
国审品种‘苏植1号’杂交结缕草抗旱性初步评价与分析
2016-07-05 08:02汪毅郭海林陈静波宗俊勤李丹丹姜亦巍刘建秀江苏省中国科学院植物研究所江苏南京210014
草业学报 2016年5期
汪毅,郭海林,陈静波,宗俊勤,李丹丹,姜亦巍,刘建秀(江苏省中国科学院植物研究所,江苏 南京 210014)
W A N G Yi,G U O H ai-Lin,C H E N Jing-Bo,Z O N G Jun-Qin,LI Dan-Dan,JIA N G Yi-W ei,LIU Jian-Xiu*Instituteof Botany,Jiangsu Province and Chinese Academy of Sciences,Nanjing 210014,China
国审品种‘苏植1号’杂交结缕草抗旱性初步评价与分析
汪毅,郭海林,陈静波,宗俊勤,李丹丹,姜亦巍,刘建秀*
(江苏省中国科学院植物研究所,江苏 南京 210014)
以‘苏植1号’杂交结缕草为研究对象,‘兰引3号’结缕草为对照品种,采用沙培试验控水处理,根据叶片萎蔫度、叶片相对含水量、根系电解质外渗率和土壤含水量4个指标对‘苏植1号’的抗旱性进行了初步评价,同时分析了两个品种的根系生长特性(根系干重、比根长)和干旱胁迫下最大光化学量子产量(Fv/Fm)的变化特点。结果表明,干旱胁迫过程中,两个品种的叶片萎蔫度逐渐增大,土壤含水量逐渐下降,不同品种的变化趋势相似,胁迫结束时,两个品种的叶片萎蔫度、叶片相对含水量、根系电解质外渗率和土壤含水量均无显著差异,说明‘苏植1号’整体抗旱表现与‘兰引3号’近似。同时,相对于‘兰引3号’而言,虽然‘苏植1号’的根系干重显著较小,但是较大的比根长和干旱胁迫中期较高的Fv/Fm值很可能使‘苏植1号’在一定程度上抵消了较小的根系生物量所带来的不利影响,是其重要的抗旱机制。总体来看,‘苏植1号’与‘兰引3号’抗旱性相当,但两者具有不同的抗旱途径。
‘苏植1号’杂交结缕草;抗旱性;评价;根系生长特性;最大光化学量子产量
http://cyxb.lzu.edu.cn
汪毅,郭海林,陈静波,宗俊勤,李丹丹,姜亦巍,刘建秀.国审品种‘苏植1号’杂交结缕草抗旱性初步评价与分析.草业学报,2016,25(5):30-39.WANG Yi,GUO H ai-Lin,CHEN Jing-Bo,ZONG Jun-Qin,LI Dan-Dan,JIA N G Yi-W ei,LIU Jian-Xiu.Preliminary evaluation of drought resistance for the hybrid zoysiagrass‘Suzhi N o.1’.Acta Prataculturae Sinica,2016,25(5):30-39.
W A N G Yi,G U O H ai-Lin,C H E N Jing-Bo,Z O N G Jun-Qin,LI Dan-Dan,JIA N G Yi-W ei,LIU Jian-Xiu*Instituteof Botany,Jiangsu Province and Chinese Academy of Sciences,Nanjing 210014,China
虽然我国水资源总量位居世界第六,但同样遭受干旱缺水的困扰,并面临水资源开发过度的问题[1]。草坪草在维护和修复生态环境等方面能够发挥重要作用,然而在水资源短缺时,通常会首先保证粮食作物的灌溉需求以及人们的生活用水,用于草坪建植等环境景观方面的供水会受到限制,因此培育抗旱性强、需水量少的草坪草新品种是一个迫切的任务,同时种质资源的抗旱性评价及相关机理分析是草坪草抗旱性研究的重点内容之一。
在暖季型草坪草中,结缕草属(Zoysia)的一些种类比如结缕草(或称日本结缕草)(Z.japonica)、中华结缕草(Z.sinica)、沟叶结缕草(Z.matrella)、细叶结缕草(Z.tenuifolia)以及部分杂交品种已经得到广泛应用,与其他草坪草一样,结缕草属植物的抗旱性研究及种质资源创新也受到较多关注。近二十年来,有关结缕草属植物的抗旱性研究,内容包括形态解剖、生长特性、种群特征、水分关系、渗透调节、抗氧化代谢等方面[2-23]。其中,对于结缕草属种质资源的抗旱性评价多有涉及:Carrow[3]根据焦叶率和萎蔫率对包括结缕草在内的7种草坪草的整体抗旱性作了排序,认为结缕草抗旱性最弱;卢少云等[8]测定了3种草坪草在土壤干旱条件下相对含水量和相对电导率的变化,发现矮生狗牙根(Cynodon dactylon×C.transvaalensis)的耐旱性较强,沟叶结缕草次之,而地毯草(Axonopuscom pressus)对干旱敏感;胡化广等[11]采用表征永久萎蔫系数对48份结缕草属种源进行了抗旱性评价,认为属内不同种的平均抗旱能力排名为细叶结缕草=沟叶结缕草>结缕草>大穗结缕草(Z.macrostachya)>中华结缕草;李志东等[13]采用株体萎蔫生物学鉴定法结合生化测定,将包括结缕草和细叶结缕草在内的7个暖季型草坪草种按抗旱性强弱显著度和灌溉周期长短顺序划分为3个级别;余晓华等[14]测定了土壤逐步干旱下4种结缕草属草坪草叶片相对含水量、质膜相对透性及脯氨酸含量的变化,认为4种草坪草抗旱能力强弱的顺序为‘兰引3号’结缕草(Z.japonica‘Lanyin N o.3’)>细叶结缕草>沟叶结缕草>日本结缕草;赵艳等[15]通过对草坪目测质量、叶绿素含量、光合速率、叶片电导率和丙二醛等指标的综合分析,评价了狗牙根属和结缕草属6个栽培品种的抗旱性能;吕静等[17]在土壤持续干旱过程中,通过测定水分饱和亏、质膜透性、叶绿素含量、脯氨酸含量、丙二醛含量等生理生化指标,运用模糊隶属函数法对4种暖季型草坪草进行了抗旱性评价,结果表明抗旱性强弱为狗牙根(C.dactylon)>结缕草>百喜草(Paspalum