
新疆不同草地类型植物物种特征与水热因子的关系研究
2016-07-05 08:01刘利利盛建东程军回刘耘华李瑞霞赵丹新疆农业大学草业与环境科学学院新疆土壤与植物生态过程重点实验室新疆乌鲁木齐830052
草业学报 2016年5期
刘利利,盛建东,程军回,刘耘华,李瑞霞,赵丹(新疆农业大学草业与环境科学学院,新疆土壤与植物生态过程重点实验室,新疆 乌鲁木齐 830052)
LIU Li-Li,S H E N G Jian-Dong*,C H E N G Jun-H ui,LIU Y un-H ua,LI Rui-Xia,Z H A O DanXinjiang Key Laboratory of Soil and Plant Ecological Processes,College of Grassland and Environmental Sciences,Xinjiang Agricultural University,Urumqi 830052,China
新疆不同草地类型植物物种特征与水热因子的关系研究
刘利利,盛建东*,程军回,刘耘华,李瑞霞,赵丹
(新疆农业大学草业与环境科学学院,新疆土壤与植物生态过程重点实验室,新疆 乌鲁木齐 830052)
本文以新疆7种草地为研究对象,调查364个样地的物种名录,分析不同草地植物科属种的变化特征,探讨了植物物种丰富度(SR)对年平均温度(M A T)、降水(M A P)、海拔的响应。研究结果表明,新疆草地以禾本科、菊科为优势科,优势属为针茅属、绢蒿属;物种丰富度变化为:温性草甸草原(7.73)>温性草原(5.04)>高寒草原(4.36)>温性荒漠草原(3.99)>温性草原化荒漠(3.02)>温性荒漠(2.08)>低平地草甸(1.43);全部样地和温性草原的SR与M A P呈正相关,与M A T、海拔呈峰型关系;低平地草甸和高寒草原的SR与M A P呈正相关而与M A T呈负相关,与海拔呈先降低再增加的趋势;温性荒漠草原的SR与M A P呈正相关,与M A T、海拔无相关性;温性荒漠的SR与M A P呈正相关,与M A T呈负相关,与海拔呈峰型关系;温性草甸草原和温性草原化荒漠的SR 与M A T、M A P均无相关性。不同草地类型对温度、降水响应的差异,是由于植被类型和温度、降水的空间分异性,对于海拔的分异,来源于海拔梯度范围和所研究地区大尺度的气候特征。
海拔;草地;年平均降水;年平均温度;物种丰富度
http://cyxb.lzu.edu.cn
刘利利,盛建东,程军回,刘耘华,李瑞霞,赵丹.新疆不同草地类型植物物种特征与水热因子的关系研究.草业学报,2016,25(5):1-12.
LIU Li-Li,S H E N G Jian-Dong,C H E N G Jun-H ui,LIU Y un-H ua,LI Rui-Xia,Z H A O Dan.Relationship between plant species characteristics and clim ate factors in different grassland types of Xinjiang.Acta Prataculturae Sinica,2016,25(5):1-12.
LIU Li-Li,S H E N G Jian-Dong*,C H E N G Jun-H ui,LIU Y un-H ua,LI Rui-Xia,Z H A O Dan
Xinjiang Key Laboratory of Soil and Plant Ecological Processes,College of Grassland and Environmental Sciences,Xinjiang Agricultural University,Urumqi 830052,China
在中国草地生态系统中,新疆草地资源是绿洲农业系统赖以生存和发展的生态屏障,其总面积为5725.88× 104h m2,仅次于西藏和内蒙古,新疆草地不仅是中国草地的代表,同时也是中亚地区的典型,其草地类型多样,包括了诸如山地草甸、高寒草甸、温性草原、温性荒漠等中国主要的草地类型。但是,由于不合理放牧[1-2]、海拔[3-4]、土壤因子[5]、降水[6-7]、温度[8]等外界原因,造成草地植物物种多样性趋于丧失和退化的状态,而这种物种组成和多样性发生改变的结果,会造成生态系统的结构和功能产生一定的变化,还会对相邻区域的生态系统造成影响,更严重的会改变整个区域的生态过程[9-10]。
