
变异链球菌耐酸毒力因子质子移位膜ATP酶在龋病进展中的动态变化
2016-07-08 04:42:54高敬黄文明重庆医科大学附属口腔医院牙体牙髓病科口腔疾病与生物医学重庆市重点实验室重庆市高校市级口腔生物医学工程重点实验室重庆401147
华西口腔医学杂志 2016年2期
高敬 黄文明重庆医科大学附属口腔医院牙体牙髓病科·口腔疾病与生物医学重庆市重点实验室·重庆市高校市级口腔生物医学工程重点实验室,重庆 401147
变异链球菌耐酸毒力因子质子移位膜ATP酶在龋病进展中的动态变化
高敬 黄文明
重庆医科大学附属口腔医院牙体牙髓病科·口腔疾病与生物医学重庆市重点实验室·
重庆市高校市级口腔生物医学工程重点实验室,重庆 401147
[摘要]目的 研究变异链球菌耐酸毒力因子质子移位膜ATP酶(F-ATPase)在不同pH环境和龋病发生发展过程中的表达,评价F-ATPase在龋病进展中的动态变化。方法 将变异链球菌菌悬液在不同pH(pH4.0~7.0)和不同葡萄糖浓度(含5%和不含葡萄糖)的BHI液体培养基中培养,检测F-ATPase基因的表达水平。将雄性Wistar大鼠随机分成致龋组和对照组,其中致龋组喂养致龋饲料及5%葡萄糖水,对照组喂养普通饲料。每2周采集菌斑样本,检测F-ATPase基因的表达水平。第11周时取大鼠上下颌骨标本,对磨牙进行龋损评定。结果 1)5%葡萄糖浓度下变异链球菌F-ATPase基因的表达高于不含葡萄糖(P<0.05),在pH5.0时F-ATPase基因的表达最高,pH4.0时表达最低(P<0.05)。2)成功构建了龋病动物模型,在龋病发生发展过程中,致龋组和对照组F-ATPase的表达逐渐增强,致龋组表达高于对照组(P<0.05)。结论 耐酸毒力因子F-ATPase的表达变化与龋病的发生发展密切相关。
[关键词]龋病; 牙菌斑; 质子移位膜ATP酶; pH
耐酸性是细菌在致龋过程中最重要的毒力表现,细菌产酸后若不能耐酸,则细菌就不能继续生存、代谢。Bender等[1]研究发现细菌的耐酸性与细菌膜对质子的透过性有关,变异链球菌的耐酸性主要依赖于细胞膜上的质子移位膜ATP酶(membranebound proton-translocating ATPase,F-ATPase),通过F-ATPase将质子泵出胞内,使胞内pH(pHi)高于胞外pH(pHo),即维持了一个跨膜梯度ΔpH (pHi-pHo),保持胞内pH接近中性,保护胞浆中与糖酵解相关酶的活性,使菌细胞在低pH的苛刻环境中仍能产酸、生存并最终导致龋病的发生[2-3]。可见,F-ATPase是变异链球菌致龋菌耐酸的引擎和马达,与龋病的发生密切相关。
目前尚缺乏关于变异链球菌耐酸毒力因子FATPase在龋病发生发展过程中的研究,为了进一步了解F-ATPase在龋病进展中的变化,本研究建立致龋动物模型纵向研究F-ATPase基因在龋病进展中的表达变化,比较F-ATPase基因在不同pH环境的表达,以评价生物膜中耐酸毒力因子F-ATPase与龋病发生的关系。
1 材料和方法
1.1 实验菌株及动物
变异链球菌(Streptococcus mutans,S. mutans)标准株UA159由四川大学口腔疾病研究国家重点实验室提供。18 d龄的雄性Wistar大鼠购于重庆医科大学实验动物中心,许可证编号为SYXK(渝)2012-0001。
1.2 主要试剂及仪器
BHI培养基(OXOID公司,英国),YJ-875型超净工作台(苏州市华宇净化设备有限公司),FQ-聚合酶链反应(polymerase chain reaction,PCR)仪(Bio-Rad公司,美国) ,200型分光光度计(Thermo Fisher公司,美国),pH计(Mettler Toledo公司,瑞士),Bio-Rad电泳仪(Bio-Rad公司,美国)。
