
3种冷季型草坪草对丝茅入侵生理响应的差异性分析
2016-08-02 09:06宗人旭刘金平
草原与草坪 2016年3期
马 娇,宗人旭,刘金平,伍 德
(西华师范大学 西南野生动植物资源保护省部共建教育部重点实验室,四川 南充 637009)
3种冷季型草坪草对丝茅入侵生理响应的差异性分析
马娇,宗人旭,刘金平,伍德
(西华师范大学 西南野生动植物资源保护省部共建教育部重点实验室,四川 南充637009)
通过取代试验设计,设置5个丝茅侵入量梯度,模拟入侵高羊茅、草地早熟禾、多年生黑麦草等3种冷季型草坪。70 d后,测定草坪草的抗氧化酶活性、细胞膜伤害程度、光合色素含量及生长发育指标,研究3种草坪草对丝茅入侵生理响应的种间差异及侵入量对草坪草危害差异。结果表明:(1)丝茅的入侵极显著提高了3种草的超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)活性(P<0.01),侵入量间SOD,POD和CAT活性有显著差异,草种间SOD和CAT活性有极显著差异(P<0.01),POD活性差异较小(P>0.05); (2)丝茅入侵伤害了细胞膜透性,极显著提高了3种草的可溶性蛋白(SP)、可溶性糖(SS)、丙二醛(MDA)和游离脯氨酸(Pro)含量(P<0.01),草种间差异为SS>MDA>SP>Pro,侵入量间差异为MDA>SP>SS>Pro;(3)丝茅入侵极显著降低了草坪草的叶绿素a(Chla)、叶绿素b(Chlb)、叶绿素a+b(Chla+b)含量(P<0.01),对Chla/b无显著影响,侵入量间差异为Chla+b>Chla>Chlb>Chla/b,草种间差异为Chla>Chlb>Chla/b>Chla+b;(4)丝茅入侵极显著降低了草坪草的分蘖数与单株生物量(P<0.01);(5)侵入量大于10%时显著提高了SP,SS和Pro含量;>20%显著提高了SOD和POD活性和MDA含量,降低了Chla、Chla+b含量及分蘖数与单株生物量;大于30%显著提高了CAT活性,降低了Chlb含量。3种草坪草对丝茅入侵生理响应差异显著,侵入量越大对草坪草伤害越大,综合防治是解决丝茅危害的根本途径。
冷季型草坪;丝茅;抗氧化酶;细胞膜透性;光合色素
亚热带及过渡性气候带常用高羊茅(Festucaarundinacea)、草地早熟禾(Poapratensisi)、黑麦草(Loliumperenne)混播建植冷季型草坪,以达到四季常绿的绿化效果。由于受最适温度范围、株丛类型、分生再生能力的限制,冷季型草坪抗杂草入侵能力较差。杂草不仅损害草坪的美观性、影响草坪草生长发育、增加养护难度和强度,且杂草是诸多病虫害的中间寄主[1-2]。防除杂草是草坪养护最重要和最基本的工作[3],除草用工占草坪养护总用工的80%,费用占养护费用的60%。国内外对冷季型混播草坪的杂草种类、危害程度及防治技术研究较多[4],对杂草攻击力及竞争力研究较少[5-6],关于杂草入侵对草坪草生理活性、细胞膜透性和光合基础影响的研究极少。分析草坪草应对杂草入侵的生理响应途径,探讨混播草种抵抗杂草胁迫的种间差异,对混播草坪的草种配比、建植技术、杂草防除技术选择及降低养护管理成本具有现实意义。