notatum)>假俭草(Eremochloa ophiuroides);俞乐等[18]通过分析比较干旱胁迫下叶片抗坏血酸含量与相关生理指标(相对含水量、相对电导率、M D A含量、P O D活性、叶绿素含量、Rubisco含量)的品种差异,认为日本结缕草抗旱能力弱于细叶结缕草;谢新春等[21]采用萎蔫及恢复时间、叶片相对含水量、叶片相对电导率、脯氨酸含量和可溶性总糖含量评价了‘兰引3号’及两个结缕草新品系的抗旱性强弱;宋华伟等[23]根据叶片相对含水量、相对电导率、脯氨酸含量、可溶性糖含量以及草坪草盖度和颜色变化,对3种结缕草及其辐射诱变选育的6种新品系进行了抗旱性综合评价。
可用于草坪草抗旱性评价的指标很多,涉及外观表现(如叶片萎蔫度、草坪质量)、生长特性(如茎叶和根系生物量)、水分状况(如叶片相对含水量、水分饱和亏、土壤含水量)、受损程度(如电解质外渗率、丙二醛含量)、光合特性(如叶绿素含量、光合速率、叶绿素荧光、光合关键酶活性)和内在调节机制(如抗氧化物质活性、渗透调节物质含量)等,这些指标反映了草坪草在干旱胁迫下的形态和生理生化等各个方面的变化规律,并且一些指标之间具有显著的相关性,在特定的研究中,通常根据具体的研究目的和试验条件而采用一至多项指标对不同草坪草种的抗旱性作出评价与分析[11,15,17,24-29]。在众多评价指标中,叶片萎蔫度(率)能够从外观上直接体现草坪草的受旱程度,叶片相对含水量是草坪草体内水分状况的良好指针,而根系电解质外渗率和土壤含水量则表明了草坪草根系的损伤程度和吸水能力,上述4个指标可以反映草坪草在干旱胁迫下的外观与内在、地上部与地下部的响应特点,并且操作简便,通过综合分析,能够获得较为可靠的评价结果。
‘兰引3号’结缕草是目前我国生产上的结缕草主栽品种之一,质地好、生长快,在与多种结缕草属植物的对比中,‘兰引3号’表现出较强的抗旱性[14-15,21,30]。‘苏植1号’杂交结缕草(Z.japonica×Z.tenuifolia‘Suzhi N o.1’)是由江苏省中国科学院植物研究所自主选育的杂交结缕草新品种,于2010年4月通过农业部草品种审定,与生产上应用的其他结缕草品种相比,该品种具有绿期长、生长速度快、质地柔软、耐践踏等优良特征,可用于长江三角洲及以南地区的观赏草坪、公共绿地、运动场草坪以及保土草坪建植。‘苏植1号’草坪外观较为细致(叶宽3.65 m m,每100 cm2的直立枝数目为243.3),正常水分管理条件下,具有较好的坪用质量,但是尚未有相关研究对该品种的抗旱性进行过评价与分析。此外,草坪草的抗旱性作为数量性状,不仅存在多样的抗旱表现,其抗旱机制及途径也不尽相同[5],迄今为止,在结缕草属优良品种的应用和推广当中,有关不同的抗旱途径如何影响抗旱表现的研究尚不够深入。
本试验以‘苏植1号’杂交结缕草为研究对象,‘兰引3号’结缕草为对照品种,通过对比干旱胁迫下两个品种的叶片萎蔫度、叶片相对含水量、根系电解质外渗率和土壤含水量4个指标的差异,初步评价了‘苏植1号’杂交结缕草的抗旱性,同时对两个品种在正常供水以及干旱胁迫下的根系生长特性(根系干重和比根长)和最大光化学量子产量(Fv/Fm)的变化特点进行分析,从而探讨其相关抗旱机制及其途径,为品种应用推广中的水分管理提供技术支撑,也为结缕草属植物品种抗旱性的遗传改良以及在干旱条件下改进栽培管理措施提供试验依据。
1 材料与方法
1.1 试验材料
以两个结缕草属植物品种‘苏植1号’和‘兰引3号’为试验材料,所用草茎均取自江苏省中国科学院植物研究所草坪草种质资源圃(北纬32°02′,东经118°28′,海拔55 m)。
1.2 试验设计
2014年6月上旬,在资源圃中挖取生长均匀的草皮,洗净沙土,地上部分留茬高度5 c m,剪去根系,将草皮剪成直径10 cm 的圆形,并栽植于装满河沙的P V C管(内直径10 cm ,高60 cm ,管底部附有带孔的盖子以用于排水)中,之后在露天进行养护管理,每周浇水3次、浇施1/2 H oagland营养液1次、修剪(管口以上草层高度达到5 c m后开始修剪,并保持该高度)1次。待地上部草层和地下部根系建成以后,于2014年8月中旬移入塑料大棚适应生长约3周(在此期间养护管理措施与露天培养阶段相同),2014年9月10日开始试验处理,10月19日结束处理。处理期间,环境最低和最高温度平均值分别为18℃和32℃,最低和最高空气湿度平均值分别为35%和80%,天气晴好时大棚内正午的光合有效辐射约为600μm ol/(m2·s)。设置对照(正常浇水,以不出现萎蔫为标准)、干旱胁迫(停止浇水)两个处理,设3次重复。干旱胁迫开始前一天(9月9日)傍晚对所有处理充分浇水(P V C管底有水开始渗出为止),并进行最后一次修剪,以此作为干旱胁迫第0天,分别在干旱胁迫第1,12,21,29,40天观测叶片萎蔫度,第2,9,15,22,29,39天测定最大光化学量子产量,第1,13,25,40天测定土壤含水量,第40天测定叶片相对含水量、根系电解质外渗率、根系干重和比根长。
1.3 指标测定
1.3.1 叶片萎蔫度(leaf wilting,L W) 观测时间为干旱胁迫第1,12,21,29,40天上午8:00,参照同类研究[26,31-32]的方法并适当调整,采用目测法根据萎蔫叶片百分比按0~3分计分,0.5分为一级,共7级,0分代表不萎蔫(目测萎蔫比率为0%),1分代表轻度萎蔫(目测萎蔫比率为25%),2分代表中度萎蔫(目测萎蔫比率为55%),3分代表重度萎蔫(目测萎蔫比率为85%及以上),0.5分、1.5分、2.5分的目测萎蔫比率分别介于0~25%,25%~55%和55%~85%之间。
