
应用拉氏图信息提高短乳杆菌谷氨酸脱羧酶催化性能
2016-07-04 07:33柯丕余黄俊胡升赵伟睿吕常江郁凯雷引林王进波梅乐和
生物工程学报 2016年1期
关键词:氨基丁酸
柯丕余,黄俊,胡升,赵伟睿,吕常江,郁凯,雷引林,王进波,梅乐和,
1 浙江大学 化学工程与生物工程学院,浙江 杭州 3100272 浙江科技学院 生物与化学工程学院,浙江 杭州 3100233 浙江大学宁波理工学院 生物与化学工程学院,浙江 宁波 315100
应用拉氏图信息提高短乳杆菌谷氨酸脱羧酶催化性能
柯丕余1,黄俊2,胡升3,赵伟睿1,吕常江1,郁凯1,雷引林3,王进波3,梅乐和1,3
1 浙江大学 化学工程与生物工程学院,浙江 杭州 310027
2 浙江科技学院 生物与化学工程学院,浙江 杭州 310023
3 浙江大学宁波理工学院 生物与化学工程学院,浙江 宁波 315100
柯丕余, 黄俊, 胡升, 等. 应用拉氏图信息提高短乳杆菌谷氨酸脱羧酶催化性能. 生物工程学报, 2016, 32(1): 31–40.
Ke PY, Huang J, Hu S, et al. Enhancing glutamate decarboxylase activity by site-directed mutagenesis: an insight from Ramachandran plot. Chin J Biotech, 2016, 32(1): 31–40.
摘 要:谷氨酸脱羧酶 (Glutamate decarboxylase,GAD) 是用于催化L-谷氨酸脱羧合成γ-氨基丁酸(γ-aminobutyrate,GABA) 的唯一酶,提高GAD的催化活力或热稳定性,有利于GABA的高效制备和生产。以热稳定性和活性为筛选目标,通过研究短乳杆菌GAD1407三维模拟结构的拉氏图,确定不稳定氨基酸残基位点K413,采用定点突变的方法构建该位点的突变体,并测定野生型酶和突变酶的热稳定性和活力。结果表明突变酶K413A和突变酶K413I分别在热稳定性和酶活力上获得了提高,突变酶K413A在50 ℃的半衰期为105 min,是野生酶的2.1倍;突变酶K413I热稳定性没有明显的提高,但其酶活力却得到了有效提高,约为野生型的1.6倍。因此,通过拉氏图提供的结构信息可为利用理性设计提高GAD活性和热稳定性提供指导。
关键词:谷氨酸脱羧酶,蛋白质工程,γ-氨基丁酸,拉氏图
Received: March 25, 2015; Accepted: April 27, 2015
Supported by: National Natural Science Foundation of China (Nos. 21176220, 31240054, 31470793), Natural Science Foundation of Zhejiang Province (No. Z13B060008), Natural Science Foundation of Ningbo (No. 2013A610087).