1.3 引物设计
根据GenBank报道的变异链球菌F-ATPase β亚基基因[4]序列及16sRNA基因序列(GenBank accession number NC_004350),采用Primer primer 5.0计算机软件设计引物。F-ATPase上游引物 5’-CGGATGCGTGTTGCTCTTACTG-3’,下游引物5’-GGCTGATAACCAACGGCTGATG-3’;16sRNA上游引物5’-TGGAACTGAGACACGGTCCA-3’,下游引物5’-CGCTTTACGCCCAGTAATTCCG-3’。引物由上海捷瑞生物工程有限公司合成。
1.4 不同pH和葡萄糖浓度下变异链球菌F-ATPase基因的表达
将变异链球菌标准株UA159复苏,培养24 h,3 000 r·min-1离心15 min,弃上清,用无菌生理盐水于紫外分光光度计540 nm处制备成吸光度为1.0的菌悬液备用。以0.5为间隔配制pH4.0~7.0系列的含5%葡萄糖和不含葡萄糖的BHI液体培养基。取变异链球菌悬液,按菌液与BHI液体培养基1∶10的体积比接种细菌,37 ℃厌氧培养至16 h指数生长期,每个样品一式3份,3 000 r·min-1离心15 min收集菌体,通过RNAiso Plus(Takara公司,日本)提取RNA,逆转录成cDNA,PCR扩增反应总体积为20 μL,含SYBR Premix Ex Taq(2x)10 μL(包括DNA Taq酶、Mg2+、dNTP、SYBR GreenⅠ荧光染料和Buffer缓冲液)、上下游引物各0.5 μL、模板cDNA 2 μL,加灭菌去离子水至20 μL。反应循环参数设置为三步法PCR扩增标准程序:95 ℃预变性3 min,95 ℃变性30 s,60 ℃退火30 s,72 ℃延伸30 s,共40个循环。在延伸阶段收集荧光信号,循环结束后附加溶解曲线分析:95 ℃ 15 s,62 ℃ 23 s,95 ℃ 15 s。实验重复3次,取均值,收集实验数据,测定各样品的循环阈值(Ct值),通过2-ΔΔCt计算目的基因的相对表达量。
1.5 龋病动物模型的建立及变异链球菌F-ATPase基因表达检测
12只18 d龄的雄性Wistar大鼠,随机分为2组,致龋组6只,喂养致龋饲料2 000号及5%葡萄糖水;对照组6只,喂养普通饲料。从第1周开始每2周采集菌斑样本(共6次),采集样本前2 h大鼠禁食,腹腔注射10%水合氯醛麻醉,以棉拭子采集磨牙面及颊舌(颊腭)面的菌斑,置于含硫乙醇酸盐传送液的离心管中洗脱,将样本收集于离心管中,提取混合菌斑RNA,进行逆转录和实时荧光定量PCR(引物及反应体系同上)。结果取均值,通过2-ΔΔCt计算目的基因的相对表达量。
第11周时处死大鼠,取上下颌骨标本,染色,根据Keyes经典评分方法[5]对磨牙进行龋损评定,将龋损分为4级。E级:仅限于釉质;Ds级:累及釉质及牙本质外层1/4以内;Dm级:累及牙本质厚度的1/4至3/4;Dx级:累及超过牙本质厚度的3/4,甚至达牙本质全层。
1.6 数据分析
采用SPSS 17.0软件进行统计分析,所有数据均以均数±标准差表示,多组之间比较采用单因素方差分析。
2 结果
2.1 变异链球菌F-ATPase基因及16sRNA内参基因
F-ATPase基因和16sRNA内参基因经常规PCR都能得到特异性扩增,2%凝胶电泳显示各基因各实验组中的cDNA产物均为单一条带(图1、2),条带片段大小与引物预期扩增片段大小一致。
图 1 F-ATPase基因扩增产物电泳图Fig 1 Electrophoresis of products of F-ATPase gene
2.2 不同pH和葡萄糖浓度下变异链球菌F-ATPase基因的表达