1.3.2 叶片相对含水量(relative water content,R W C) 取样时间为干旱胁迫第40天,参照Chai等[33]的方法,剪取新鲜叶片0.15 g左右,立即称取叶片鲜重(F W),随后将叶片在20℃水中浸泡4 h后称取叶片饱和重(T W),再将叶片在80℃下烘干72 h后称取叶片干重(D W),按公式R W C(%)=100×(F W-D W)/(T W-D W)计算叶片相对含水量。
1.3.3 根系电解质外渗率(root electrolyte leakage,R E L) 取样时间为干旱胁迫第40天,参照Sun等[34]的方法,剪去所有地上部分后,将地下部分(包括地下茎和根系)从P V C管中倒出,用水清洗沙土,称取0.2 g左右洗净根系,浸入去离子水中振荡24 h后测定溶液初始电导率(Ci),然后在高压蒸汽灭菌锅中120℃加热15 min,再行振荡24 h后测定溶液最终电导率(Cmax),按公式R E L(%)=100×Ci/Cmax计算根系电解质外渗率。
1.3.4 土壤含水量(soil water content,S W C) 测定时间为干旱胁迫第1,13,25,40天上午10:00,采用土壤水分测量仪(T ZS型,浙江托普仪器有限公司),将探针插入栽培基质(河沙)表层以下8 c m处进行测定。
1.3.5 比根长(specific root length,S R L) 采用根系扫描仪(Perfection V700,E PS O N)扫描全部根系,以Win R HIZ O Pro 2012b软件分析,获得根系长度,根系长度与根系干重之比即为比根长。
1.3.6 根系干重(root dry weight,R D W) 将全部根系在80℃下烘干72 h后称取根系干重。
1.3.7 最大光化学量子产量(Fv/Fm) 测定时间为干旱胁迫第2,9,15,22,29,39天下午14:30-16:00,采用便携式叶绿素荧光仪(O S1p,O P TI-Sciences),用叶夹夹住5~7枚叶片,每管材料测3个重复,暗适应30 min后测定。
1.4 数据分析
采用SPSS 19.0软件进行方差分析,Excel 2010和W O R D 2010软件进行图表绘制。
2 结果与分析
2.1 干旱胁迫对叶片萎蔫度的影响
如图1所示,干旱胁迫第12天,‘苏植1号’和‘兰引3号’都开始出现萎蔫,胁迫第29天,两个品种均达到中度萎蔫,至胁迫第40天,两个品种的叶片萎蔫度分别为2.7和2.5,均接近重度萎蔫,整个处理期间,除胁迫第21天外,在胁迫第1,12,29,40天,品种间的叶片萎蔫度均没有显著差异,表明在本次试验条件下,干旱胁迫对两个品种的叶片外观具有近似的影响。
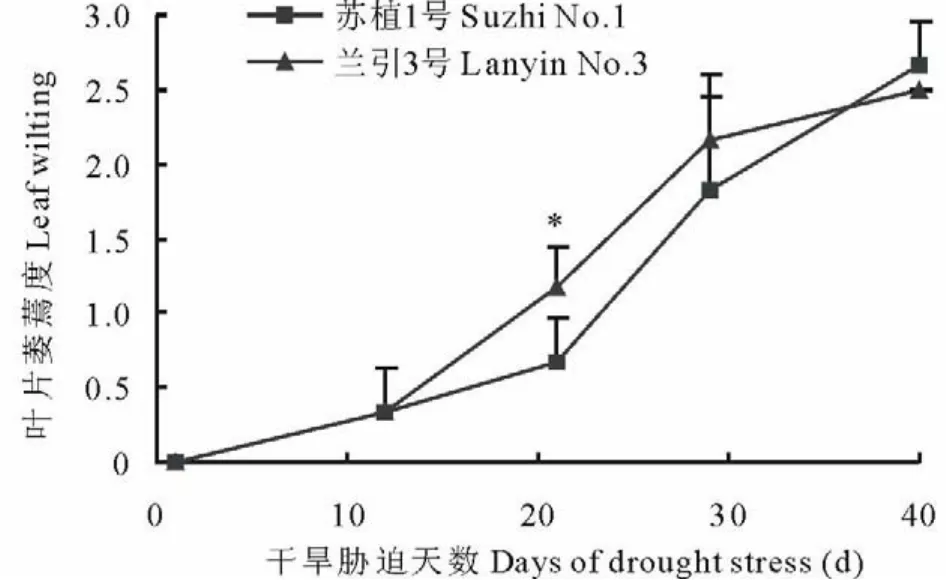
图1 干旱胁迫过程中两个品种的叶片萎蔫度Fig.1 Leaf wilting of two cultivars during drought stress
2.2 干旱胁迫对叶片相对含水量的影响
如图2所示,干旱胁迫第40天,‘苏植1号’和‘兰引3号’的叶片相对含水量均显著下降,分别为各自对照的87.2%和80.0%,‘苏植1号’下降幅度略小于‘兰引3号’,但品种间的差异未达显著水平,说明干旱胁迫结束时两个品种具有近似的内部水分状况。
2.3 干旱胁迫对根系电解质外渗率的影响
如图3所示,干旱胁迫第40天,‘苏植1号’和‘兰引3号’的根系电解质外渗率均显著上升,分别为各自对照的144.2%和132.0%,‘苏植1号’上升幅度略大于‘兰引3号’,但品种间的差异未达显著水平,表明干旱胁迫结束时两个品种根系质膜的损伤程度相近。
2.4 干旱胁迫对土壤含水量的影响
如图4所示,随干旱胁迫程度加重,‘苏植1号’和‘兰引3号’的土壤含水量呈逐渐下降趋势,至胁迫第13天,两个品种均显著低于各自对照,同时‘苏植1号’明显高于‘兰引3号’,至胁迫第25,40天,品种间不再具有显著差异,胁迫第40天时,‘苏植1号’和‘兰引3号’的土壤含水量分别为各自对照的44.2%和40.5%,以上情况表明,在处理前期,‘苏植1号’土壤含水量的下降程度低于‘兰引3号’,至处理中后期,两个品种的下降程度相同,但就整个干旱胁迫过程而言,两个品种的土壤含水量总体上具有类似的变化趋势。
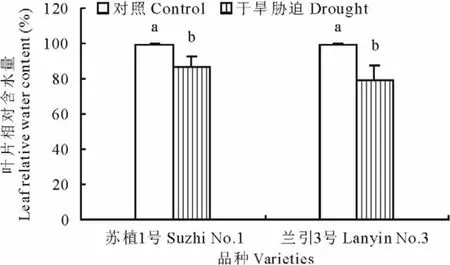
图2 干旱胁迫第40天两个品种的叶片相对含水量Fig.2 Leaf relative water content of two cultivars on day 40 of drought stress
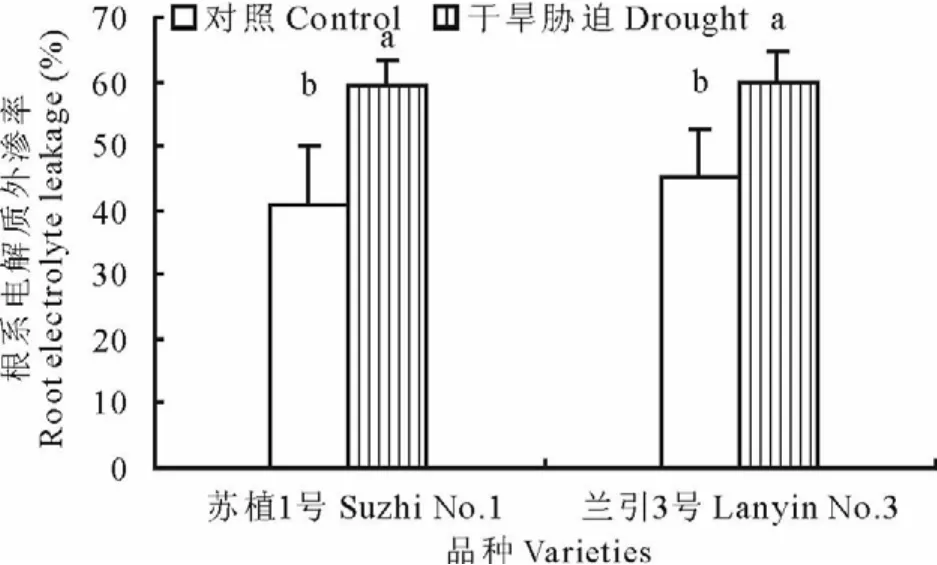
图3 干旱胁迫第40天两个品种的根系电解质外渗率Fig.3 Root electrolyte leakage of two cultivars on day 40 of drought stress
2.5 干旱胁迫对根系生长特性的影响
2.5.1 根系干重 如图5所示,在对照条件下,‘苏植1号’的根系干重显著低于‘兰引3号’,说明两个品种间本身存在遗传差异;同时在干旱胁迫第40天,‘苏植1号’和‘兰引3号’的根系干重分别为对照的101.0%和84.3%,‘苏植1号’与对照无显著差异,而‘兰引3号’则显著低于对照,表明干旱胁迫对‘苏植1号’的根系生物量没有影响,却使‘兰引3号’的根系生物量明显下降。尽管如此,‘兰引3号’在干旱胁迫下仍然具有显著大于‘苏植1号’的根系生物量。