国家自然科学基金 (Nos. 21176220, 31240054, 31470793),浙江省自然科学基金 (No. Z13B060008),宁波市自然科学基金 (No. 2013A610087) 资助。
网络出版时间:2015-05-25 网络出版地址:http://www.cnki.net/kcms/detail/11.1998.Q.20150525.1045.002.html
γ-氨基丁酸 (γ-aminobutyric acid,GABA)是一种天然存在的非蛋白氨基酸,是哺乳动物中枢神经系统中一种十分重要的抑制性神经递质,具有降血压、利尿、抗惊厥、预防癫痫、改善睡眠、抗抑郁、促进激素分泌和保肝利肾等多种生理功能[1-2]。在工业应用中,GABA可作为中间体来生产可降解生物塑料尼龙-4[3]以及N-吡咯烷酮[4]等化工产品。目前,GABA的制备方法主要有化学合成法、植物富集法和微生物转化法,由于微生物转化法具有生产周期短、反应条件温和、产量高以及环境友好等优点,因此利用GAD或具有该酶活力的细胞进行GABA的生物制备具有重要的工业生产应用价值[5-6]。谷氨酸脱羧酶 (Glutamate decarboxylase,GAD;EC 4.1.1.15) 可以磷酸吡哆醛 (PLP) 为辅酶专一性地催化L-谷氨酸的α-羧基脱羧生成γ-氨基丁酸[7]。但由于天然的谷氨酸脱羧酶存在酶活低和热稳定性差的缺陷,在实际应用中受到了很大的限制。
对天然酶的理化性质进行改造以获取催化效率更高、选择性更强、稳定性更好的新酶,对于酶的工业应用有着十分重要的意义。首先,良好的热稳定性意味着酶不易失活,保存期限更久。其次,耐热性强和活性高的酶可更广泛的应用于工业生产。目前已有很多种属的GAD基因被克隆并重组表达,对GAD基因进行研究与改造能从本质上提高GAD的催化活力和稳定性,是提高微生物转化法制备GABA产量最直接和最有效的手段[8-10]。例如,Lin等[11]以短乳杆菌CGMCC No. 1306的GAD为研究对象,通过采用易错PCR结合定点饱和突变的方法,获得了一个催化活力得到显著提高的突变酶Q51H,在pH 4.8时其催化效率 (kcat/Km) 是野生酶的2.56倍。Shi等[12]通过对短乳杆菌Lactobacillus brevis Lb85进行易错PCR和定点突变等手段,筛选出GadB1E312S和GadB1T17I/D294G/Q346H突变体,在pH 6.0时它们的催化效率 (kcat/Km) 分别是野生型的2.30倍和25.26倍,这些突变位点的替换能够有效地拓宽酶的催化pH范围和提高其 GABA的合成能力。Jun等[13]对大肠杆菌Escherichia coli中的GAD 进行三维结构分析,发现GAD同型六聚体N-末端14个残基形成三重螺旋束有助于提高该酶的热稳定性,通过优化N-末端结构域间的疏水和静电相互作用,突变酶GadB-TMQ5D/V6I/T7E显示出良好的热稳定性,其半失活温度 (T5015) 较野生型酶提高7.7 ℃。