不同pH和葡萄糖浓度下变异链球菌F-ATPase基因的表达见图3。实验结果显示,1)在pH5.0时FATPase基因的表达最高,然后依次是pH5.5、6.0、6.5,7.0,4.5,最低的是pH4.0(P<0.05);2)5%葡萄糖浓度下F-ATPase的表达高于不含葡萄糖(P< 0.05)。
2.3 大鼠磨牙龋病动物模型的建立
11周后处死大鼠,取磨牙进行Keyes评分,结果见图4和表1。实验成功构建了龋病动物模型,各组Wistar大鼠磨牙均有不同程度的龋损发生,但是龋损发生的严重程度、累及范围不尽相同。致龋组龋齿计分明显高于对照组,且龋坏病变的程度要高于对照组。对照组牙齿的损坏最小,仅达到牙本质中层,致龋组龋损多数达牙本质中层,其次是牙本质深层。
图 2 16sRNA基因扩增产物电泳图Fig 2 Electrophoresis of products of 16sRNA gene
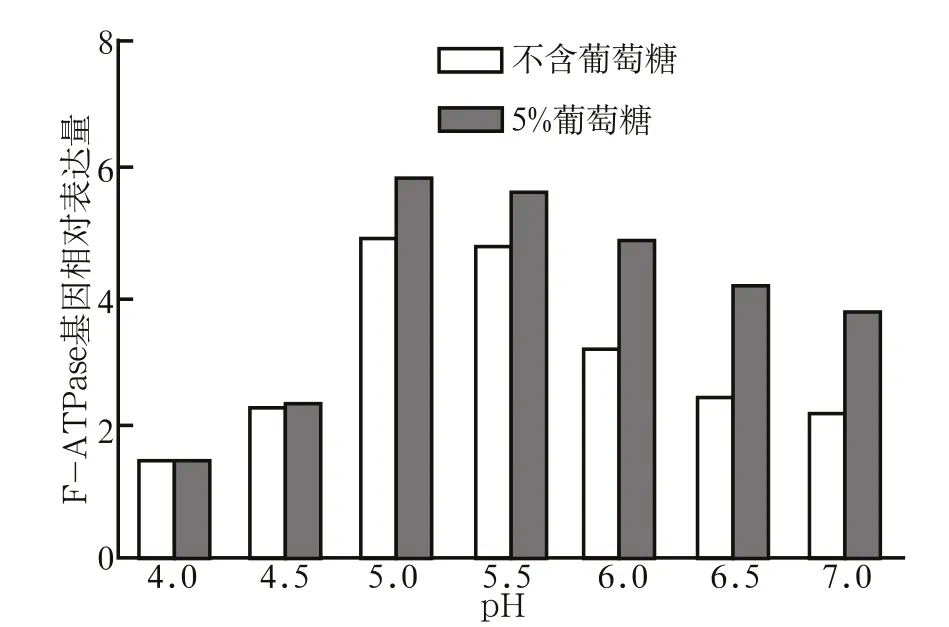
图 3 不同pH和葡萄糖浓度下变异链球菌F-ATPase基因的表达 Fig 3 F-ATPase expression under different pH and glucose con- centration
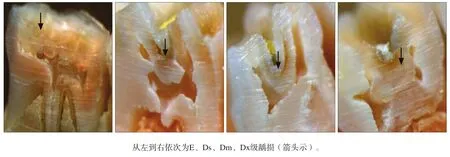
图 4 大鼠龋齿 体视显微镜 × 30Fig 4 The caries of rats stereomicroscope × 30
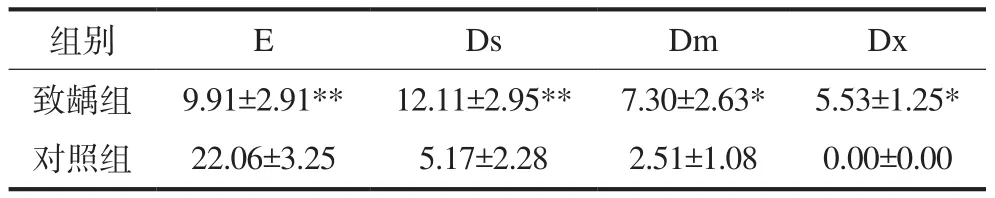
表 1 各组大鼠磨牙龋齿Keyes评分Tab 1 The Keyes score in the molar teeth of each rat group
2.4 龋病过程中F-ATPase基因的表达
龋病过程中各组F-ATPase基因的表达见表2。在龋病的发生发展过程中,随时间的推移各组F-ATPase基因的表达逐渐增强,致龋组表达高于对照组(P< 0.05),第9周后,致龋组增长程度有一定下降,对照组的表达下降。
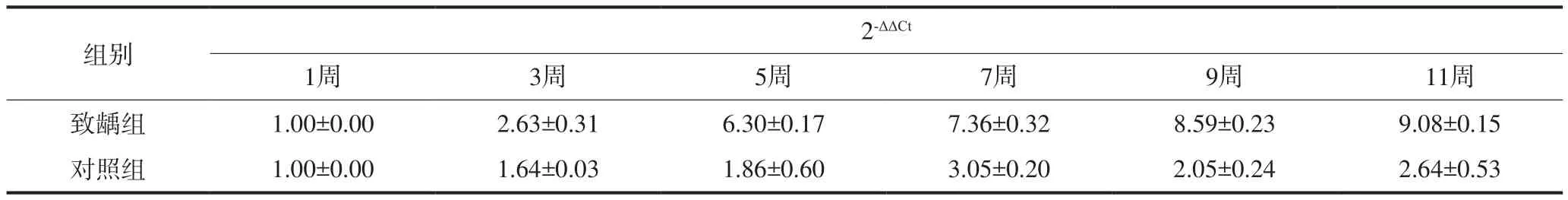
表 2 龋病过程中各组F-ATPase基因的表达Tab 2 F-ATPase expression in each group during the de velopment of caries
3 讨论
F-ATPase具有调节胞浆pH以及决定菌细胞耐酸性的生物学特性。细菌细胞的胞外质子向胞内扩散降低胞内pH,影响对酸敏感的糖酵解的活性,为了最大程度的发挥糖酵解的活性,致龋菌通过细胞膜和F-ATPase抵抗胞浆的酸化,F-ATPase将质子泵出细胞从而调节pH,使致龋菌在低pH值环境下仍能进行糖酵解[6-7]。
早期龋病研究表明,环境pH和糖在龋病的发生发展过程中起着重要的作用,而口腔原位环境是一个复杂且动态变化的环境,获取大鼠致龋过程中牙菌斑pH较困难,为了研究高糖浓度下变异链球菌FATPase基因的表达与pH之间的关系,本研究设计了体外实验,结果显示,变异链球菌在5%葡萄糖浓度下耐酸毒力因子F-ATPase表达高于不含葡萄糖环境。研究[8]表明,在高糖浓度下,变异链球菌的黏附毒力、产酸毒力、耐酸毒力及耐药毒力均高于低糖或无糖环境下,由此推测,高糖浓度下变异链球菌致龋能力上调。当外界环境中有充足的糖源时,变异链球菌能迅速合成胞外多糖,伴随着产酸耐酸能力的增强,环境中pH迅速下降,变异链球菌在酸压力状态下通过上调耐酸因子F-ATPase表达来维持生存代谢。Kuhnert等[4]对浮游状态的变异链球菌F-ATPase基因表达研究证实,在pH5.0时mRNA的表达水平大约是pH7.0时的2倍。