丝茅(Imperatakoenigii)为禾本科白茅属植物,又称茅针、茅根、白茅根,在热带和亚热带广泛分布[7],是世界上10大恶性杂草之一,目前,尚无有效、持久的防除措施。丝茅根茎发达、种子繁多,是亚热带及过渡性气候带冷季型草坪最常见杂草[8],一旦入侵很难彻底剔除。入侵1~3年后,在草坪中形成局部优势斑块,甚至完全取代草坪草,严重危害草坪种群结构与景观功能。围绕丝茅入侵、扩散及防除进行了大量研究[9-10],结果表明,丝茅入侵降低了草坪草的攻击力和竞争力[11],但有关其危害机理及草坪草应对危害的响应机理报道较少。以3种冷季型草坪草为材料,通过取代试验设计,模拟幼坪期5个丝茅入侵梯度,通过测定草坪草的抗氧化酶活性、细胞膜伤害程度、光合色素含量及生长发育指标,分析3种草坪草对丝茅入侵生理响应的种间差异及侵入量对草坪草危害差异,研究丝茅入侵对草坪草新陈代谢及生长发育的影响,为草坪建植养护、杂草防治提供依据。
1 材料和方法
1.1试验材料及设计
以高羊茅、草地早熟禾、黑麦草及野生丝茅草为材料。采用高34 cm、口径50 cm塑料花盆,河沙与腐殖土1∶1充分混合为基质。
采用复合取代试验设计,于2013年3月,将 3种草坪草分别均匀播种、定苗100株/盆,待2~3真叶时,用丝茅根茎插穗(长2~3 cm,含2~3节)替代部分草坪草幼苗,形成草坪草∶丝茅草为90∶10、80∶20、70∶30、60∶40、50∶50的5个模拟丝茅草入侵梯度,单播草坪草为对照(CK),各处理设3次重复。于25℃的室内培养,期间正常浇水、除杂,于25、50 d修剪(留茬高度7 cm),70 d测定试验指标。
1.2测定项目与方法
1.2.1抗氧化酶活性指标测定超氧化物歧化酶(SOD)活性测定采用氮蓝四唑(NBT)光化还原法[12],过氧化物酶(POD)活性测定采用愈创木酚法[12],过氧化氢酶(CAT)活性测定采用紫外吸收法[13]。
1.2.2细胞膜伤害程度指标测定丙二醛(MDA)采用硫代巴比妥酸(TBA)法[13],脯氨酸(Pro)采用酸性茚三酮法[13],可溶性糖(SS)含量的测定采用蒽酮比色法[12],可溶性蛋白(SP)含量测定采用考马斯亮蓝G-250染色法[13]。
1.2.3光合色素含量测定采用乙醇丙酮混合提取法[14]。
1.2.4生长指标测定每盆随机选草坪草10株,测分蘖数后,105℃下烘至恒重后称干重,计算单株生物量。
1.3数据分析
用SAS 10.0软件对所有数据进行方差分析和析因分析,并用Duncan法对各参数进行显著性检验。
2 结果与分析
2.1抗氧化酶活性指标的响应差异