2.5.2 比根长 如图6所示,在对照条件下,‘苏植1号’的比根长显著高于‘兰引3号’,这是由于两个品种间本身存在遗传差异;干旱胁迫第40天,‘苏植1号’和‘兰引3号’的比根长分别为各自对照的102.1% 和89.2%,两个品种与各自对照的差异均不显著,表明干旱胁迫对‘苏植1号’和‘兰引3号’各自的比根长没有明显的影响。
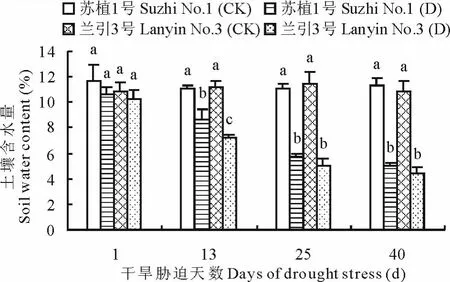
图4 干旱胁迫过程中两个品种的土壤含水量Fig.4 Soil water content of two cultivars during drought stress
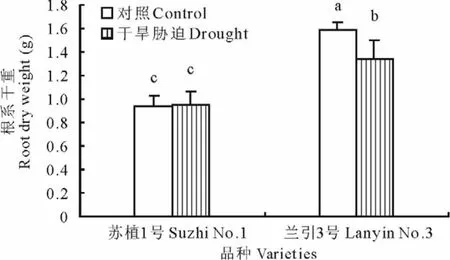
图5 干旱胁迫第40天两个品种的根系干重Fig.5 Root dry weight of two cultivars on day 40 of drought stress
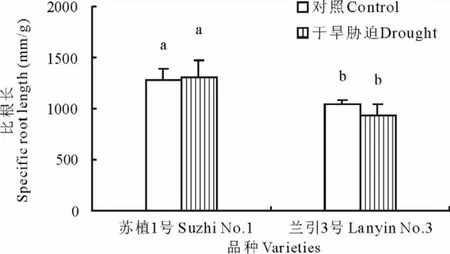
图6 干旱胁迫第40天两个品种的比根长Fig.6 Specific root length of two cultivars on day 40 of drought stress
2.6 干旱胁迫对最大光化学量子产量(Fv/Fm)的影响
如图7所示,干旱胁迫下随着胁迫天数增加,‘苏植1号’和‘兰引3号’的最大光化学量子产量(Fv/ Fm)呈逐渐下降趋势,虽然在干旱胁迫第2天,‘苏植1号’低于‘兰引3号’,但是在干旱胁迫中期(胁迫第9,15,22,29天),‘苏植1号’却持续高于‘兰引3号’,品种间的差异达到显著水平,至胁迫第29天,‘兰引3号’的Fv/Fm值开始显著低于其对照,而‘苏植1号’此时仍维持在对照水平,直至胁迫第39天,两个品种的Fv/Fm值均显著低于各自对照,‘苏植1号’和‘兰引3号’分别为各自对照的92.4%和92.3%,品种间不再具有显著差异,以上情况表明,在干旱胁迫中期,‘苏植1号’能够维持比‘兰引3号’更高的PSⅡ光能转换效率。
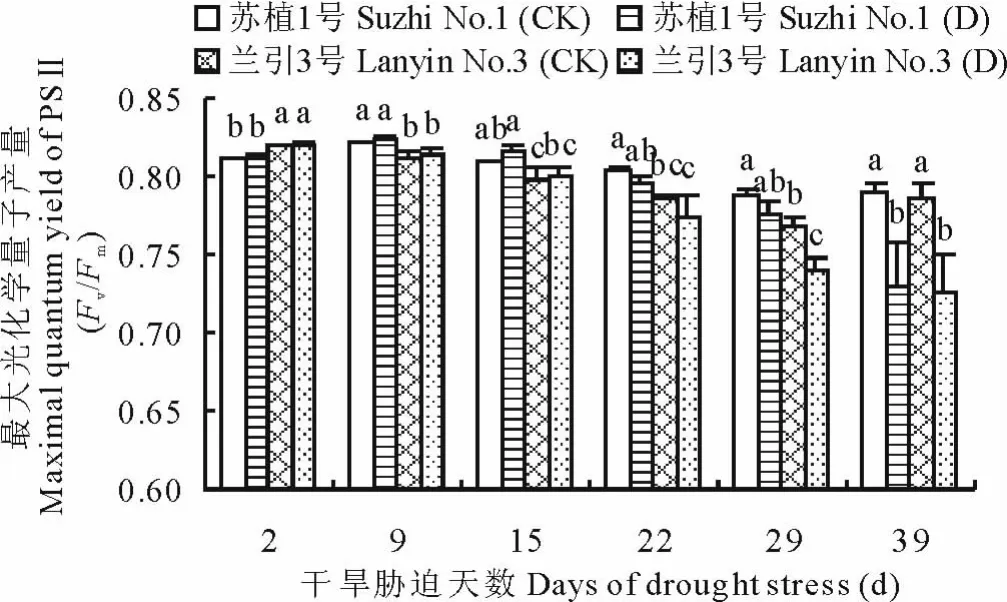
图7 干旱胁迫过程中两个品种的最大光化学量子产量(Fv/Fm)Fig.7 M aximal quantu m yield of PSⅡ(Fv/Fm)of two cultivars during drought stress
3 结论与讨论
干旱胁迫下,草坪草会出现各种形态指标和生理指标的变化,通过对这些指标进行综合分析,可以鉴定和评价不同草坪草种的抗旱性。本试验中,以叶片萎蔫度、叶片相对含水量、根系电解质外渗率和土壤含水量这4个指标对‘苏植1号’杂交结缕草的抗旱性进行了初步评价。叶片萎蔫度(率)被用来评估草坪草受旱后叶片的失水程度,是植株损伤和草坪质量下降的直观反映,能够衡量草坪草抗旱性的强弱[26,31]。叶片相对含水量是表征草坪草保水力的一个重要指标,反映了植株内部的水分状况,干旱胁迫下具有较高的叶片相对含水量通常表明具有较强的抗旱性[18,24,26,28,33]。干旱胁迫还造成草坪草膜系统损伤,使质膜透性增大,表现在地上部叶片或者地下部根系的电解质外渗率升高,由于根系质膜的损伤会直接影响根系活力,并且根系直接与干旱土壤接触,因此根系电解质外渗率比叶片电解质外渗率更好地反映了草坪草的耐脱水能力,干旱胁迫下能够保持相对较低的根系电解质外渗率是抗旱性较强的表现[15,34-35]。此外,土壤含水量可被用于指示土层中根系的垂直分布情况,能够反映不同土层中的根系活动力或者说根系的吸水能力,在尚有水分供给时,一定土层中较低的土壤含水量往往表明分布于该土层的根系具有较强的活动能力[4]。与一些生理生化指标(如丙二醛含量、抗氧化酶活性等)相比,上述4个指标能够反映草坪草植株在干旱胁迫下的外观与内在、地上部与地下部的响应特点,并且操作简便,测定时干扰因素较少,可以有效保证结果的可靠性。同时,仅凭任何单一的指标很难全面整体地对草坪草的抗旱性作出评价,需要对多个代表性指标进行综合分析。本试验中,在干旱胁迫下,‘苏植1号’和‘兰引3号’的叶片萎蔫度和土壤含水量分别具有相同或近似的变化趋势,而胁迫结束时,两个品种的叶片萎蔫度、叶片相对含水量、根系电解质外渗率和土壤含水量均无显著差异。综合上述4个指标来看,干旱胁迫下,‘苏植1号’和‘兰引3号’的外观萎蔫程度、内部水分状况、根系质膜的受损程度以及根系吸水力的维持程度是接近的,因此可以认为,在本试验条件下,两个品种的整体抗旱表现相近。相关研究表明,‘兰引3号’在各种结缕草属植物当中具有较强的抗旱性[14-15,21],而在本试验中发现,‘苏植1号’与‘兰引3号’抗旱性相当。
植物的抗旱性(drought resistance)可分为逃离干旱(drought escape,逃旱)、避免干旱(drought avoidance,避旱)和忍耐干旱(drought tolerance,耐旱)三大策略,每一策略之下又存在多种机制,由于草坪草在干旱环境下仍需维持可接受的坪用质量,因此避旱和耐旱策略显得尤其重要[36]。在应对干旱胁迫的过程中,一方面,草坪草可以通过维持或发展庞大、深长而密集的根系来促进水分和养分的吸收,或者通过降低蒸散率以及促进气孔关闭来减少水分消耗,这些机制属于避旱策略[2,35-41];另一方面,草坪草还能够通过渗透调节、抗氧化防御、碳水化合物代谢和蛋白质代谢等生理生化过程来减轻干旱胁迫的损伤,这些机制属于耐旱策略[5,42-46]。草坪草的抗旱性作为数量性状,受到多个基因、多种代谢过程的控制,同时环境条件对各个基因的表达和各种代谢过程也会施加不同的影响,因此,草坪草最终所表现出来的抗旱性,是一个复杂的综合性状,是在多种因素影响下、多种抗旱机制通过不同的途径发挥作用的结果。