在全球气候变化的大背景下,全球平均气温持续波动上升,极端降水事件频发,降水格局发生变化[11],同时,生物多样性也面临着生态系统功能需求不断增加带来的挑战[12],这使更多的学者关注气候因子对物种多样性的影响。温度影响种子的萌发[13]以及土壤水分的蒸散,同时也是控制植物群落动态的主要因素[14],Alward等[15]发现温度能够显著影响植物物种水平上的生产力,例如,随着温度的增加,草地非禾本植物的数量有明显的增长[14]。而Engler等[16]和Dirnbck等[17]发现,气温、降雨等气候因素的改变将导致众多高山植物丧失适宜的生境,植物多样性大幅减少,但K napp等[18]在K onza草原的研究发现降水与物种多样性之间存在正相关,并不是物种丰富度,也有研究认为,由于草原对降水量以及频度和持续时间反应敏感,降水的变化将影响草原的物种组成、养分和碳循环速率与水的关系,并且可以看到草原的碳汇转变为碳源[19-20]。其次,海拔的变化在一定程度上代表了气候的变化,控制了水热条件引起的物种选择、资源竞争、生境变化,也是影响物种多样性的重要因素[21]。同时,有学者提出了重要的水分-能量动态理论,其主张物种丰富度的地理格局受水分和能量共同有效性的限制[22]。以上不同的研究结果表明,环境因子从根本上导致了物种多样性的变化,然而由于不同的草地类型和研究区域,使其并没有形成共识,仍需做进一步探讨。
新疆草地在中国草地生态系统中占有重要地位,冯缨等[23]对新疆天山北坡不同草地类型物种组合格局及其环境作用进行了分析,郭正刚等[24]探讨了新疆阿勒泰地区不同草地类型群落特征以及海拔对其多样性的影响,另有Li等[25]对新疆植物进行了大量的研究,发现水分蒸散量与物种丰富度之间呈显著正相关,植物物种丰富度与年降水量呈正相关,与温度呈先增加再降低的趋势[26]。但是物种变化是一个长期的过程,该地区尚缺乏植物自然适应气候变化过程中,生物群落属性(如群落物种组成、物种丰富度)和非生物因素(如温度、降水、海拔)的系统研究,本研究通过对新疆不同草地类型植被调查,结合气候因子的相关资料,探讨新疆草地植物的物种组成,同时分析草地植物物种丰富度对温度、降水、海拔的响应。
1 材料与方法
1.1 研究区域概况
本研究区域位于我国新疆维吾尔自治区,其南北长约1500 k m,东西长约1900 k m,总面积为166×104k m2,约占全国面积的1/6。新疆气候属于典型的温带大陆干旱性气候,光热资源充足,日照时数达2550~3500 h,无霜期长达180~220 d,降水量稀少。年平均降水量为146 m m,降水多集中于6-8月份,年平均气温9~12℃。最低平均温度为-26℃(1月),最高温度为33℃(7月)。该区域以荒漠土、棕钙土、栗钙土、黑钙土、盐碱土等为主。
草地是新疆的主要植被类型之一,其总面积为5725.88×104h m2,占该区域总面积的34.44%。按照草地类型划分方法,新疆草地划分为低平地草甸类、高寒草甸类、高寒草原类、温性草原化荒漠类、温性草甸草原类、温性草原类、温性荒漠草原类、温性荒漠类、温性山地草甸类、沼泽类这10种草地类型[27]。
1.2 样地设置
根据1∶100万的《中国植被图》所标记的草地类型和空间分布情况,在2011,2012和2013年对473个样地,分别进行了物种丰富度的调查,这些样地均未受到明显的人为干扰且代表了新疆草地的主要群丛和植被类型。为了消除不同样方大小对物种丰富度的影响,本文仅选用364个标准样地(样方的面积均为1 m×1 m)进行分析(图1)。不同草地基本特征如表1所示。
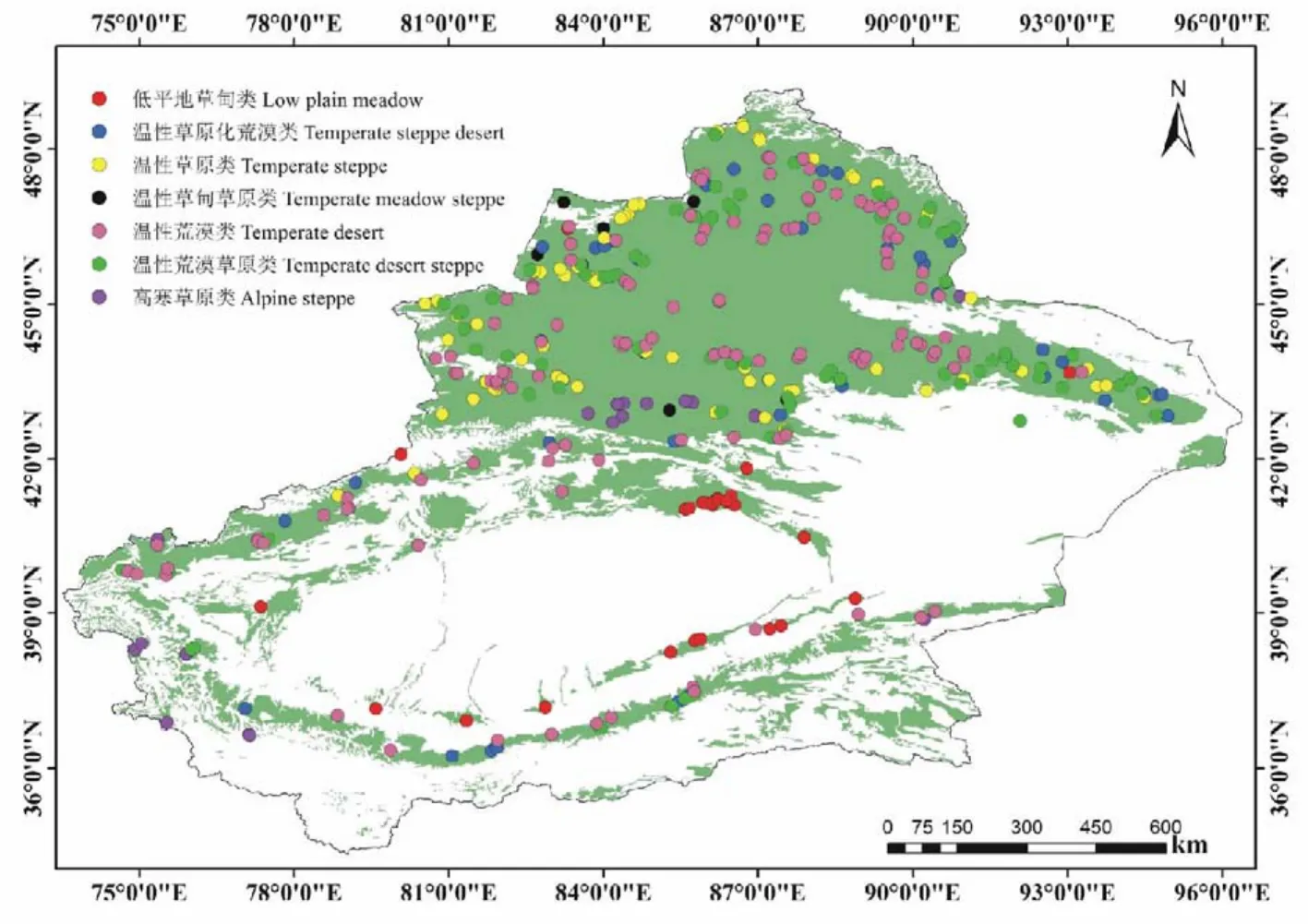
图1 新疆草地样地分布Fig.1 Sites distribution in Xinjiang grasslands
表1 不同草地类型基本特征Table 1 Basic characteristics in different types of grassland
1.3 物种丰富度调查
物种丰富度的调查均在2011-2013年7月中旬至8月下旬,植物生长达到最大生物量时进行。在每个样地内,选择未受人类活动干扰的区域随机设置一个100 m×100 m的调查区域。在其对角线上设置一条100 m样线,在调查中沿样线设置10个1 m×1 m样方,主样地为5个分种样方,顺序为2,4,6,8,10,辅样地为3个分种样方,顺序为2,6,10,分别记录每个样方中的物种名录,该名录数量得到平均值即为物种丰富度。