在描述肽键的刚性和可变程度时,研究者常运用二面角 (φ,ψ) 来确定α碳原子和肽平面间单键的旋转。由于不同氨基酸残基可允许的φ和ψ值范围不同,以φ和ψ作为横纵坐标得到的二维图形称为拉氏图 (Ramachandran plot)。由于拉氏图中不同区域显示不同的结构稳定性,利用这些信息,可以找出蛋白质骨架中的不稳定残基位点,并通过定点突变等方式改善酶的结构,进而改变酶活力或者热稳定性[14]。例如,Mahdieh等[15]利用拉氏图信息寻找Chondroitinase ABCⅠ三维结构中不合理的残基位点Q140,并对其进行定点突变,得到的突变酶Q140A在40 ℃下的半衰期为7 min,约为野生型的3.5倍。
作者所在的研究机构在前期研究中已经从GABA高产菌株短乳杆菌Lb. brevis CGMCC No. 1306中克隆了GAD1407基因,并实现了其在大肠杆菌中的可溶表达[16],但该酶尚存在酶活低和热稳定性差的缺陷,为了进一步提高其催化性能及热稳定性,本文拟通过构建GAD1407结构拉氏图,并分析酶分子中氨基酸残基的构象分布,根据拉氏图提供的信息,确定了结构不稳定点,结合定点突变技术对其进行改造,以期提高正突变概率和实验效率。
1 材料与方法
1.1材料
1.1.1酶与试剂
Fast Digest DpnⅠ限制酶、Fast Pfu DNA聚合酶、1 kb DNA Ladder marker均为TaKaRa公司产品 (宝生物工程 (大连) 有限公司,中国);质粒提取试剂盒、PCR产物纯化试剂盒、考马斯亮蓝蛋白质浓度测定试剂盒购自生工生物工程 (上海) 股份有限公司 (中国);DNA凝胶回收试剂盒、Ni-NTA层析介质购自北京全式金生物技术有限公司 (中国);种子培养基为LB培养基,表达培养基为TB培养基,均含有50 μg/mL卡那霉素。
1.1.2菌株与质粒
表达宿主菌E. coli BL21 (DE3)、pET28a(+)-GAD1407重组质粒为浙江大学生物工程研究所保藏。
1.1.3培养基与溶液配制
LB培养基:10 g/L胰蛋白胨,5 g/L酵母粉,10 g/L氯化钠。
TB培养基:12 g/L胰蛋白胨,24 g/L酵母粉,4 mL/L甘油,17 mmol/L磷酸二氢钾,72 mmol/L磷酸氢二钾。
破胞缓冲液 (pH 7.4):2 mmol/L磷酸二氢钾,10 mmol/L磷酸氢二钠,2.7 mmol/L氯化钾,1 mmol/L苯甲基磺酰氟,137 mmol/L氯化钠。
1.2突变文库的构建
根据Lb. brevis CGMCC No. 1306的谷氨酸脱羧酶基因设计19对定点突变引物,如表1所示。