草坪草的根系生长特性(根系生物量和比根长)与根系吸收水分的能力密不可分,是其避旱策略的一个重要部分。干旱胁迫引起高羊茅(Festuca arundinacea)根系干重下降,抗旱性较强的品种下降的幅度较小,干旱还使高羊茅比根长增加,抗旱性较强的品种增加的幅度较大[35];M arcu m等[2]的研究表明,根系深度、根系重量和深层分枝性是结缕草抗旱性的重要机制;但也有研究发现根系生物量与草坪草的抗旱性并不一定是正相关的关系[37]。本试验中,在对照条件或干旱处理下,‘苏植1号’的根系干重均小于‘兰引3号’,而比根长均大于‘兰引3号’,较小的根系干重说明‘苏植1号’的根量较少,不利于水分和养分的吸收,而较大的比根长又使‘苏植1号’的根系扩大了与栽培基质的接触面积,弥补了根量较少的不足。干旱胁迫下两个品种土壤含水量的变化情况,印证了‘苏植1号’和‘兰引3号’根系吸水能力的变化在整体上是同步的。PSⅡ光能转换效率与光合代谢关系紧密,是草坪草耐旱策略的一个重要方面。环境胁迫影响PSⅡ光能转换效率,导致最大光化学量子产量(Fv/Fm)的下降[47];干旱胁迫下,多种草坪草的Fv/Fm值随着胁迫程度加重而呈下降趋势[35,48-50]。本试验中,‘苏植1号’在干旱胁迫中期能够维持比‘兰引3号’更高的Fv/Fm值,这在光合代谢等方面能够发挥积极作用,不过到干旱胁迫末期时,品种间不再具有显著差异。由以上分析推测,虽然‘苏植1号’的根系生物量显著小于‘兰引3号’,但是其较大的比根长和干旱胁迫中期较高的Fv/Fm值很可能使其在一定程度上抵消了较小的根系生物量所带来的不利影响,而在整体表现上具有与‘兰引3号’相当的抗旱性,这种情况说明不同草种的抗旱机制及其途径存在差异。在以往的研究中也有类似发现,比如高羊茅的根系分布既深且广,具备出色的避旱机制,但其渗透调节水平较低,遭受严重干旱之后难以恢复,而结缕草则由于根系相对较浅,避旱性不及高羊茅,但是该草种渗透调节水平较高,有较强的耐旱性,在干旱胁迫结束后能够较快得到恢复[5]。
从整个试验过程来看,‘苏植1号’和‘兰引3号’的叶片萎蔫度和土壤含水量的变化趋势大体上是近似的,但同时也观察到,‘苏植1号’土壤含水量的下降程度在干旱胁迫前期曾一度低于‘兰引3号’,而后者在干旱胁迫中期曾一度出现较高的叶片萎蔫度。推测其原因,可能是由于‘兰引3号’具有较为宽大的叶片,在土壤水分充足时,叶片蒸腾作用较强,造成其在干旱胁迫前期水分消耗较多,引起土壤含水量较早下降,此时,‘兰引3号’借助更为强烈的叶片卷曲来减少蒸腾失水,外观上即表现出比‘苏植1号’更高的叶片萎蔫度,随着干旱胁迫进程的发展,两个品种体内和体外的水分状况渐趋一致,不再具有显著差异。上述现象是避旱策略的一种机制,通过减少水分从叶片散失,草坪草可以在一定程度上保持相对合理的体内水分状况,从而延缓脱水[5,49]。但是,对于草坪草来说,叶片出现卷曲和萎蔫必然影响到草坪质量,因此,在干旱发生时,如何在保证草坪质量的前提下又能够有效地限制从叶片散失的水分,或许更具实践意义。
本研究采用叶片萎蔫度、叶片相对含水量、根系电解质外渗率和土壤含水量这4个指标,以结缕草主栽品种‘兰引3号’为对照,初步评价了‘苏植1号’杂交结缕草的抗旱性,并对两个品种的根系生长特性(根系干重、比根长)和干旱胁迫下最大光化学量子产量(Fv/Fm)的变化特点进行了分析,结果发现,‘苏植1号’在本次试验条件下,其抗旱性与‘兰引3号’相当,同时,本研究还证实了在干旱胁迫下,结缕草属植物不同品种即使具有相似的抗旱表现,却很可能存在不同的抗旱途径,植株的整体抗旱性是多种抗旱机制及其具体抗旱途径共同发挥作用的结果。在实践当中,应根据不同品种的抗旱机制,有针对性地采取适当的栽培管理措施,提高其抗旱性,还可以借助一定的遗传改良手段,把具有不同抗旱途径的品种进行聚合育种,选育出更加抗旱的新种质。当然,要对草坪草不同品种进行抗旱性的全面评价,仍然需要建立在一定数量的代表性种源、严谨的指标筛选以及可靠的分析方法上面,这一部分研究内容有待深入;同时,有关干旱胁迫结束以后草坪草的生长恢复情况及其内在的生理生化机制,也是不同品种总体抗旱性的一个重要方面,在本次试验中未能获得解答;另外,有关PSⅡ光能转换效率如何影响不同品种抗旱性的深层机理,也需进一步探讨与求证。
References:
[1]Zhang Z B,Xu P.W ater and food security in China.Chinese Journal of Eco-A griculture,2008,16(5):1305-1310.
[2]M arcu m K B,Engelke M C,M orton S J,etal.Rooting characteristics and associated drought resistance of zoysiagrasses.A grono m y Journal,1995,87:534-538.
[3]Carrow R N.Drought resistance aspects of turfgrasses in the Southeast:root-shoot responses.Crop Science,1996,36(3):687-694.