1.4 数据来源
海拔数据是在每个样点用G PS测定所得,而为了获取每个样地一一对应的精确年平均降水(m ean annual precipitation)、年平均温度(m ean annual te m perature)数据,首先从新疆均匀分布的气象站获得(中国国家气象局气候数据库)气象数据,然后利用每个样点的经度、纬度、海拔信息为预测因子,进行多元回归方法,采用地理信息系统(GIS)插值法得到每个样点温度、降水的数据[25]。
1.5 统计分析
本文采用非平衡单因素方差分析(U nbalanced one-way A N O V A)和最小显著差异法(LSD)来比较不同草地类型间不同水平上植物科属种的变化特征。通过线性和非线性拟合来反映不同草地类型中对降水、温度的响应。本文所有分析中,显著性水平为P<0.05。
2 结果与分析
2.1 不同草地类型植物科属频数
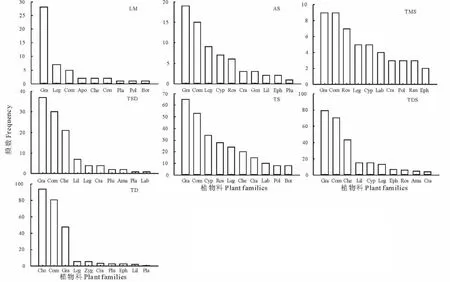
图2 不同草地类型植物科频数统计(位于前十)Fig.2 Plant families frequency statistics in different grassland types(in the top ten)
在调查的364个样地中,对不同草地位于前10的植物科频数统计发现(图2),禾本科为最大的优势科,除温性荒漠以藜科为最大的优势科外,在其他草地中均以禾本科出现频数最高;菊科为次优势科,除了低平地草甸外在其他草地中均表现出次优势特征;第三大优势科为豆科,在所有草地中均表现出相对重要地位;其次,以莎草科和车前科分布相对广泛。
在所有草地中,针茅属为最大优势属,在多数草地上(高寒草原、温性草甸草原、温性草原、温性荒漠草原)都是出现频数最高;第二大优势属为绢蒿属,分布较广,除低平地草甸和温性草甸草原外,在其他草地上均表现出突出优势,且在温性荒漠上为频数最高的优势属,其另一重要属为角果藜属;另外,苔草属在高寒草原、温性草甸草原和温性草原上也占有重要地位;而低平地草甸上芦苇属表现出最大优势,甘草属次之(图3)。
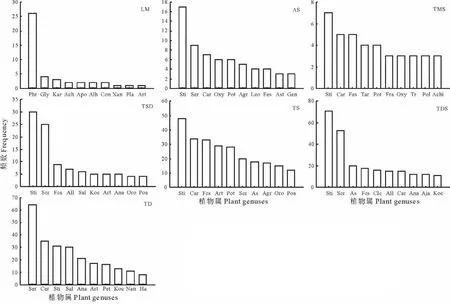
图3 不同草地类型植物属频数统计(位于前十)Fig.3 Plant genuses frequency statistics in different grassland types(in the top ten)
2.2 不同草地类型间植物科属种的变化特征
单因素方差分析结果显示,温性草甸草原拥有最多的科,平均为4.77个,远高于其他草地类型(P<0.05);温性草原和高寒草原科数量分别为3.43和3.33个,与其他草地之间均有显著差异;温性荒漠草原(2.75个)和温性草原化荒漠(2.37个),数量相当;温性荒漠和低平地草甸的科最少,分别为1.60和1.35,均显著低于其他草地类型(图4a)。
属最多的草地为温性草甸草原,平均为6.4个,高于其他草地;温性草原(4.61个)和高寒草原(3.92个)之间数量相当;温性荒漠草原属为3.57个,温性草原化荒漠样地平均为2.74个属,温性荒漠有1.92个属,此3种草地间有显著差异;而温性荒漠和低平地草甸(为1.38个)植物属之间无显著差异,而与其余5种草地均有显著差异(图4b)。
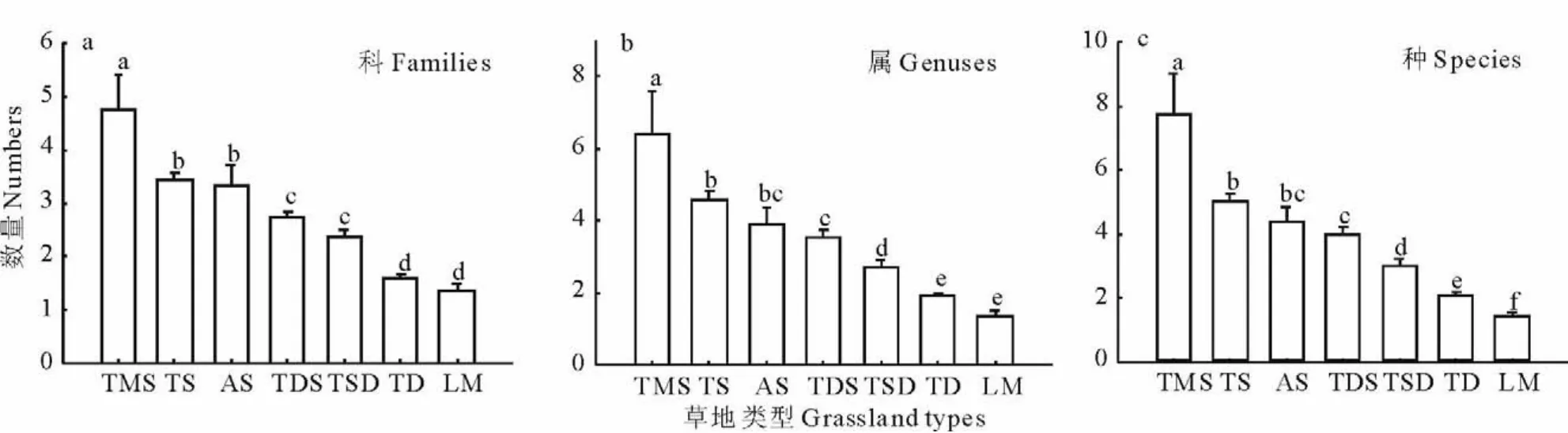
图4 不同草地类型间植物科属种的变化特征Fig.4 Variation of plant families,genuses,species cross different grassland types in Xinjiang
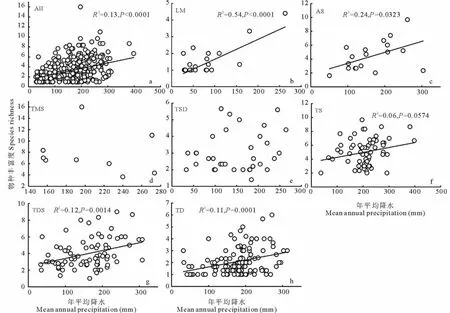
图5 不同植被类型物种丰富度与年平均降水量之间的关系Fig.5 Relationships between species richness and mean annual precipitation on different grasslands types
温性草甸草原拥有最高的物种丰富度,为7.73个物种,与其他草地间有显著差异;温性草原物种丰富度(5.04个物种),与高寒草原(4.36个物种)之间也无显著差异,与其他草地之间存在显著差异;温性荒漠草原物种丰富度为3.99个,除了与高寒草原无显著性差异之外,与其余草地之间均有显著差异;温性草原化荒漠,有3.02个物种,与其他草地数量均存在显著差异;温性荒漠和低平地草甸物种数分别为2.08和1.43个,两者物种数差异显著(图4c)。