随丝茅侵入量增加,3种草坪草的SOD,POD和CAT活性均呈增加趋势(图1A,B,C)。析因分析发现,总处理对SOD,POD和CAT活性均有极显著影响(P<0.01)(表1)。不同草种间POD活性差异较小(P>0.05)(图1B),SOD(图1A)和CAT(图1C)活性有极显著差异(P<0.01)。SOD活性在早熟禾和黑麦草中显著高于高羊茅(P<0.05)(图1A),黑麦草CAT活性显著高于早熟禾和高羊茅,早熟禾CAT活性显著最低(图1C)。丝茅侵入量对3种草SOD,POD和CAT活性均有极显著影响(P<0.01)(表1),影响顺序为SOD>CAT>POD。不同侵入量间SOD,POD和CAT活性有显著差异(P<0.05),侵入量>20%后显著提高了草坪草SOD和POD的活性,>30%后显著提高了CAT活性。黑麦草SOD和CAT活性对丝茅入侵最敏感,早熟禾POD活性对高侵入量反应较快。
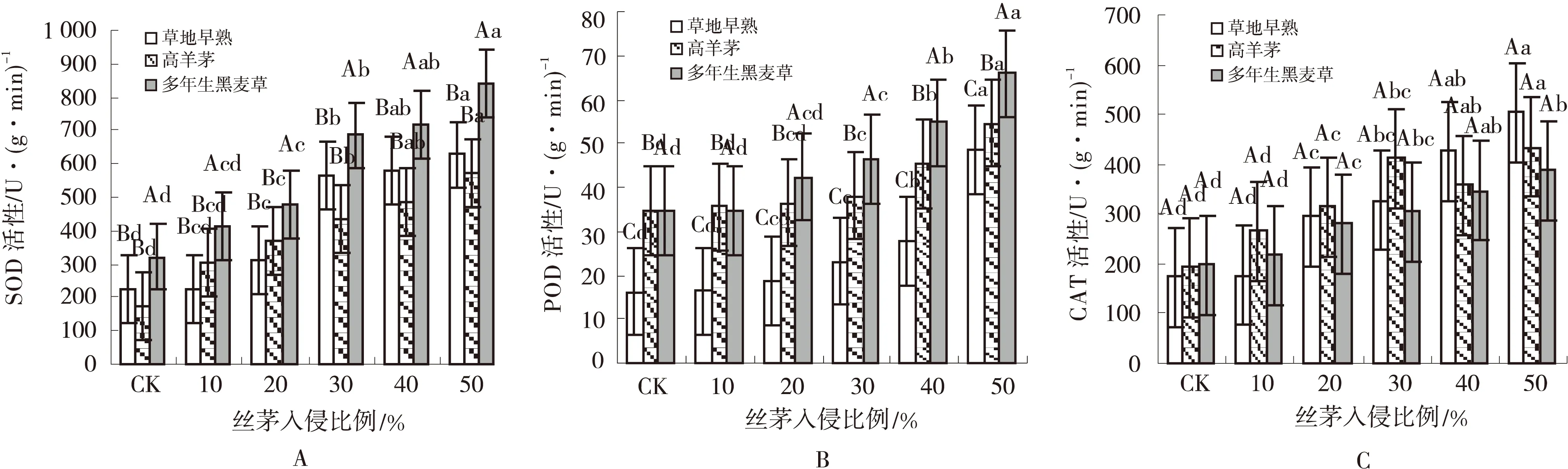
图1 丝茅入侵下的草坪草SOD,POD和CAT活性Fig.1 SOD,POD and CAT activity of turfgrasses invaded by I.koenigii 注:不同大写字母表示草种间差异显著(P<0.05),不同小写字母表示侵入量间差异显著(P<0.05),下图同
2.2细胞膜伤害程度指标的响应差异
总处理间、草种间及侵入量间SP、SS、Pro和MDA含量均有极显著差异(P<0.01)(表1)。草种间差异大小为SS>MDA>SP>Pro,侵入量间差异大小为MDA>SP>SS>Pro。高羊茅和黑麦草的SP、SS、Pro和MDA含量显著大于早熟禾(P<0.05)(图2A,B,C,D)。随丝茅侵入量增加草坪草中SP、SS、Pro和MDA含量均为增加,侵入量>10%时SP、SS、Pro含量均显著高于CK(P<0.05)(图2A,B,C),侵入量大于20%时MDA含量著高于CK(P<0.05)(图2D)。Pro含量在20%、30%、40%和50%侵入量间无显著差异,SS含量在大于30%侵入量后无显著增加,SP和MDA含量则随侵入量增加不断增加。
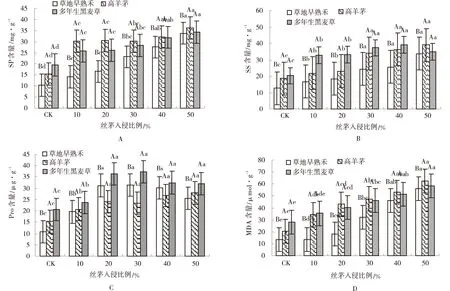
图2 丝茅入侵下的草坪草SP、SS、Pro和MDA含量Fig.2 SP,SS,Pro and MDA content of turfgrasses invaded by I.koenigii
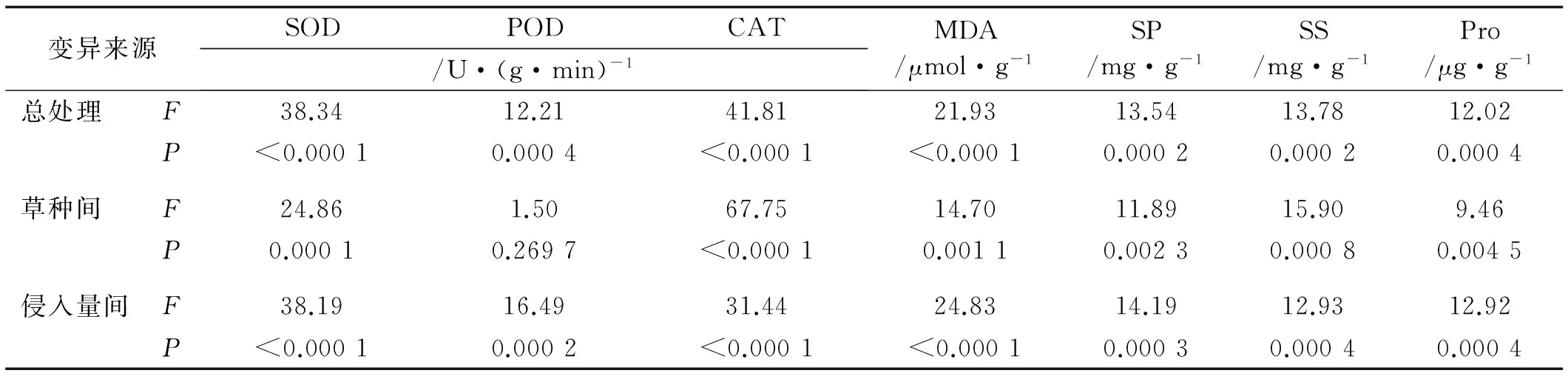
变异来源SODPODCAT/U·(g·min)-1MDA/μmol·g-1SP/mg·g-1SS/mg·g-1Pro/μg·g-1总处理F38.3412.2141.8121.9313.5413.7812.02P<0.00010.0004<0.0001<0.00010.00020.00020.0004草种间F24.861.5067.7514.7011.8915.909.46P0.00010.2697<0.00010.00110.00230.00080.0045侵入量间F38.1916.4931.4424.8314.1912.9312.92P<0.00010.0002<0.0001<0.00010.00030.00040.0004