以含有GAD1407基因的质粒为模板,进行定点PCR扩增,PCR反应体系为:10 μL 5×PCR缓冲液,4 μL dNTPs (2.5 mmol/L),1 μL上游引物 (10 mmol/L),1 μL下游引物 (10 mmol/L),1 μL质粒模板 (50 ng/μL),Fast Pfu DNA聚合酶5 U,高温灭菌的超纯水补至总体积50 μL。定点PCR扩增程序:94 ℃变性5 min;94 ℃变性30 s,55 ℃退火30 s,72 ℃延伸4 min,30个循环;72 ℃延伸4 min。所得到的PCR产物经PCR产物纯化试剂盒纯化后用DpnⅠ消化0.5 h,消化产物继续采用PCR产物纯化试剂盒纯化,然后用热激法将PCR产物以10%比例 (V/V)转化至E. coli BL21 (DE3) 感受态细胞中,转化液涂布于含有50 μg/mL卡那霉素的LB固体培养基中,37 ℃培养过夜,即得定点突变文库。
1.3野生型酶和突变酶表达和纯化
从19种氨基酸的定点突变文库中分别随机挑取1–3个单菌落,培养并提取质粒,测定核苷酸序列,以确定是否全部引入预期的突变。将经测序鉴定的质粒转化到E. coli BL21 (DE3)中,挑取单菌落接种至含有50 μg/mL卡那霉素的5 mL LB液体培养基中,37 ℃、200 r/min条件下培养过夜,再将此过夜培养物以2%比例(V/V) 的接种量接种至含50 μg/mL卡那霉素的100 mL的TB培养基中,37 ℃培养至OD600值为0.6–0.8时,加入适量体积的IPTG至终浓度为0.5 mmol/L,然后在25 ℃、150 r/min条件下诱导培养8 h后收集菌体。
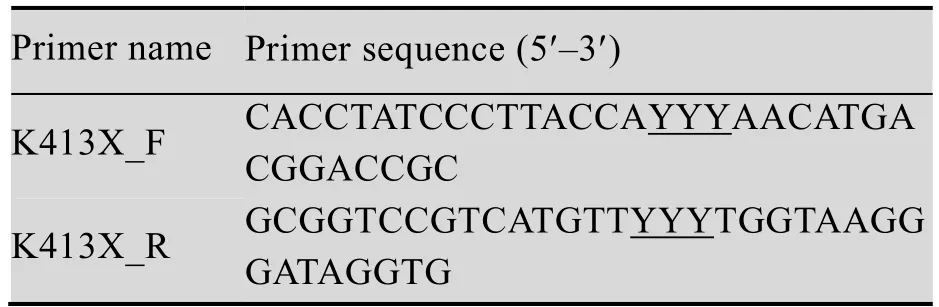
表1 定点突变引物及其序列Table 1 Primer used for site-directed mutagenesis
1.4粗酶液制备和纯化
粗酶液制备:将收集的菌体用磷酸盐缓冲液洗涤两次,后用10%发酵液体积的破胞缓冲液重悬,超声波破碎细胞,超声破胞工作条件为:功率300 W,工作3 s,间歇6 s,循环90次。经破碎后的悬液于10 000 r/min、4 ℃条件下离心处理30 min,收集上清液,即粗酶液。
蛋白纯化:采用Ni-NTA亲和层析对所得的粗酶液进行分离纯化,经上样、清洗和洗脱,收集洗脱液,透析去除小分子得到纯酶,采用SDS-PAGE检测纯化后的蛋白纯度。用考马斯亮蓝法测定纯化后的蛋白浓度。
1.5突变文库筛选
将上述成功引入目的突变的19种菌株,分别经培养、表达、制备粗酶、纯化得到纯酶,然后在20 ℃、55 ℃两个温度下处理10 min,冰浴5 min,离心取上清液,测定野生型和突变酶残余活力,将55 ℃处理后的比活力对比20 ℃处理后的比活力,获得两个温度下的残余活力比值,筛选出残余活力比值高于野生型酶残余活力比值的突变酶。
1.6野生型酶和突变酶酶学参数及热稳定性测定