2.3 不同植被类型物种丰富度与年平均降水之间的关系
新疆草地所有样地物种丰富度(S R)与年平均降水(M A P)呈显著线性关系(图5a),即降水量每增加100 m m,物种数相应的增加1.11个。进一步分析新疆不同草地类型S R与M A P之间的关系,低平地草甸和高寒草原,S R与M A P也呈显著线性关系(图5b,c),降水量每增加100 m m,其物种数分别增加1.19和1.58个。温性草原、温性荒漠草原和温性荒漠,S R与M A P均呈显著线性关系(图5f~h),即随着降水量每增加100 m m,物种丰富度分别增加0.71,0.94和0.58个。温性草甸草原、温性草原化荒漠S R与M A P均无显著相关性(图5d,e),说明年平均降水量的变化对这两个草地物种数的变化影响不大。
2.4 不同植被类型物种丰富度与年平均温度之间的关系
新疆草地总体物种丰富度(S R)与年平均温度(M A T)呈先增加再降低的趋势(图6a),当增加到3.79个物种时,达到最大值。低平地草甸和高寒草原的S R与M A T也呈显著线性关系(图6b,c),并且随着年平均温度每增加1℃,物种丰富度分别减少0.21和0.24个。温性草原和温性荒漠,S R与M A T呈先增加再降低的趋势,物种数最高分别达到5.60和2.24个(图6f,h)。而温性草甸草原、温性草原化荒漠和温性荒漠草原的S R与M A T均无显著相关性,说明温度变化对这3个草地的物种丰富度影响不大。
2.5 不同植被类型物种丰富度与海拔之间的关系
新疆草地所有样地物种丰富度(S R)与海拔呈显著峰型关系(图7a),即随着海拔的升高物种数增加到3.68个时,达到最大值。进一步分析新疆不同草地类型S R与海拔之间的关系,低平地草甸和高寒草原,S R与海拔呈先降低再增加的趋势(图7b,c),随着海拔的升高,物种丰富度分别减少到1.13和2.48个时,达到最小值。温性草原S R与海拔呈先增加再降低的趋势(图7f),当物种数达到5.67个时为最大值。温性荒漠S R与海拔呈线性关系(图7h),即随着海拔每增加1000 m,物种丰富度降低0.3个。温性草甸草原、温性草原化荒漠和温性荒漠草原S R与海拔均无显著相关性(图7d,e,g),说明海拔对此3种草地的物种丰富度影响不显著。
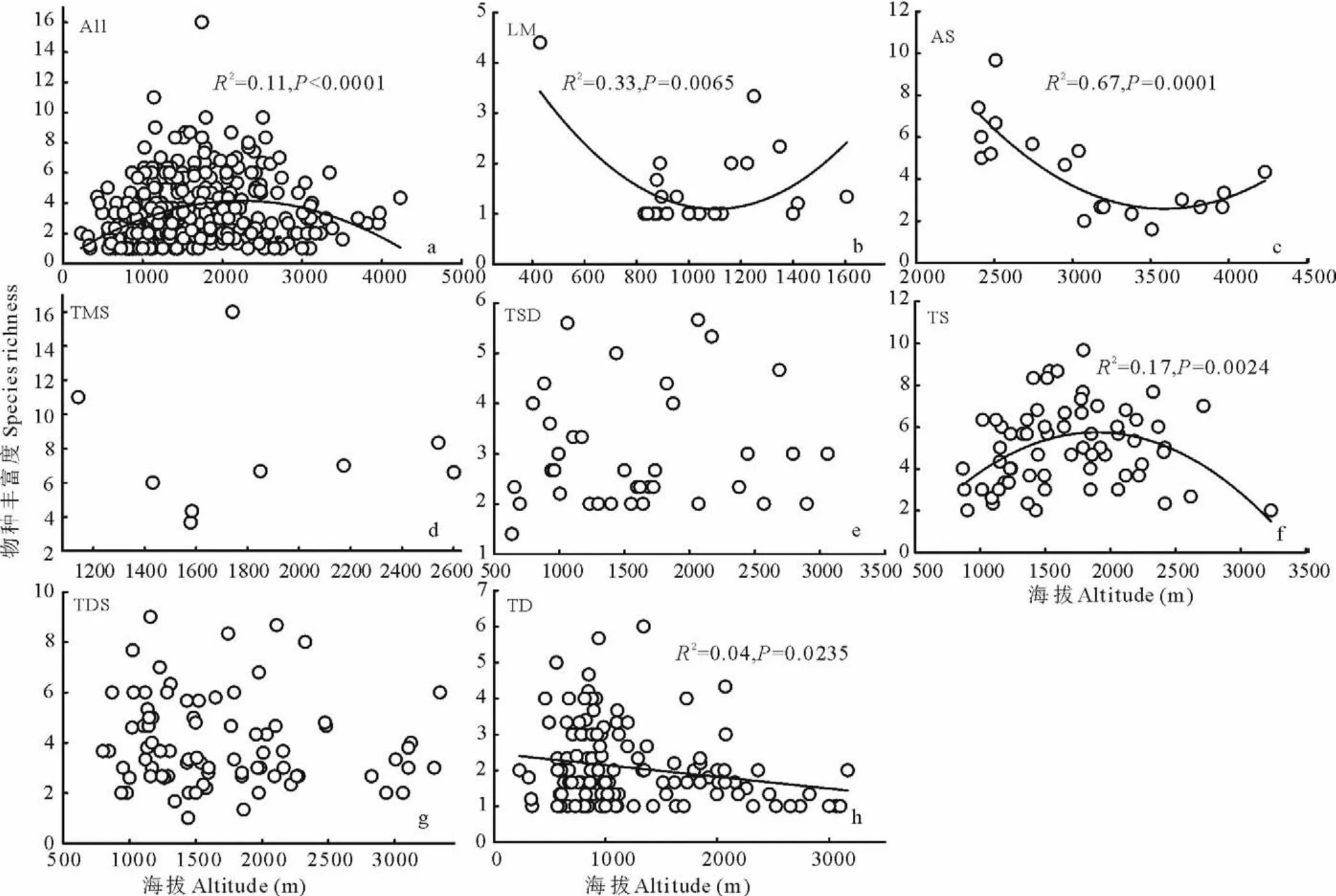
图7 不同草地类型物种丰富度与海拔之间的关系Fig.7 Relationships between species richness and altitude on different grasslands types
3 讨论
3.1 新疆草地的植物物种特征
通过调查发现,新疆草地中,温性草甸草原之间植物科、属、种均无显著差异,温性草甸草原有最高的物种数,温性荒漠和低平地草甸物种丰富度最低,这种研究结果与Bai等[28]在蒙古草原和M a等[29]在西藏草原的研究结果一致,但是他们的结果得出物种丰富度均为20个左右,荒漠类为10个左右,我们的结果与其有一定的差别,温性草甸草原物种为7.73个,温性荒漠为2.08个,而低平地草甸为1.43个,比较发现,新疆草地的物种丰富度明显低于内蒙古和西藏,造成这种结果的很大原因是由于水分和盐分的限制,相比于内蒙古和西藏,新疆的降水量较少,而蒸发量较大,从而使得盐分聚集于土壤表面,且含盐量很高。有研究表明,土壤盐分是引起干旱区植物群落多样性变化的关键因子之一[30],在高土壤盐分的胁迫条件下,可形成局域群落内的物种减少,且高胁迫条件下群落内物种间竞争加剧,可利用资源受限,种间从互利向竞争转变,物种趋于独立分布,造成多样性降低[31-32]。因此,温性荒漠和低平地草甸的物种丰富度很低,尤其是低平地草甸,土壤盐分多,物种单一,以芦苇属为主要植物类型。
3.2 温度与降水对物种丰富度的影响