2.3光合色素含量的响应差异
丝茅入侵极显著降低了草坪草的Chla、Chlb、Chla+b含量(P<0.01)(表2),对Chla/b无显著影响。侵入量间差异大小为Chla+b>Chla>Chlb>Chla/b。随侵入量增加光合色素含量均呈下降趋势,>20%侵入量显著降低了Chla和Chla+b含量,>30%显著降低了Chlb含量。草种间Chla,Chlb和Chla/b存在极显著差异(P<0.01),差异顺序为Chla>Chlb>Chla/b>Chla+b。高羊茅Chla含量显著低于早熟禾和黑麦草(P<0.01)(图3),黑麦草Chlb含量显著低于早熟禾和高羊茅,高羊茅Chla+b含量显著低于早熟禾,黑麦草Chla/b显著高于早熟禾和高羊茅。草种与侵入量总处理对草坪草光合色素含量均有显著影响,影响顺序为Chla+b>Chla>Chlb>Chla/b。
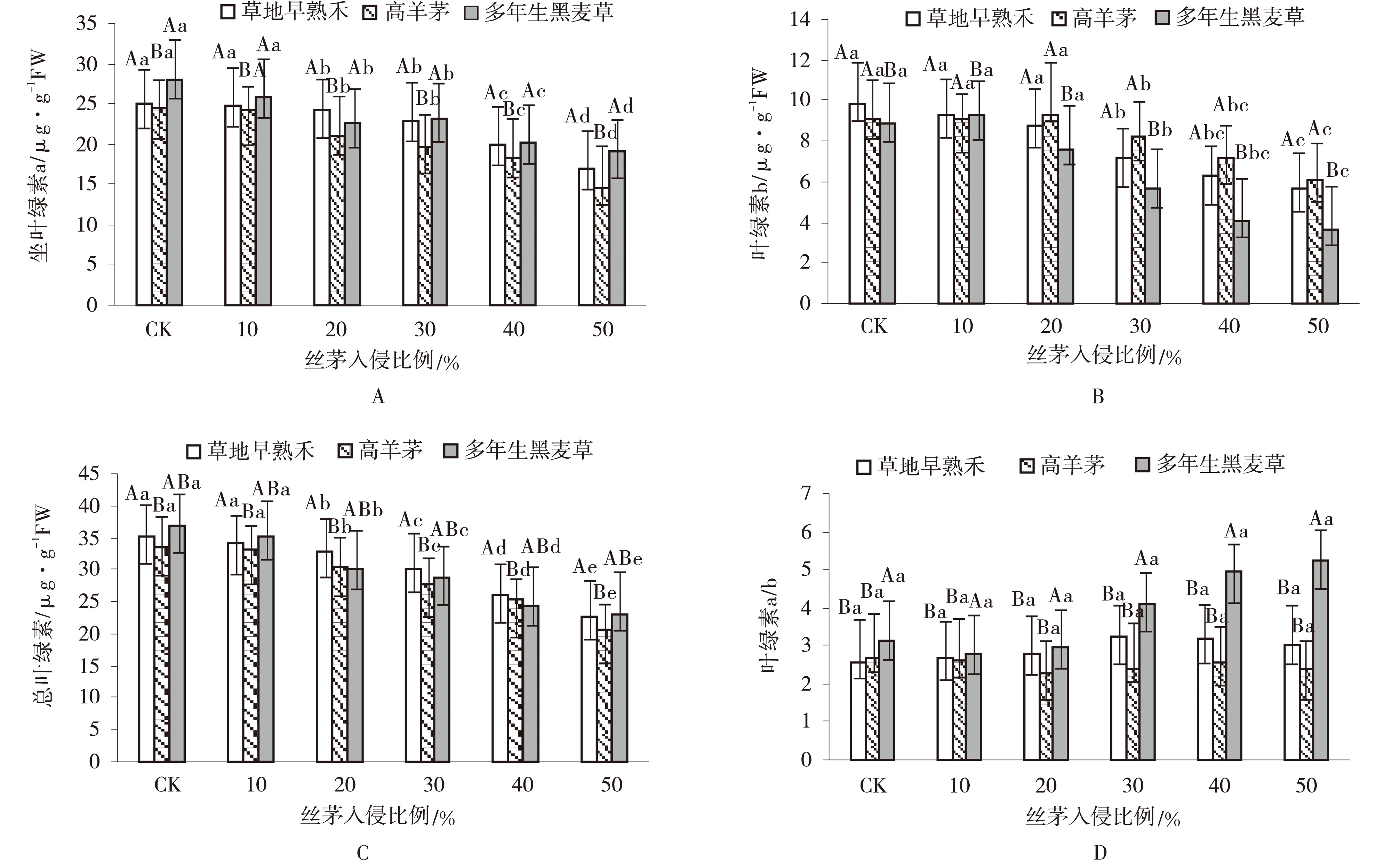
图3 丝茅入侵下的草坪草Chla、Chlb、Chla+b和Chla/b含量Fig.3 Chla,Chlb,Chla+b and Chla/b contents of turfgrasses invaded by I.koenigii
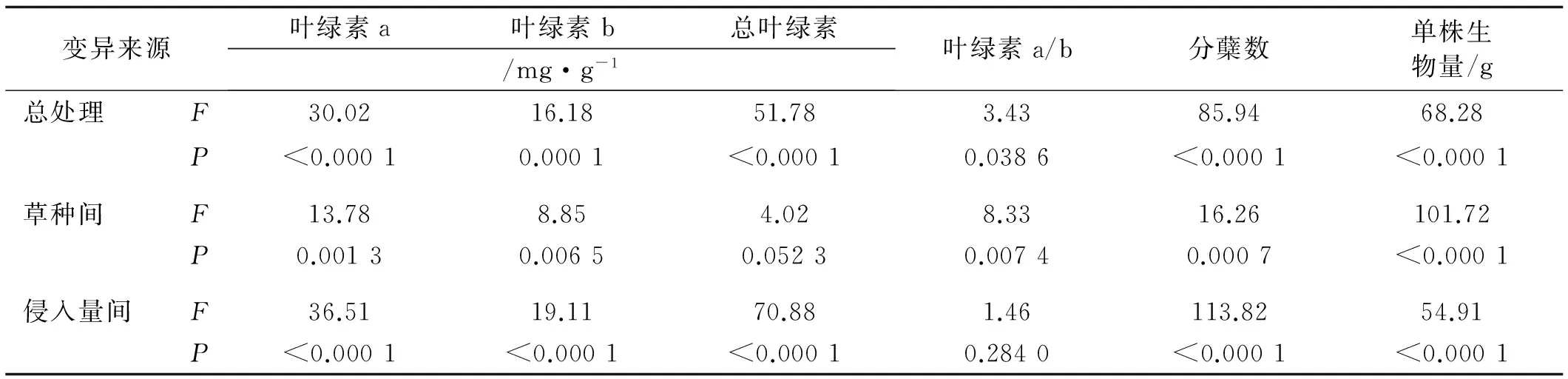
变异来源叶绿素a叶绿素b总叶绿素/mg·g-1叶绿素a/b分蘖数单株生物量/g总处理F30.0216.1851.783.4385.9468.28P<0.00010.0001<0.00010.0386<0.0001<0.0001草种间F13.788.854.028.3316.26101.72P0.00130.00650.05230.00740.0007<0.0001侵入量间F36.5119.1170.881.46113.8254.91P<0.0001<0.0001<0.00010.2840<0.0001<0.0001
2.4生长指标的响应差异
丝茅入侵极显著降低了草坪草的分蘖数与单株生物量(P<0.01)(表2),草种间分蘖数和单株生物量存在极显著差异(P<0.01),侵入量对分蘖数的影响大于生物量。黑麦草的单株生物量显著大于高羊茅与早熟禾(P<0.05)(表3),高羊茅的分蘖数显著高于黑麦草和早熟禾,早熟禾的生物量与分蘖数最低。10%侵入量对草坪草生长影响较小,>20%侵入量均显著降低了草坪草的分蘖数与生物量。侵入量达50%时,早熟禾、高羊茅、黑麦草生物量分别比CK下降了40%、38%和35%,分蘖数分别下降了77%,60%和76%。
3 讨论