研究[9]发现,当培养基中的pH 由8.0下降至5.0时,变异链球菌的F-ATPase活性提高4倍,从而增强了其质子泵作用。本实验研究也证实,高糖浓度下变异链球菌F-ATPase基因的表达高于无糖或低糖环境,F-ATPase的表达在pH5.0时最高,pH4.0时最低,pH5.0到7.0时F-ATPase表达缓慢降低,在pH5.0以下时F-ATPase的表达急剧减少。学者[10]研究对比了干酪乳杆菌、变异链球菌和血链球菌3种耐酸性分别为高、中、低的口腔耐酸菌膜结合F-ATPase的pH活性范围,发现其最适pH值分别为5.5、6.0和7.0,证明了细菌的相对耐酸性与F-ATPase的活性和最适pH值有关,活性越大,最适pH值越低。在酸性环境中排除胞内多余H+能力越强,因而越耐酸。可见在最适pH以内,F-ATPase的表达随pH降低而升高,而当pH降低至5.0以下时,酶活性超过最适pH使F-ATPase的表达量下降,同时强酸的环境压力可以使细菌合成ATP不足,并破坏细胞质内的蛋白结构使之死亡[11]。通过对大肠杆菌的研究,Kasimoglu等[12]发现,F-ATPase的表达量在转录和翻译水平的调控是多层次的,其中细菌的生长率在F-ATPase的产生和表达中起着比较重要的作用,大量细菌死亡也会导致生物膜中F-ATPase的表达量大大减少。同样在pH5.0以下的环境,随着变异链球菌的大量死亡,F-ATPase的基因表达在高糖和无糖环境中差异较小。
龋齿Keyes评分结果显示,致龋组龋齿计分明显高于对照组,且龋坏病变的程度要高于对照组,证明本实验龋齿模型的建立成功。实验中所使用的致龋饲料为高糖饲料,大鼠牙菌斑长期暴露在高糖的环境中,致龋菌有足够的底物产酸,造成牙菌斑内pH持续下降,在龋病发生发展过程中,致龋组和对照组F-ATPase表达均逐渐增强,致龋组表达高于对照组,第9周后,致龋组增长程度有一定下降,对照组的表达有一定下降。在实验过程中第5周时大鼠龋坏开始出现,在龋坏启动前期,变异链球菌等致龋菌大量产酸,使菌斑内pH达到脱矿临界值pH5.0~ 5.5。F-ATPase是一种应激蛋白,当外界环境pH下降时,变异链球菌F-ATPase基因表达上调,从而合成更多的F-ATPase蛋白以排除更多的氢离子维持细菌的生存,当更多的氢离子排到外环境时,环境中的pH继续下降酸蚀牙齿致龋。当龋病发展到第9周时,牙菌斑内微生物已经形成一个稳定的动态平衡状态,随着唾液蛋白等的中和缓冲作用,牙菌斑内pH值上升[13],F-ATPase表达下调。学者[14]分离临床牙菌斑中变异链球菌,结果表明高龋牙菌斑变异链球菌分离株黏附、产酸和耐酸能力高于无龋牙菌斑分离株。本实验中致龋组耐酸因子表达亦高于对照组。对照组大鼠使用的普通饲料内碳水化合物含量较低,大鼠本身的口腔自洁能力较强,唾液分泌较多,对菌斑内pH缓冲能力强,使菌斑pH维持在一个稳定的范围,F-ATPase的表达也维持在一个稳定的范围。
本文对变异链球菌耐酸毒力因子F-ATPase在不同pH中的基因表达进行了研究,并建立了大鼠致龋模型,结果显示,变异链球菌F-ATPase在pH5.0时表达最高,致龋组大鼠在龋病发展过程中F-ATPase表达上调,提示耐酸毒力因子F-ATPase在釉质脱矿及龋病的形成中发挥着重要的作用。
[参考文献]
[1] Bender GR, Sutton SV, Marquis RE. Acid tolerance, proton permeabilities, and membrane ATPases of oral streptococci [J]. Infect Immun, 1986, 53(2):331-338.