草地的物种丰富度和物种多样性主要受环境因素控制,即温度、降水和有效性养分[33]等。我们的研究发现,新疆草地总体上来看,高的降水量主导高的物种多样性,与Bai等[28]、Chen等[34]及Li等[26]分别在内蒙古和新疆的研究结果相同,说明在干旱、半干旱区域,水分是限制物种多样性的主要原因,但是本文的结果降水对于物种多样性的解释只有13%,而随着降水量的增加土壤中的氮含量会有所增加,有研究认为,在干旱、半干旱区域,物种丰富度主要的限制因素是土壤氮[35],也有认为植物多样性的提高,使土壤中的限制性养分能够更完全地被利用,减少生态系统中N素的淋溶损失[36]。由此可见,对新疆草地物种多样性影响的另一种因素可能是土壤中的氮。对于温性草原化荒漠,受水分限制较大,但是S R与M A P并没有表现出显著的相关性,这一结果与代爽等[37]发现相同,其认为很多植物类群的物种丰富度与M A P之间没有表面的相关性,O’Brien[22]的研究表明,物种多样性很大程度是由水分决定的,但是植物所能获得的有效水分并不简单地决定于降水,还受到能量所决定的蒸散水平的强烈影响,充分解释了其不受降水影响的原因,支持水分-能量假说。
温度对于群落物种丰富度的影响是一个更直接的过程[38],其决定着水分的蒸散能力。我们的研究发现,新疆草地总体、温性草原、温性荒漠,物种丰富度随着M A T呈先增加再降低的趋势,这与Li等[26]对维管束植物的研究结果相同,而在全球尺度上,Kreft和Jetz[39]发现,在蒸散量为505 m m之前,被子植物丰富度随年最大可能蒸散量增加显著升高,之后便呈下降趋势。主要是由于随着温度的升高,促进植物的生长和碳同化作用[40],然而随着温度(能量)达到最适宜温度,水分蒸散加强,植物所能利用的有效水分减少,使植物受到干旱胁迫,导致多样性降低[41]。低平地草甸和高寒草原均表现出物种丰富度与M A T呈显著负相关,白永飞等[35]在内蒙古草原探讨了植物多样性沿水热梯度的变化,得出相同的结论。温性草甸草原、温性草原化荒漠和温性荒漠草原物种丰富度与温度之间无相关性,这又与代爽等[37]对草本植物的研究以及Chen等[34]在内蒙古草原的研究结果相同,由此可知不同的植被类型、不同的研究区域及方法,其所产生的结果有所差别。而不同草地类型对温度、降水的响应情况的差异性,也可能是由于研究单元的空间分布面积较大,单元内气候的空间分异强烈,而年平均温度和年平均降水量只能反映研究单元内总体的气候状况,不能较为全面地反映研究单元内气候的空间分异和季节分异[42]。
3.3 海拔对物种丰富度的影响
海拔是水热条件的综合因子,我们的研究发现新疆草地总体与温性草原,物种丰富度随着海拔的升高而先升高再降低,均是海拔达到2000 m左右时物种丰富度达到了最大值,这一发现与郭正刚等[24]在阿勒泰地区与Zhang等[43]在云南的研究结果相同,而徐远杰等[44]在伊犁河谷的研究同样发现其丰富度的第二个峰值也是出现在海拔2000 m左右,说明海拔在2000 m左右是物种多样性发生较大变化的一个重要转折点,同时也是物种多样性较丰富的地带。对于温性荒漠,物种丰富度与海拔呈负相关,W ang等[45]的研究也得出相同的结论,在低平地草甸和高寒草原,随着海拔升高,物种丰富度呈先降低再增加趋势,主要是由于在低海拔地区环境条件相对优越,竞争力强的物种会占优势,而随着海拔升高环境相对较恶劣,使这些竞争力强的物种也受到环境的威胁,物种降低[46],随着物种对环境的适应力增强,在中间海拔梯度时,水热条件比高海拔梯度优越,所以物种又出现上升的趋势。而温性草甸草原、温性草原化荒漠和温性荒漠草原S R与海拔均无显著相关性,O hle m uller和Wilson[47]在新西兰的探讨发现物种多样性与海拔之间并没有显著的相关性,对于各结果的分异来源于海拔梯度范围和所研究地区大尺度的气候特征[48]。
4 结论
本文通过对新疆不同草地类型物种的调查,统计不同草地物种的科属种构成,并分析物种丰富度对温度、降水和海拔的响应,得出以下初步结论:
1)新疆草地以禾本科、菊科为优势科,以针茅属、绢蒿属、苔草属为优势属;植物在科属种水平上数量的变化均为:温性草甸草原>温性草原>高寒草原>温性荒漠草原>温性草原化荒漠>温性荒漠>低平地草甸。
2)新疆草地物种丰富度相对较低的原因,主要归结于新疆的降水量少,蒸发量大,土壤盐分聚集于表层,对植物造成干旱胁迫和盐胁迫,使物种多样性较低。
3)新疆不同草地物种丰富度对年平均温度、降水的差异,主要原因是由于植被类型的差异以及气候因子的空间分异性,而其对于海拔的分异来源于海拔梯度范围和所研究地区大尺度的气候特征。
References:
[2]Shan Y,Chen D,Guan X,etal.Seasonally dependent im pacts of grazing on soil nitrogen mineralization and linkages to ecosyste m functioning in Inner M ongolia grassland.Soil Biology & Bioche mistry,2011,43(9):1943-1954.