植物抗氧化酶系统激活是应对胁迫的基本反应,通过抗氧化酶活性表征植物的受胁迫程度。SOD,POD和CAT协同作用可及时清除植物代谢过程或逆
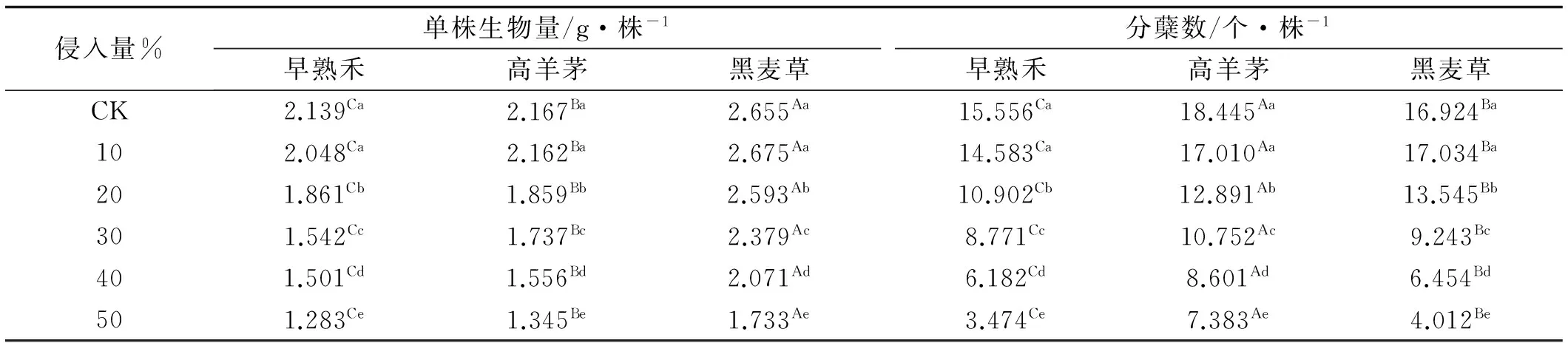
表3 草坪草生长指标对丝茅入侵响应差异
注:同列大写字母不同表示草种间差异显著(P<0.05),小写字母不同表示丝茅侵入量间差异显著(P<0.05)
境胁迫下产生的过多自由基,维持细胞内较低的活性氧水平,保护植物细胞免受损伤。丝茅入侵显著提高了草坪草种SOD,POD和CAT活性,其中SOD活性对丝茅入侵响应最快(侵入量10%),POD次之(侵入量20%),CAT较慢(侵入量30%),结果与植物干旱胁迫下抗氧化酶活性激活顺序一致[15]。抗氧化酶类中,SOD是抵御ROS氧化损伤的第一道防线,可将多余的超氧阴离子结合成H2O2[16],POD和CAT则进一步分解为无害的H2O和O2[17],试验中黑麦草SOD活性显著高于其他草种,说明丝茅入侵对黑麦草影响最大,CAT活性在草种间有极显著差异,表明不同草种清除自由基的方式与清除能力存在差异,POD活性在草种间无明显差异,或许与POD有类似调节植物生长、诱导细胞壁延伸等功能有关。
叶绿素含量受到光照、温度、矿质元素、逆境等外界因素及核基因、质基因等内在因素的共同影响,叶绿素含量的变化与叶绿体生理活性、光合能力及对逆境的适应性和抗性有关[18],叶绿素含量下降是叶片衰老最明显的特征[19]。诸多研究认为光、水分胁迫对叶绿素的合成、含量与比例有重要影响,但关于杂草胁迫对叶绿素影响未见报道。此次试验丝茅入侵显著降低了草坪草叶绿素含量,促进了叶片的老化。>20%侵入量显著降低了吸收和转化光能的Chla含量,大于30%入侵量显著降低了传递光能的Chlb含量,但侵入量对Chla/b无显著影响,说明3种草坪草均能利用“叶绿素循环”,在杂草胁迫下维持较高的相对稳定的光能利用效率。
有研究认为,高SP含量可维持较低的细胞渗透势,抵抗胁迫带来的伤害[20]。也有研究认为,胁迫抑制了蛋白质的合成并诱导蛋白质的降解,使植株体内的总蛋白质含量降低[21]。试验中,草坪草SP含量随丝茅侵入量不断增加,说明杂草胁迫使不溶性蛋白转变成可溶性蛋白以增强渗透调节能力,限制了草坪草蛋白质的合成。在逆境条件下,植物体内Pro大量积累,积累的脯氨酸除调节细胞渗透势外,在稳定生物大分子结构、降低细胞酸性、解除氨毒以及作为能量库调节细胞氧化还原势等方面起重要作用。试验中,Pro含量呈先增后减趋势,侵入量20%时急剧增加,大于30%侵入量后则不断下降。或许是在重度胁迫下,体内蛋白质只降解为SP,SP分解为可维持细胞完整性的结构性或功能性氨基酸,而降低了分解为Pro的速度。植物器官衰老或逆境下常发生膜脂过氧化作用[23],MDA是膜脂过氧化的分解产物,其含量可反映细胞膜的伤害程度[24-25]。草坪草叶片MDA含量随丝茅侵入量增加而不断增加,说明丝茅入侵对草坪草细胞膜造成伤害,不同草种MDA含量不同,表明草种间膜脂过氧化的反应程度和对杂草胁迫的耐受程度存在差异,草地早熟禾MDA含量最高受胁迫最严重。有研究认为,可溶性糖(SS)的渗透调节能力高于其他渗透调节物质,与其他渗透调节物质相比有一定的滞后性,在胁迫中、后期才开始积累[22],本试验中SS含量受丝茅侵入量影响小于SP和MDA,且变化幅度较小,不同草种间SS含量有显著差异,早熟禾SS含量最低。