[4]Qian Y L,Fry J D.Irrigation frequency affects zoysiagrass rooting and plant water status.H ortScience,1996,31(2):234-237.
[5]Qian Y L,Fry J D.W ater relations and drought tolerance offour turfgrasses.Journal of the A m erican Society for H orticultural Science,1997,122(1):129-133.
[6]W hite R H,Engelke M C,A nderson S J,etal.Zoysiagrass water relations.Crop Science,2001,41(1):133-138.
[7]Li Y,Ling P P,Liu J X,etal.M orphology and distribution diversity of rhizo m e and adventitious root of Zoysia spp.in China.Journal of Plant Resources and Environ m ent,2002,11(2):39-44.
[8]Lu S Y,Chen S P,Chen S M,etal.Responses of proline content and activity of antioxidant enzy m es in warm-season turfgrasses to soil drought stress.Acta H orticulturae Sinica,2003,30(3):303-306.
[9]Liu Z X,Zheng Q Y,W ang X.The form and conductance of leaves sto m a for six turfgrasses.Pratacultural Science,2005,22(8):71-75.
[10]H u H G,Liu J X,M a K Q,etal.Assess m ent of evapotranspiration on Zoysia spp.Acta A grestia Sinica,2006,14(3):206-209,222.
[11]H u H G,Liu J X,Xuan J P,etal.Assess m ent of drought resistance of Zoysia.Acta Prataculturae Sinica,2007,16(1):47-51.
[12]H u H G,Liu J X.M orphological m echanis ms of drought resistance of Zoysia Willd.Acta A grestia Sinica,2008,16(1):94-96.
[13]Li Z D,Li K H,H e H R,etal.Preliminary study on drought resistance and water saving irrigation of 7 warm season turfgrasses in south China.Pratacultural Science,2008,25(11):120-124.
[14]Y u X H,Zhang J M,W ang M Z,et al.Response and resistance of four turfgrasses of Zoysia Willd.to drought stress.Northern H orticulture,2008,(5):121-124.
[15]Zhao Y,Sun J X,W ang Z L.Evaluation on drought resistance of warm-season turfgrass varieties.Journal of Shanghai Jiaotong U niversity(A gricultural Science),2008,26(3):183-187.
[16]W ang Q,Sun J X,A n Y.The effect of the population properties and stress-tolerance physiological characteristics of Zoysia grass under water stresses.Acta Prataculturae Sinica,2009,18(2):33-38.
[17]Lv J,Liu W D,W ang L,etal.Study on drought resistance of four warm-season turfgrasses.Journal of Central South U niversity of Forestry & Technology,2010,30(3):100-104.
[18]Y u L,Liu Y H,Zhou L P,etal.A study on the changes of ascorbic acid and related physiologicalindexes in different cultivars of Zoysia under drought stress.Acta Prataculturae Sinica,2013,22(4):106-115.