[3]Lu H,Cong J,Liu X,etal.Plant diversity patterns along altitudinal gradients in alpine m eadows in the Three River H ead water Region,China.Acta Prataculturae Sinica,2015,24(7):197-204.
[4]Luo L M,Miao Y J,W u J S,etal.Variation in the biodiversity of m ontane shrub grassland co m m unities along an altitudinal gradient in a Lhasa River basin valley.Acta Prataculturae Sinica,2014,23(6):320-326.
[5]Arnesen G,Beck P S A,Engelskjn T.Soil acidity,content of carbonates,and available phosphorus are the soil factors best correlated with alpine vegetation:Evidence fro m Tro ms,North Norway.Arctic,A ntarctic,and Alpine Research,2007,39(2):189-199.
[6]Wu J,Shen Z,Zhang X.Precipitation and species co m position prim arily determine the diversity-productivity relationship of alpine grasslands on the Northern Tibetan Plateau.Alpine Botany,2014,124(1):13-25.
[7]Fry E L,M anning P,Allen D G P,etal.Plant functional group co m position m odifies the effects of precipitation change on grassland ecosyste m function.Plos O ne,2013,8(2):396-396.
[8]W ang Z H,Brow n J H,Tang Z Y,etal.Te m perature dependence,spatial scale,and tree species diversity in eastern Asia and North A m erica.Proceedings of the National Acade m y of Sciences,2009,106(32):13388-13392.
[9]Guo Z G,Cheng G D,W ang G X.Plant diversity of alpine Kobresia m eadow in the northern region of the Tibetan Plateau.Journal of Glaciology and Geocryology,2004,26(1):95-100.
[10]M elo A,Rangel T,Diniz J.Environ m ental drivers of beta-diversity patterns in New-W orld birds and m a m m als.Ecography,2009,32:226-236.
[11]Loreau M.Biodiversity and ecosyste m functioning:recent theoretical advances.Oikos,2000,91(1):3-17.
[12]Loreau M,Naee m S,Inchausti P,etal.Biodiversity and ecosyste m functioning:current knowledge and future challenges.Science,2001,294(5543):804-808.