丝茅庞大的根系系统与快速生长能力,强大的分生再生潜力和超强的种子结实与扩散力,通过地上、地下资源竞争对草坪草形成现实胁迫力,对草坪群落组成、结构及演替方向具有潜在攻击力。丝茅产生的次生代谢产物,引起土层含水量降低、酸度增高、通透性变差等变化[26],引起草坪草根系分生、更新、吸收能力下降,致使草坪植物受到更为复杂的胁迫。丝茅入侵降低了3草种叶色素含量,影响了草坪植物的光合同化能力;增加了抗氧化酶活性,将更多的能量用于抵御或应对胁迫,改变了草坪的物质能量分配及新陈代谢速度;提高了细胞膜透性,造成SP、SS、Pro和MDA流失,影响了细胞的健康和寿命。丝茅胁迫致使草坪生理生化发生变化,外在体现为草坪草根系浅表化、植株矮小化、生物量累积及分蘖能力下降。3种混播草种的生物学特征与生态学特点不同[27],对丝茅入侵的生理响应存在着显著种间差异,但>20%侵入量均显著降低了3种草坪草的物质累积与分蘖能力,最终必将破坏草坪群落组成、结构、功能,引起草坪退化,造成草坪外观质量、生态质量和使用质量下降。
混播草坪由建群种(>50%)、伴生种(30%)和保护种(10%)组成,依混播的目的性、兼容性、竞争性、主导性决定混播草种的比例及作用。混播草种生物学特征与生态学特点不同,株丛类型、生长方式和分生再生能力差异较大。为避免混播草种种间竞争与相互影响,试验分别用复合取代设计,研究了3种草坪草对丝茅入侵的生理响应能力,结果表明抗氧化酶活性、叶绿素含量、细胞膜透性及生长指标对丝茅入侵均有极显著响应,不同侵入量间有显著差异。虽早熟禾的SP、SS、Pro和MDA含量显著低于高羊茅和黑麦草,黑麦草抗氧化酶活性显著大于高羊茅和早熟禾,高羊茅叶绿素含量显著低于早熟禾和黑麦草,表明了3种草对丝茅入侵的生理响应差异,但无法综合表征3种草抗丝茅入侵能力的大小。有研究用模糊数学隶属度公式定量转换,对植物的抗逆性进行综合评定,用抗氧化酶活性、叶绿素含量、细胞膜透性等指标的隶属函数值的平均值进行抗性顺序[28],虽有一定的参考价值,但把不同属性不同重要性指标在没有加权指数情况下,仅用平均值进行比较似缺乏严谨性。丝茅入侵对草坪草造成立体、系统、持久的胁迫,在时间、空间与环境因子的契合作用下,草坪草生理代谢、形态结构、外观性状不断受到影响。所以,应依据“预防为主,综合防治”原则,分析杂草与环境的契合联系,对丝茅入侵途径、繁殖特点及危害规律进行深入研究,探讨防治丝茅入侵与扩散的有效方法与措施,是解决丝茅危害的根本出路。
[1]谭永钦,张国安,郭尔祥.草坪杂草生态位研究[J].生态学报,2004,24(6):1300-1305.