动力学参数测定:用0.2 mol/L的醋酸-醋酸钠缓冲液 (pH 4.8) 配制不同浓度 (1–100 mmol/L)的底物L-谷氨酸钠 (L-MSG),测定在不同底物浓度条件下的反应初速度。将不同底物浓度[S]下对应的反应速率[V]代入米氏方程 (1-1):
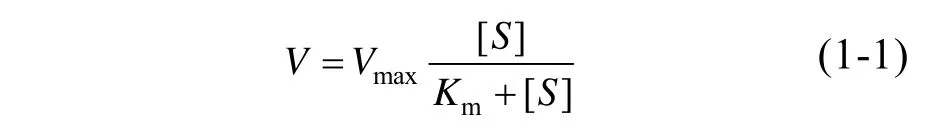
通过Origin 8.0软件非线性拟合计算相应的Km和Vmax值,然后根据kcat=Vmax/[E0] ([E0]为酶初始浓度,单位μmol/L) 计算求得kcat和催化效率kcat/Km。
热失活的半衰期 (t1/2) 测定条件:将1 mg/mL的野生型酶或突变酶,分别于PBS缓冲液 (pH 7.4) 中50 ℃条件下保温不同时间取样,测定残余酶活,用Origin 8.0软件拟合热失活曲线,计算半衰期t1/2。
比活力:定义为每分钟每毫克蛋白生成的GABA量。取纯化的酶液15 μL (1 mg/mL),加入400 μL底物溶液中,于48 ℃反应10 min,反应结束后迅速放入沸水浴中10 min以终止反应,然后将样品离心,收集上清液,采用高效液相色谱法测定产物中GABA含量,具体参照文献17。
2 结果与分析
2.1GAD1407的拉试图构建及分析
根据郁凯[18]构建的GAD1407三维结构模型,如图1所示是Swiss Model方法建立的结构模型图。该模型的结构特征主要有以下部分:Lb. brevis CGMCC No. 1306 GAD的整体结构呈现α/β折叠类型,共有12个α-螺旋和4组β-折叠,相邻的二级结构之间由Loop相连接。此外,利用PROCHECK程序进行二面角合理性的评价,生成的拉氏图如图2所示。
拉氏图中的每一个区域的颜色从深到浅排列的次序依次为最佳合理区 (Most favored regions)、额外合理区 (Additional allowed regions)、一般合理区 (Generously allowed regions) 以及不合理区 (Disallowed regions)。如图2可知,413位点Lys在MODELLER和Swiss Model两种建模方法得到的结构评价均处于构象不稳定区,说明413位点的构象中非共价键合原子间的距离小于极限距离,斥力很大,构象能量高,所以很不稳定。此外,由GAD1407的三维结构模拟图 (图1) 可知,第413个位点(Lys) 位于C端的一个β-转角上,远离酶的催化中心 (PLP-Lys279)。
2.2定点突变基因测序和筛选
从每个氨基酸相应定点突变文库中挑取单菌落,培养后提取质粒进行DNA测序。
将成功引入氨基酸突变的菌株进行培养、表达、纯化后,按照1.5突变文库初筛,获得55 ℃处理10 min后野生型酶与突变酶残余活力对比图 (图3) 以及20 ℃处理10 min后野生型酶与突变酶相对活力对比图 (图4)。
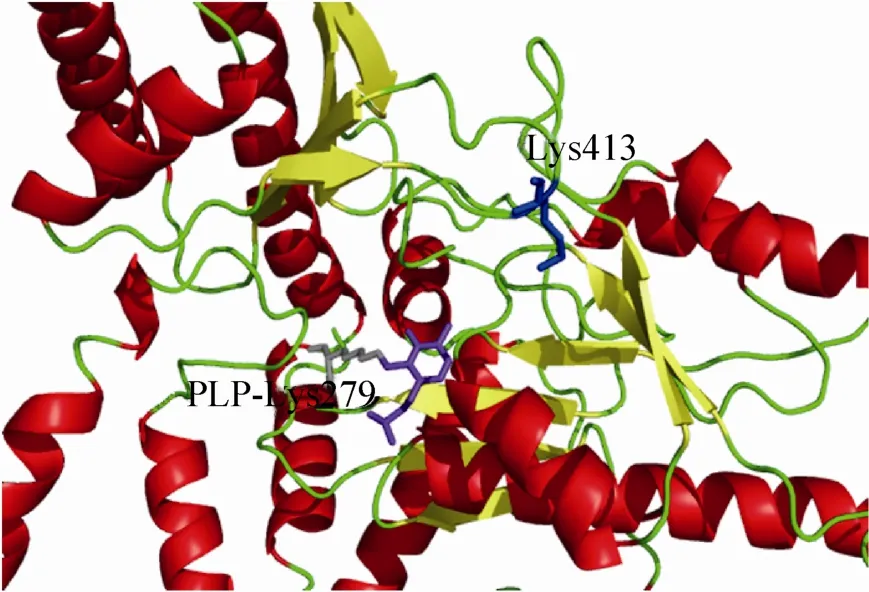