[2] Matsui R, Cvitkovitch D. Acid tolerance mechanisms utilized by Streptococcus mutans[J]. Future Microbiol, 2010, 5(3):403-417.
[3] Lemos JA, Quivey RG Jr, Koo H, et al. Streptococcus mutans: a new Gram-positive paradigm[J]. Microbiology, 2013, 159(Pt 3):436-445.
[4] Kuhnert WL, Zheng G, Faustoferri RC, et al. The F-ATPase operon promoter of Streptococcus mutans is transcriptionally regulated in response to external pH[J]. J Bacteriol, 2004, 186(24):8524-8528.
[5] Keyes PH. Dental caries in the molar teeth of rats. Ⅱ. A method for diagnosing and scoring several types of lesions simultaneously[J]. J Dent Res, 1958, 37(6):1088-1099.
[6] 杨德琴, 刘天佳, 付春华, 等. 变形链球菌F-ATPase亚基基因uncEBF基因多态性的研究[J]. 华西口腔医学杂志, 2007, 25(3):219-222. Yang DQ, Liu TJ, Fu CH, et al. Genetic diver sity of FATPase subunits gene uncEBF amplified from Streptococcus mutans clinical isolates[J]. West China J Stomatol, 2007, 25(3):219-222.
[7] Argimón S, Caufield PW. Distribution of putative virulence genes in Streptococcus mutans strains does not correlate with caries experience[J]. J Clin Microbiol, 2011, 49(3): 984-992.
[8] Sun M, Kang Q, Li T, et al. Effect of high-fructose corn syrup on Streptococcus mutans virulence gene expression and on tooth demineralization[J]. Eur J Oral Sci, 2014, 122 (3):216-222.
[9] Nakanishi-Matsui M, Sekiya M, Nakamoto RK, et al. The mechanism of rotating proton pumping ATPases[J]. Biochim Biophys Acta, 2010, 1797(8):1343-1352.
[10] Sturr MG, Marquis RE. Comparative acid tolerances and inhibitor sensitivities of isolated F-ATPases of oral lactic acid bacteria[J]. Appl Environ Microbiol, 1992, 58(7):2287-2291.