[2]强胜,李广英.南京市草坪夏季杂草分布特点及防除措施研究[J].草业学报,2000,9(1):48-54.
[3]刘金平,毛凯,游明鸿.草坪在生态城市建设中的作用及其应用[J].草业科学,2002,19(2):50-53.
[4]于风芝.冷季型草坪主要杂草发生规律及化学防除研究[J].草业科学,2000,17(2):39-42.
[5]熊琳.紫茎泽兰和4种牧草苗期相对竞争力的比较研究[D].重庆:西南大学,2009.
[6]朱宏伟,孟玲,李保平.黑麦草与入侵杂草紫茎泽兰苗期的相对竞争力[J].应用与环境生物学报,2007,13(1):29-32.
[7]冷琴,杨雅玲.国产白茅属部分生物学特性的研究[J].南京大学学报(自然科学),2002,38(5):703-714.
[8]张丽慧,赵艳,刘金平.修剪对丝茅草入侵麦冬草坪生物量结构及景观价值的影响[J].西南农业学报,2015,28(1):371-375.
[9]Willard T R,Shilling D G,Gaffney J F,etal.Mechanical and chemical control of cogongrass (Imperatacylindrica)[J].Weed Technol,1996,10:722-726.
[10]Otsamo R.Early development of three planted indigenous tree species and natural under storey vegetation in artificial gaps in anAcaciamangiumstand on anImperatacylindricagrassland site in South Kalimantan,Indonesia[J].New Forest,2000,19:51-68.
[11]刘金平,游明鸿.丝茅草侵入量与高羊茅相对竞争力及对幼坪性状影响的分析[J].草业学报,2012,21(6):315-320.
[12]李合生.植物生理生化实验原理和测定技术[M].北京:高等教育出版社,2003.
[13]熊庆娥.植物生理实验教材[M].成都:四川科学技术出版社,2003.
[14]王学奎.植物生理生化试验原理和技术[M].北京:高等教育出版社,2006.
[15]胡义,胡庭兴,陈洪,等.干旱胁迫及复水对香樟幼树生理特性及生长的影响[J].西北植物学报,2015,35(2):294-301.
[16]山仑,陈培元.旱地农业生理生态基础[M].北京:科学出版社,1998.
[17]刘冰,梁婵娟.生物过氧化氢酶研究进展[J].中国农学通报,2005,21(5):223-224.
[18]苏行,胡迪琴,林植芳,等.广州市大气污染对两种绿化植物叶绿素荧光特性的影响[J].植物生态学报,2002,26(5):599-604.