[19]H u H G,Zhang Z M,Ji F F,etal.Effects of rewatering on turf quality and physiological recovery of Zoysiagrass.Pratacultural Science,2014,31(6):1058-1062.
[20]H u H G,Zhang Z M,Ji F F,etal.Variation analysis of drought resistance of germ plas m resources of Zoysia Willd.in Chinese m ain region.Acta A grestia Sinica,2014,22(2):224-228.
[21]Xie X C,Xie Y F,Zhang J M,etal.Study on drought resistance of three Zoysiagrasses under drought stress.Guangdong A gricultural Sciences,2014,(17):38-41,46.
[22]H u H G,Zhang Z M,Ji F F,etal.Variation analysis of water use efficiency in Chinese germ plas m of Zoysia Willd.Chinese Journal of Grassland,2015,37(1):45-50.
[23]Song H W,Liu Y,Cao R X,etal.A co m parison of drought resistance of three zoysiagrasses and their new strains induced by60Co-γ radiation.Journal of South China A gricultural U niversity,2015,36(6):55-61.
[24]H uang B R,Duncan R R,Carrow R N.Drought-resistance m echanis ms of seven warm-season turfgrasses under surface soil drying:Ⅰ.Shoot response.Crop Science,1997,37(6):1858-1863.
[25]H uang B R,Duncan R R,Carrow R N.Drought-resistance m echanis ms of seven warm-season turfgrasses under surface soil drying:Ⅱ.root aspects.Crop Science,1997,37(6):1863-1869.
[26]A braha m E M,H uang B R,Bonos S A,etal.Evaluation of drought resistance for Texas bluegrass,Kentucky bluegrass,and their hybrids.Crop Science,2004,44(5):1746-1753.
[27]Zhou Y,La m brides C J,Fukai S.Drought resistance of C4grasses under field conditions:genetic variation a m ong a large nu m ber of berm udagrass(Cynodon spp.)ecotypes collected fro m different clim atic zones.Journal of A grono m y and Crop Science,2013,199(4):253-263.
[28]Pirnajm edin F,M ajidi M M,Gheysari M.Root and physiological characteristics associated with drought tolerance in Iranian tall fescue.Euphytica,2015,202(1):141-155.
[29]Zhu H S,W ang B P,Dong X Y,etal.Effects of drought stress and rewatering on the growth and physiological characteristics of Shanxi wild Poa pratensis seedlings.Acta Prataculturae Sinica,2015,24(7):79-88.
[30]Zhang J M,Zhao M.A co m parative study between turf-type Zoysia japonica cv.Lanyin 3 and Zoysia japonica.Pratacultural Science,1996,13(2):47-50.
[31]Zhang X Z,Ervin E H,Evanylo G K,etal.Im pact of biosolids on horm one m etabolis m in drought-stressed tallfescue.Crop Science,2009,49(5):1893-1901.
[32]Luo N,Liu J X,Y u X Q,etal.Natural variation of drought response in Brachypodium distachyon.Physiologia Plantaru m,2011,141(1):19-29.
[33]Chai Q,Jin F,M erewitz E,etal.Growth and physiologicaltraits associated with drought survival and post-drought recovery in perennial turfgrass species.Journal of the A m erican Society for H orticultural Science,2010,135(2):125-133.
[34]Sun J M,M eyer W,Cross J,etal.Growth and physiological traits of canopy and root syste ms associated with drought resistance in tall fescue.Crop Science,2013,53(2):575-584.
[35]H uang B R,Fry J D.Root anato mical,physiological,and m orphological responses to drought stress for tallfescue cultivars.Crop Science,1998,38(4):1017-1022.
[36]H uang B R,DaCosta M,Jiang Y W.Research advances in m echanis ms of turfgrass tolerance to abiotic stresses:fro m physiology to m olecular biology.Critical Reviews in Plant Sciences,2014,33(2-3):141-189.
[37]H ays K L,Barber J F,Kenna M P,etal.Drought avoidance m echanis ms of selected berm udagrass genotypes.H ortScience,1991,26(2):180-182.
[38]H uang B R,Fry J D.Turfgrass evapotranspiration.Journal of Crop Production,1999,2(2):317-333.
[39]H uang B R,Gao H W.Root physiological characteristics associated with drought resistance in tallfescue cultivars.Crop Science,2000,40(1):196-203.
[40]W ang Z L,H uang B R.Genotypic variation in abscisic acid accu m ulation,water relations,and gas exchange for Kentucky
bluegrass exposed to drought stress.Journal of the A m erican Society for H orticultural Science,2003,128(3):349-355.
[41]Cathey S E,Kruse J K,Sinclair T R,etal.Transpiration and visual appearance of warm season turfgrasses during soil drying.Environ m ental and Experim ental Botany,2013,89:36-43.
[42]Bian S M,Jiang Y W.Reactive oxygen species,antioxidant enzy m e activities and gene expression patterns in leaves and roots of Kentucky bluegrass in response to drought stress and recovery.Scientia H orticulturae,2009,120(2):264-270.
[43]Zhang X Y,H e J G,Sun X H,etal.Transform ation of Lolium perenne with a fructan:fructan 1-fructosyltransferase gene fro m Agropyron cristatum and enhance m ent of drought tolerance in transgenic plants.Acta Prataculturae Sinica,2011,20(1):111-118.
[44]Zhao Y,Du H M,W ang Z L,et al.Identification of proteins associated with water deficit in C4perennial grass species,Cynodon dactylon×Cynodontransvaalensis and Cynodon dactylon.Physiologia Plantaru m,2011,140(1):41-55.
[45]M erewitz E B,Du H M,Y u W J,etal.Elevated cytokinin content in ipt transgenic creeping bentgrass pro m otes drought tolerance through regulating m etabolite accu m ulation.Journal of Experim ental Botany,2012,63(3):1315-1328.
[46]Yang Z M,Xu L X,Y u J J,etal.Changes in carbohydrate m etabolis m in two Kentucky bluegrass cultivars during drought stress and recovery.Journal of the A m erican Society for H orticultural Science,2013,138(1):24-30.
[47]Krause G H,W eis E.Chlorophyllfluorescence and photosynthesis:the basics.A nnual Review of Plant Physiology and Plant Molecular Biology,1991,142:313-349.
[48]Koscielniak J,Filek W,Biesaga-Koscielniak J.The effect of drought stress on chlorophyllfluorescence in Lolium-Festuca hybrids.Acta Physiologiae Plantaru m,2006,28(2):149-158.
[49]Carm o-Silva A E,Soares A S,M arques da Silva J,etal.Photosynthetic responses of three C4grasses of different m etabolic subtypes to water deficit.Functional Plant Biology,2007,34(3):204-213.