[13]Baskin C C,Baskin J M.Seeds:Ecology,Biogeography,and Evolution of Dorm ancy and Germination[M].San Diego:Acade mic Press,2001.
[14]A dler P B,Hillerisla m bers J.The influence of clim ate and species co m position on the population dyna mics of ten prairie forbs.Ecology,2008,89(11):3049-3060.
[15]Alward R D,Detling J K,Milchunas D G.Grassland vegetation changes and nocturnal global warming.Science,1999,283 (5399):229-231.
[16]Engler R,Randin C F,Thuiller W,etal.21st century clim ate change threatens m ountain flora unequally across Europe.Global Change Biology,2011,17(7):2330-2341.
[18]K napp A K,Fay P A,Blair J M,etal.Rainfall variability,carbon cycling,and plant species diversity in a m esic grassland.Science,2002,298(5601):2202-2205.
[19]Fay P A,Blair J M,S mith M D,etal.Relative effects of precipitation variability and warming on grassland ecosyste m function.Biogeosciences Discussions,2011,8:6859-6900.
[20]Zhang L,W ylie B K,Ji L,etal.U pscaling carbon fluxes over the Great Plains grasslands:Sinks and sources.Journal of Geophysical Research:Biogeosciences(2005-2012),2011,116(1):622-635.
[21]W ang J B,Zhang D G,Cao G M,etal.Regional characteristics of the alpine meadow degradation succession on the Qinghai-Tibetan Plateau.Acta Prataculturae Sinica,2013,22(2):1-10.
[22]O’Brien E M.Biological relativity to water-energy dyna mics.Journal of Biogeography,2006,33(11):1868-1888.
[23]Feng Y,Zhang Y M,Pan B R.Grassland types with environ m ental relations in middle zone of northern slop of Tianshan M ountains.Arid Land Geography,2006,29(2):237-242.
[24]Guo Z G,Liang T G,Liu X Y,etal.Species diversity of grassland co m m unities in the Aletairegion of the northern Xinjiang province.Acta Botanica Boreali-occidentalia Sinica,2003,23(10):1719-1724.
[25]Li L P,W ang Z,Zerbe S,etal.Species richness patterns and water-energy dyna mics in the drylands of North west China.PloS O N E,2013,8(6):e66450.
[26]Li L P,A bdusalih N,W ang S P,etal.Distribution patterns and clim atic explanations of species richness of vascular plants in Xinjiang,China.Arid Zone Research,2011,28(1):25-30.
[27]Xu P.Grassland Resources and Use in Xinjiang[M].Uru mchi:H ealth Science and Technology Publishing H ouse in Xinjiang,1993:202-204.
[28]Bai Y F,W u J G,Pan Q M,etal.Positive linear relationship between productivity and diversity:evidence fro m the Eurasian steppe.Journal of A pplied Ecology,2007,44:1023-1034.
[29]M a W,H e J S,Yang Y,etal.Environ m ental factors covary with plant diversity-productivity relationships a m ong Chinese grassland sites.Global Ecology and Biogeography,2010,19(2):233-243.
[30]W ang S X,Dong X G,Liu L.Natural vegetation diversity differentiation under water and salt gradients in the Oasis of Yanqi Basin,Xinjiang.Journal of Glaciology and Geocryology,2010,(3):999-1006.
[31]M aestre F T,Cortina J.Do positive interactions increase with abiotic stress?A test fro m a se mi-arid steppe.Proceedings of the Royal Society of London B:Biological Sciences,2004,271(Suppl 5):331-333.
[32]Cavieres L A,Badano E I.Do facilitative interactions increase species richness at the entire co m m unity level.Journal of Ecology,2009,97(6):1181-1191.
[33]W u J S,Zhang X,Shen Z,etal.Species richness and diversity of alpine grasslands on the Northern Tibetan Plateau:Effects of grazing exclusion and growing season precipitation.Journal of Resources and Ecology,2012,3(3):236-242.
[34]Chen L,Li H,Zhang P,etal.Clim ate and native grassland vegetation as drivers of the co m m unity structures of shrub-encroached grasslands in Inner M ongolia,China.Landscape Ecology,2014,30:1-15.
[35]Bai Y F,Li L H,W ang Q B,etal.Changes in plant species diversity and productivity along gradients of precipitation and elevation in the Xilin River Basin,Inner M ongolia.Acta Phytoecologica Sinica,2000,24:667-673.
[36]Tilm an D,W edin D,K nops J.Productivity and sustainability influenced by biodiversity in grassland ecosyste ms.Nature,1996,379:718-720.
[37]Dai S,W ang X P,Liu C,etal.Relationship between shrub species richness and clim ate across central Inner M ongolia,China.Acta Scientiaru m Naturaliu m U niversitatis Pekinensis,2013,49(4):689-698.
[38]Vonlanthen C M,Ka m m er P M,Eugster W,etal.Alpine vascular plant species richness:the im portance of daily m axim u m te m perature and p H.Plant Ecology,2006,184(1):13-25.
[39]Kreft H,Jetz W.Global patterns and determinants of vascular plant diversity.Proceedings of the National Acade m y of Sciences,2007,104(14):5925-5930.
[40]W alther G R.Plants in a warm er world.Perspectives in Plant Ecology,Evolution and Syste m atics,2003,6(3):169-185.
[41]M cCain C M,Sanders N J.M etabolic theory and elevational diversity of vertebrate ectotherms.Ecology,2010,91(2):601-609.
[42]Feng J M.Spatial patterns of species diversity of seed plants in China and their clim atic explanation.Biodiversity Science,2008,16(5):470-476.
[43]Zhang S B,Chen W Y,H uang J L,etal.Orchid species richness along elevational and environ m ental gradients in Y unnan,China.PLoS O N E,2015,10(11):e0142621.
[44]Xu Y J,Chen Y N,Li W H,etal.Distribution pattern and environ m ental interpretation of plant species diversity in the m ountainous region of Ili River Valley,Xinjiang,China.Chinese Journal of Plant Ecology,2010,34(10):1142-1154.
