗传的初步研究[J].遗传学报,1980,(3):257-262.
[15]童冠文.条斑紫菜经济性状的遗传分析[D].青岛:中国海洋大学,2010.
[16]陈昌生,徐燕,纪德华,等.坛紫菜品系间杂交藻体选育及经济性状的初步研究[J].2007,31(1):97-104.
[17]Xu Y,Xie C T,Chen C S,et al.Genetic analyses of six quantitative traits of a doubled haploid population ofPorphyrahaitanensisChang et Zheng(Bangiales,Rhodophyta)[J].Journal of Applied Phycology,2012,24(1):89-96.
[18]Xu Y,Huang L,Ji DH,et al.Construction of a dense genetic linkage map and mapping quantitative trait loci for economic traits of a doubled haploid population ofPyropiahaitanensis(Bangiales,Rhodophyta)[J].BMC plant biology,2015,15(1):228.
[19]李斌,顾万春,卢宝明.白皮松天然群体种实性状表型多样性研究[J].生物多样性,2002,10(2):181-188.
[20]葛颂,王明庥,陈岳武.用同工酶研究马尾松群体的遗传结构[J].林业科学,1988,24(4):399-409.
[21]Bacilieri R,Ducousso A,Kremer A.Genetic,morphological,ecological and phenological differentiation betweenQuercuspetraea(Matt.)Liebl.andQuercusroburL.in a mixed stand of northwest of France[J].Silvae Genetica,1995,44(1):1-9.
[22]Kavale M G,Kazi M A,Sreenadhan N,et al.Morphological,ecological and molecular characterization ofPyropiavietnamensis(Bangiales,Rhodophyta)from the Konkan region,India[J].Phytotaxa,2015,224(1):45-58.
[23]陈昌生,谢潮添,纪德华,等.野生坛紫菜种群遗传多样性的ISSR分析[J].水产学报,2008,32(5):717-724.
[24]严兴洪,陈俊华.坛紫菜优良品系的筛选与特性分析[EB/OL].[2016-04-03].http://cpfd.cnki.com.cn/Article/CPFDTOTAL-ZGHI200708001156.htm.
[25]纪德华,谢潮添,陈昌生,等.野生坛紫菜的选育及经济性状的研究[J].海洋学报,2008,30(1):140-146.
[26]梁艳,徐燕,陈昌生,等.坛紫菜优质新品系(Q-1)主要经济性状的研究[J].渔业科学进展,2009,30(4):108-116.
[27]Jia J,Wang P,Jin D,et al.The application of RAPD markers in diversity detection and veriety identification ofPorphyra[J].Acta Botanica Sinica,1999,42(4):403-407.
Analysis on the variation pattern of phenotypic traits ofPyropia haitanensis germplasms
WU Yanqing,XIE Chaotian,XIE Xiaoxue,CHEN Changsheng,JI Dehua,XU Kai,XU Yan*
(Fisheries College of Jimei University,Fujian 361021,China)
Phenotypic traits are the core selection standards in breeding of new varieties.We chose 73 wildPyropiahaitanensisstrains as experimental material which collected from Fujian coast.We analyzed the genetic diversity ofP.haitanensisthrough phenotype data including 5 basic morphological characteristics and 14 quantitative traits.The results indicated that 4 morphological characteristics including leaf type,color,sawtooth and distortion degree had extremeby significant(P<0.01)relationships with 4 quantitative traits including the length,width,fresh weight and thickness.In addition,the base shape of thallus significantly(P<0.05)correlated with the length,width,fresh weight and thickness.By comparing the average values of different traits,we found that linear thallus showed the highest length growth rate,the thallus with heart shape base exhibited the highest weight growth rate,thallus without sawtooth was the thinnest strain.According to the results of nested analysis on variance among strains,the mean variance components was 72.79% among strains which far larger than mean variance components of 21.46% within strains,which explained the main source ofP.haitanensisphenotypic variation originate between strains.We also found that coefficient of phenotypic variation ofP.haitanensisthallus was ranged from 7.14% to 35.45%,and the variation coefficient of thickness was the lowest,while the mean weight per thallus was the highest.These indicated that thickness only had little potential to improve,but weight was worth to be the target trait.Present study could offer some basic information for futureP.haitanensisbreeding.
Pyropiahaitanensis;phenotype traits;analysis of variance;coefficient of phenotypic variation
2016-05-13
福建省种业创新与产业化工程资助项目(项目编号:2014S1477-10);福建自然科学基金项目(项目编号:2014J05041,2014 J07006);浙江省重中之重一级学科开放基金项目(项目编号:xkzsc09).
仵燕青(1990-),女,硕士生,研究方向为坛紫菜遗传育种.E-mail:1107700709@qq.com
徐燕(1981-),助理研究员,研究方向为坛紫菜遗传育种.E-maill:yanxu@jmu.edu.cn
S968.43
A
1006-5601(2016)03-0173-08
仵燕青,谢潮添,谢小雪,等.坛紫菜种质品系表型性状的变异规律分析[J].渔业研究,2016,38(3):173-180.
猜你喜欢
中国海洋大学学报(自然科学版)(2022年8期)2022-08-15
孩子(2021年1期)2021-02-02
园林科技(2020年2期)2020-01-18
幽默大师(2019年10期)2019-10-17
山东林业科技(2019年2期)2019-06-03
婚姻与家庭·婚姻情感版(2018年4期)2018-04-09
科技创新与应用(2017年20期)2017-07-15
广西林业科学(2016年2期)2016-03-20
小朋友·快乐手工(2015年6期)2015-07-01
湖泊科学(2015年4期)2015-02-17