[50]Xu C B,Li X M,Zhang L H.The effect of calciu m chloride on growth,photosynthesis,and antioxidant responses of Zoysia japonica under drought conditions.Plos O ne,2013,8(7):e68214.
[1]张正斌,徐萍.中国水资源和粮食安全问题探讨.中国生态农业学报,2008,16(5):1305-1310.
[7]李亚,凌萍萍,刘建秀,等.中国结缕草属植物(Zoysia spp.)地下部分分布和形态类型的多样性.植物资源与环境学报,2002,11(2):39-44.
[8]卢少云,陈斯平,陈斯曼,等.三种暖季型草坪草在干旱条件下脯氨酸含量和抗氧化酶活性的变化.园艺学报,2003,30(3):303-306.
[9]刘自学,郑群英,汪玺.6种草坪草叶片的气孔特征与气孔传导力.草业科学,2005,22(8):71-75.
[10]胡化广,刘建秀,马克群,等.结缕草属植物蒸散量评价.草地学报,2006,14(3):206-209,222.
[11]胡化广,刘建秀,宣继萍,等.结缕草属植物的抗旱性初步评价.草业学报,2007,16(1):47-51.
[12]胡化广,刘建秀.结缕草属植物抗旱形态机理.草地学报,2008,16(1):94-96.
[13]李志东,黎可华,何会蓉,等.华南地区7个暖季型草坪草种的抗旱性与灌溉节水的初步研究.草业科学,2008,25(11):120-124.
[14]余晓华,张巨明,王明祖,等.四种结缕草属草坪草对土壤干旱胁迫的响应及抗旱性研究.北方园艺,2008,(5):121-124.
[15]赵艳,孙吉雄,王兆龙.狗牙根和结缕草部分品种的抗旱性能评价.上海交通大学学报(农业科学版),2008,26(3):183-187.
[16]王齐,孙吉雄,安渊.水分胁迫对结缕草种群特征和生理特性的影响.草业学报,2009,18(2):33-38.
[17]吕静,刘卫东,王丽,等.4种暖季型草坪草的抗旱性分析.中南林业科技大学学报,2010,30(3):100-104.
[18]俞乐,刘拥海,周丽萍,等.干旱胁迫下结缕草叶片抗坏血酸与相关生理指标变化的品种差异研究.草业学报,2013,22(4):106-115.
[19]胡化广,张振铭,季芳芳,等.复水处理对结缕草草坪质量和生理恢复的影响.草业科学,2014,31(6):1058-1062.
[20]胡化广,张振铭,季芳芳,等.中国主要地区结缕草属植物种质资源抗旱性变异分析.草地学报,2014,22(2):224-228.
[21]谢新春,解彦峰,张巨明,等.干旱胁迫下3种结缕草的抗旱性分析.广东农业科学,2014,(17):38-41,46.
[22]胡化广,张振铭,季芳芳,等.中国结缕草属植物种质资源水分利用效率变异分析.中国草地学报,2015,37(1):45-50.
[23]宋华伟,刘颖,曹荣祥,等.3种结缕草及其60Co-γ辐射诱变新品系抗旱性比较.华南农业大学学报,2015,36(6):55-61.
[29]朱慧森,王保平,董晓燕,等.干旱及复水对山西野生草地早熟幼苗生长与生理特性的影响.草业学报,2015,24(7):79-88.
[30]张巨明,赵鸣.兰引3号草坪型结缕草与日本结缕草的比较研究.草业科学,1996,13(2):47-50.
[43]张小芸,何近刚,孙学辉,等.转果聚糖合成关键酶基因多年生黑麦草的获得及抗旱性的提高.草业学报,2011,20(1):111-118.
Preliminary evaluation of drought resistance for the hybrid zoysiagrass‘Suzhi No.1’
A n experim ent was carried out to evaluate the drought resistance of hybrid zoysiagrass‘Suzhi N o.1’,co m pared with Zoysiajaponica‘Lanyin N o.3’as the control.Plant m aterials were cultured in sand in P V C tubes in a greenhouse and drought treatm ents were im ple m ented by withholding water.Leaf wilting(L W),leaf relative water content(R W C),root electrolyte leakage(R E L),soil water content(S W C),root growth characteristics including root dry weight(R D W)and specific root length(S R L),as well as m axim u m quantu m yield of PSⅡ(Fv/Fm)were m easured under drought stress.T he drought resistance of‘Suzhi N o.1’was initially evaluated based on L W,R W C,R E L and S W C.Both cultivars showed gradual increases in L W,exhibiting slight wilting on the 12th day of drought stress,m oderate wilting on the 29th day and severe wilting after 40 days.S W C values decreased gradually with both cultivars displaying similar trends during the stress period.Drought stress also caused significant declines in R W C and rises in R E L.By the end of the treatm ent(40th day),no differences in L W,R W C,R E L and S W C were found indicating that the perform ance of‘Suzhi N o.1’was similar to‘Lanyin N o.3’under drought stress.Although‘Suzhi N o.1’had s m aller R D W to‘Lanyin N o.3’,it also had larger S R L and could keep higher Fv/Fmvalues in the middle of the stress period,potentially an im portant drought resistance m echanis m.With relatively denser roots and higher photoche mical efficiency,itis probable that‘Suzhi N o.1’was able to offset the adverse effects of s m aller root bio m ass.In su m m ary,‘Suzhi N o.1’and‘Lanyin N o.3’had similar drought resistance but with different resistant m echanis m s.
hybrid zoysiagrass‘Suzhi N o.1’;drought resistance;evaluation;root growth characteristic;m axim al quantu m yield of PSⅡ
.E-m ail:turfunit@aliyun.com
10.11686/cyxb2015356
2015-07-20;改回日期:2016-02-25
国家自然科学基金海外及港澳学者合作研究基金(31328019),江苏省水利科技项目(2014001,2014019)和江苏省科技基础设施建设项目(B M 2015019)资助。
汪毅(1974-),男,江苏南通人,实验师,在职硕士。E-m ail:wang-yi-8@hotm ail.com
猜你喜欢
作物研究(2022年4期)2022-09-21
今日农业(2021年14期)2021-11-25
今日农业(2020年13期)2020-12-15
中华养生保健(2020年4期)2020-11-16
建材发展导向(2019年11期)2019-08-24
西北林学院学报(2018年4期)2018-08-02
河南水利年鉴(2017年0期)2017-05-19
浙江农业学报(2017年1期)2017-05-17
灾害医学与救援(电子版)(2017年3期)2017-02-06
安徽医药(2014年4期)2014-03-20