[11] MacGilvray ME, Lapek JD Jr, Friedman AE, et al. Cardiolipin biosynthesis in Streptococcus mutans is regulated in response to external pH[J]. Microbiology, 2012, 158(Pt 8): 2133-2143.
[12] Kasimoglu E, Park SJ, Malek J, et al. Transcriptional regulation of the proton-translocating ATPase (atpIBEFHAGDC) operon of Escherichia coli: control by cell growth rate[J]. J Bacteriol, 1996, 178(19):5563-5567.
[13] van Ruyven FO, Lingström P, van Houte J, et al. Relationship among mutans streptococci, “low-pH” bacteria, and lodophilic polysaccharide-producing bacteria in dental plaque and early enamel caries in humans[J]. J Dent Res, 2000, 79(2):778-784.
[14] Botelho MP, Maciel SM, Cerci Neto A, et al. Cariogenic microorganisms and oral conditions in asthmatic children[J]. Caries Res, 2011, 45(4):386-392.
(本文编辑 李彩)
[中图分类号]R 780.2
[文献标志码]A [doi] 10.7518/hxkq.2016.02.019
[收稿日期]2015-07-12; [修回日期] 2015-10-18
[基金项目]国家自然科学基金(31371473);重庆市卫生局医学科研计划(2012-2-129);重庆市卫生局医学科研计划重点项目(2011-1-062);重庆市渝北区科技项目(2011)
[作者简介]高敬,硕士,E-mail:326188065@qq.com
[通信作者]黄文明,讲师,硕士,E-mail:jxhwm@163.com
Dynamic changes of aciduric virulence factor membrane-bound proton-translocating ATPase of Streptococcus mutans in the development of dental caries
Gao Jing, Huang Wenming. (Dept. of Conservative Dentistry and Endodontics, Stomatological Hospital of Chongqing Medical University; Chongqing Key Laboratory of Oral Diseases and Biomedical Sciences; Chongqing Municipal Key Laboratory of Oral Biomedical Engineering Higher Education, Chongqing 401147, China)
Supported by: Natural Science Foundation of China (31371473); Medical Scientific Research Plan of Chongqing Municipal Health Bureau (2012-2-129); Key Program for Medical Scientific Research Plan of Chongqing Municipal Health Bureau (2011-1-062); Scientific and Technical Program of Chongqing Yubei District (2011). Correspondence: Huang Wenming, E-mail: jxhwm@163.com.
[Abstract]Objective To observe the dynamic changes of membrane-bound proton-translocating ATPase (F-ATPase) in the development of dental caries, the expression of Streptococcus mutans F-ATPase under different pH concentrations and during the development of dental caries is analyzed. Methods Streptococcus mutans cultured under different pH (pH4.0-7.0) concentrations and containing 5% glucose and no glucose containing BHI were collected. RNA was extracted. Subsequently, F-ATPase gene was detected using real-time polymerase chain reaction. Male Wistar rats were divided randomly into caries group and control group. The rats in the caries group were fed caries feed and 5% glucose water, whereas those of control group were fed normal feed. Total RNA was extracted from plaque samples, which were collected from rats’ oral cavity every two weeks. F-ATPase gene was detected by real-time PCR. In the 11th week, the upper and lower jaw bone specimens of rats were taken, and molar caries damage assessed. Results The expression of F-ATPase in the caries group was higher than that in the control group (P<0.05). In addition, the gene was expressed highest in pH5.0 and the lowest in pH4.0 (P<0.05). 2) The expression of F-ATPase progressively increased during the caries development in both groups; expression in the caries group was higher than that in control group (P<0.05). Conclusion Acid-resisting viru-lence factor F-ATPase is related closely with the incidence and development of dental caries.
[Key words]dental caries; dental plaque; membrane-bound proton-translocating ATPase; pH
猜你喜欢
护理实践与研究(2022年4期)2022-01-01 07:02:44
天津医科大学学报(2021年1期)2021-01-26 00:57:16
当代医药论丛(2020年20期)2020-12-02 04:09:24
广告大观(2020年4期)2020-10-20 14:02:55
健康之家(2019年9期)2019-12-14 03:44:43
意林·少年版(2019年11期)2019-06-30 23:24:00
问健康画报(2017年2期)2017-07-01 01:35:54
中国卫生标准管理(2015年17期)2016-01-20 09:26:40
实用临床医学(2014年6期)2014-02-28 09:18:31
中华养生保健(2013年12期)2013-12-16 01:34:18