[20]韩蕊莲,李丽霞,梁宗锁.干旱胁迫下沙棘叶片细胞膜透性与渗透调节物质研究[J].西北植物学报,2003,23(1):23-27.
[21]白志英,李存东,刘渊.干旱胁迫下小麦叶片脯氨酸和蛋白质含量变化与染色体的关系[J].植物遗传资源学报,2007,8(3):325-330.
[22]刘红云,梁宗锁,刘淑明,等.持续干旱及复水对杜仲幼苗保护酶活性和渗透调节物质的影响[J].西北林学院学报,2007,22(3):55-59.
[23]刘延吉,张珊珊,田晓艳,等.盐胁迫对NHC牧草叶片保护酶系统,MDA含量及膜透性的影响[J].草原与草坪,2008(2):30-33.
[24]王太亮,韩兆胜,庞妙甜,等.NaCL胁迫对狗牙根生理生化特性的影响[J].草原与草坪,2015,30(1):63-67.
[25]杜利霞,董宽虎,乔志宏,等.NaCL胁迫对赖草幼苗生理特性的影响[J].草原与草坪,2013,33(1):1-4.
[26]刘金平,段婧.丝茅入侵对高羊茅根系及土壤性状的影响[J].草地学报,2012,20(5):870-875.
[27]孙吉雄.草坪学[M].北京:中国农业出版社(第二版),2006.
[28]孙凌霞,孙萍,蔡仕珍,等.四种冷季型草坪草对二氧化硫胁迫的生理响应研究[J].草业学报,2014,24(4):237-244.
Analysis on physiological response of three cool-season turfgrasses invaded byImperatakoenigii
MA Jiao,ZONG Ren-xu,LIU Jin-ping,WU De
(KeyLaboratoryofSouthwestChinaWildlifeResourcesConservation(MinistryofEducation),ChinaWestNormalUniversity,Nanchong637009,China)
Five invasive gradients were used to simulate the invading ofImperatakoenigiiin 3 cold-season turfgrass species communities (tall fescue,kentucky bluegrass and perennial ryegrass) through the composite replace experimental design.After 70 days,the antioxidant enzyme activity,cell membrane damage degree,photosynthetic pigment content and growth development were determined.The study was aimed to analyze the interspecific physiological differences of turfgrass invaded byI.koenigiiand damage differences of turfgrasses under the condition of different invasive ratio.The results showed that:1) The invasive ratio significantly increased SOD,POD and CAT activity of 3 species (P<0.01 ).But SOD,POD and CAT activity all existed significant difference among the invasive ratios.SOD and CAT activity had a significant difference among turfgrass species.2)InvadedI.koenigiidamaged turfgrass cell membrane permeability,and significantly increased soluble protein (SP),soluble sugar (SS),malondialdehyde(MDA) and proline (Pro) content (P<0.01).For turfgrass species,a significant difference was found between SP,SS,Pro and MDA contents,with the order of SS > MDA > SP > Pro.For the invasive ratio ofI.koenigii,the difference order was MDA > SP > SS > Pro.3) The invasive ratio ofI.koenigiisignificantly decreased Chla,Chlb,Chla+b contents (P<0.01).The difference order was Chla+b > Chla > Chlb > Chla/b among the invasive ratios ofI.koenigii,and the difference order was Chla > Chlb > Chla/b > Chla+b among turfgrass species.4) The invasive ofI.koenigiialso significantly decreased tiller number and single plant biomass (P< 0.01).5) More than 10% invasive ratio,SP,SS and Pro contents of turfgrass significantly increased;While the invasive ratio was more than 20%,the activity of SOD and POD and MAD content also significantly increased,but the contents of Chla and Chla+b,tiller number and single plant biomass decreased.Under more than 30% of invasive ratio,the content of CAT significantly increased,but Chla content decreased.The physiological response of three species to invadedI.koenigiiwas significant different.The damage degree was increased along with the increase of invasive ratio.Therefore,integrated control was the fundamental way to solve the problem.
Cool-season turfgrass;Imperatakoenigii;antioxidase enzyme;cell membrane permeability; photosynthetic pigments
2015-12-03;
2016-01-14
四川省科技支撑计划(2011NZ0064)项目资助
马娇(1992-),女,四川南充人,在读硕士。
E-mail:735284596@qq.com
S 688.4
A
1009-5500(2016)03-0035-07
刘金平为通讯作者。
猜你喜欢
奥秘(创新大赛)(2022年8期)2022-09-14
中国果业信息(2021年10期)2021-12-07
湖南农业大学学报(自然科学版)(2021年2期)2021-05-06
养殖与饲料(2020年6期)2020-02-18
农业与技术(2020年2期)2020-02-05
北方经济(2019年8期)2019-10-11
中国种业(2019年8期)2019-08-26
中国检察官·经典案例(2019年2期)2019-04-08
山西农经(2019年5期)2019-02-22
绿色科技(2017年13期)2017-07-31