[45]W ang W,W ang Q,Li S,etal.Distribution and species diversity of plant co m m unities along transect on the Northeastern Tibetan plateau.Biodiversity & Conservation,2006,15(5):1811-1828.
[46]Liu Z,Li Q,Chen D D,etal.Patterns of plant species diversity along an altitudinal gradient and its effect on above-ground bio m ass in alpine m eadows in Qinghai-Tibet Plateau.Biodiversity Science,2015,23(4):451-462.
[47]O hle m uller R,Wilson J B.Vascular plant species richness along latitudinal and altitudinal gradients:a contribution fro m New Zealand te m perate rainforests.Ecology Letters,2000,3(4):262-266.
[48]Nogués-Bravo D,Araújo M B,Ro m dal T,etal.Scale effects and hu m an im pact on the elevational species richness gradients.Nature,2008,453(7192):216-219.
[3]卢慧,丛静,刘晓,等.三江源区高寒草甸植物多样性的海拔分布格局.草业学报,2015,24(7):197-204.
[4]罗黎鸣,苗彦军,武建双,等.拉萨河谷山地灌丛草地物种多样性随海拔升高的变化特征.草业学报,2014,23(6):320-326.
[9]郭正刚,程国栋,王根绪.青藏高原北部高海拔地区嵩草草甸植物多样性分析.冰川冻土,2004,26(1):95-100.
[21]王建兵,张德罡,曹广民,等.青藏高原高寒草甸退化演替的分区特征.草业学报,2013,22(2):1-10.
[23]冯缨,张元明,潘伯荣.天山北坡中段草地类型的生态梯度组合格局与环境分析研究.干旱区地理,2006,29(2):237-242.
[24]郭正刚,梁天刚,刘兴元,等.新疆阿勒泰地区草地类型及植物多样性的研究.西北植物学报,2003,23(10):1719-1724.
[26]李利平,努尔巴依·阿布都沙力克,王少鹏,等.新疆野生维管束植物物种丰富度分布格局的水热解释.干旱区研究,2011,28(1):25-30.
[27]许鹏.新疆草地资源及其利用[M].乌鲁木齐:新疆科技卫生出版社,1993:202-204.
[30]王水献,董新光,刘磊.新疆焉耆盆地绿洲水盐双梯度下天然植被多样性分异特征.冰川冻土,2010,(5):999-1006.
[35]白永飞,李凌浩,王其兵,等.锡林河流域草原群落植物多样性和初级生产力沿水热梯度变化的样带研究.植物生态学报,2000,(6):667-225.
[37]代爽,王襄平,刘超,等.内蒙古灌木群落物种丰富度与气候的关系.北京大学学报:自然科学版,2013,49(4):689-698.
[42]冯建孟.中国种子植物物种多样性的大尺度分布格局及其气候解释.生物多样性,2008,16(5):470-476.
[44]徐远杰,陈亚宁,李卫红,等.伊犁河谷山地植物群落物种多样性分布格局及环境解释.植物生态学报,2010,34(10):1142-1154.
[46]刘哲,李奇,陈懂懂,等.青藏高原高寒草甸物种多样性的海拔梯度分布格局及对地上生物量的影响.生物多样性,2015,23(4):451-462.
Relationship between plant species characteristics and climate factors in different grassland types of Xinjiang
For 364 field sites representing seven grassland types in Xinjiang,species lists were co m piled,and the distribution of plant fa milies,genera and species across the different grassland types cataloged.In addition,we exa mined the species richness(S R)response to m ean annual precipitation(M A P),m ean annual te m perature(M A T)and altitude.T he Poaceae and Ateraceae were the do minant fa milies,and Stipa,and Seriphidium were a m ong the do minant genera at all sites.T he grassland types ranked for S R(fro m high to low):te mperate m eadow steppe(7.73),te m perate steppe(5.04),alpine steppe(4.36),te m perate desert steppe (3.99),te m perate steppe desert(3.02),te m perate desert(2.08),low plain m eadow(1.43).Patterns of S R response to M A P,M A T and altitude were also detected.In Xinjiang grassland te m perate steppe,S R was positively and linearly related to M A P,but unim odally related to M A T and altitude.In alpine steppe and low plainm eadow,S R was positively and linearly related with M A P and negatively related with M A T,but S R decreased fro m low to mid altitude and then increased with increasing altitude.In te m perate desert steppe,S R was significantly positively related to M A P,but was not significantly related to M A T and altitude.In alpine steppe and low plain m eadow,the S R was positively and linearly related to M A P and negatively related to M A T,but unim odally related to altitude.N o significant relationship was found between S R and M A T,M A P,or altitude in te m perate m eadow steppe and te m perate steppe desert grasslands.T he above results in the different grassland types might be explained by response characteristics of particular species,spatial variation of te m perature and precipitation,and the altitude range,steepness of slope and large scale clim ate characteristics of the study area.
altitude;grassland;m ean annual precipitation;m ean annual te m perature;species richness
.E-m ail:sjd_2004@126.com
10.11686/cyxb2015517
2015-11-17;改回日期:2016-01-04
中国科学院战略性先导科技专项(X D A05050400)和新疆维吾尔自治区土壤学重点学科资助。
刘利利(1990-),女,新疆巴州人,在读硕士。E-m ail:455523008@qq.com
猜你喜欢
成都信息工程大学学报(2022年4期)2022-11-18
科学24小时(2022年10期)2022-10-11
小哥白尼(趣味科学)(2022年5期)2022-08-15
青海草业(2022年2期)2022-07-23
安徽师范大学学报(2020年6期)2020-12-18
绿色中国(2019年14期)2019-11-26
活力(2019年21期)2019-04-01
儿童故事画报·智力大王(2016年7期)2017-02-08
文史春秋(2016年3期)2016-12-01
河北林业科技(2016年5期)2016-11-08