图1 GAD1407的三维结构模拟图Fig. 1 Three-dimensional structures of GAD 1407 by homology modelling. PLP is shown as purple sticks, which forms a Schiff base with Lys 279 (gray sticks), Site 413 (Lys) is shown as blue sticks.
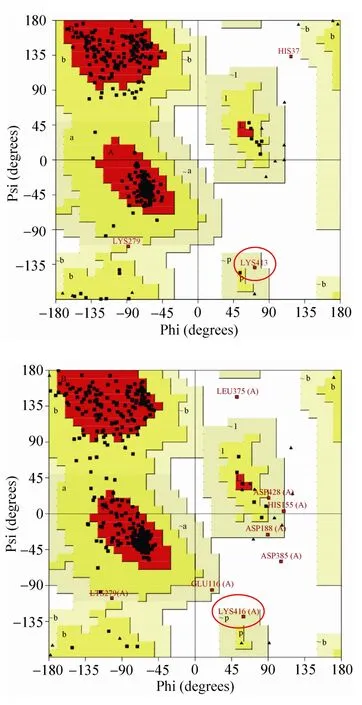
图2 MODELLER (上) 和Swiss Model (下) 建模结果的拉氏图Fig. 2 Ramachandran plot of GAD 1407 model generated by MODELLER (up) and Swiss Model (down).
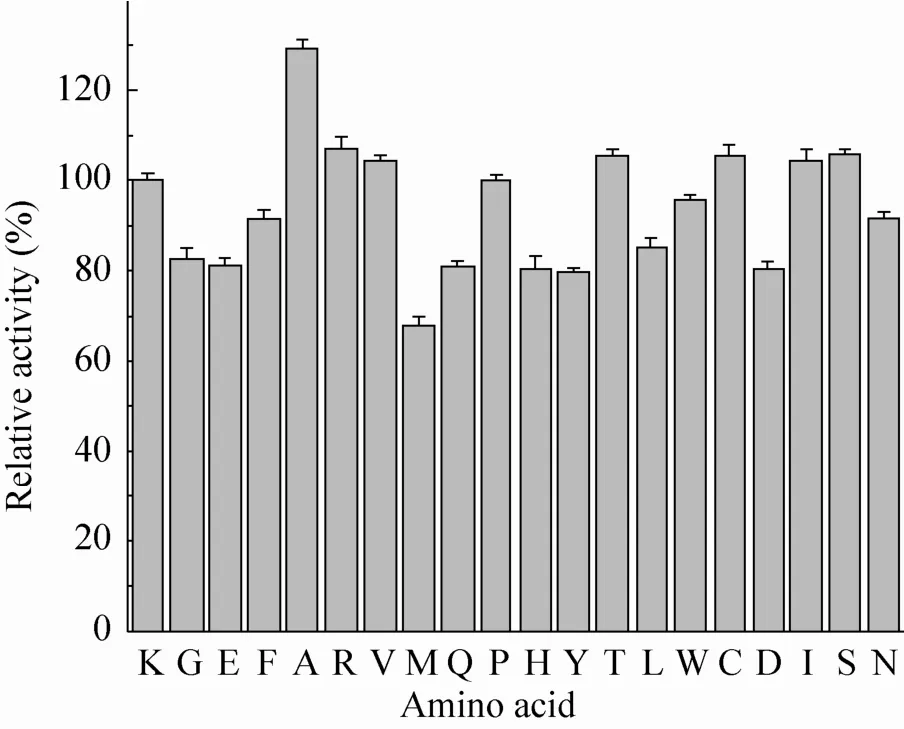
图3 55 ℃处理10 min后野生型酶与突变酶活力对比Fig. 3 The relative activity of wide type and mutant GAD at 55 ℃. Enzymes were heated at 55 ℃ for 10 min, after which residual activities were determined taking L-MSG as substrate. The error bars show standard deviation calculated for three replicate experiments.
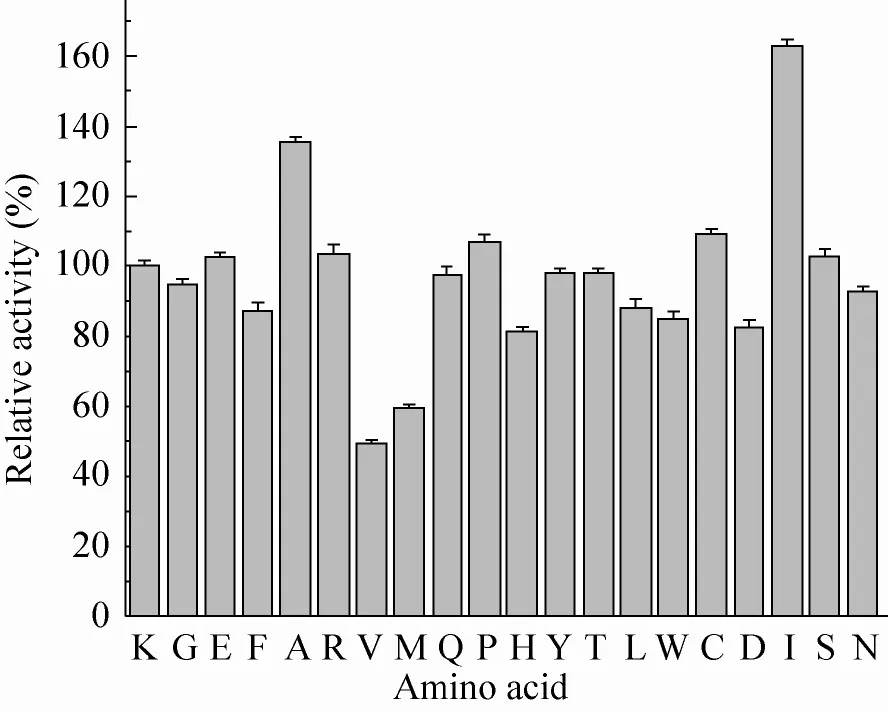
图4 20 ℃处理10 min后野生型酶与突变酶活力对比Fig. 4 The relative activity of wide type and mutant GAD at 20 ℃. Enzymes were heated at 20 ℃ for 10 min, after which residual activities were determined taking L-MSG as substrate. The error bars show standard deviation calculated for three replicate experiments.
由图3可知,55 ℃处理10 min后,当413位点突变为丙氨酸 (Ala) 时,突变酶K413A残余活力约为野生型酶的1.23倍,这说明其能有效提高酶在55 ℃条件下的热稳定性。同时在常温下,突变酶K413A的活力也是野生型酶的1.35倍。此外,由图4可知,突变酶K413I在活力上也有很好的提升,其他突变酶在活力上的变化并不明显。故根据上面的结果,筛选出2个突变体,即突变酶K413I和突变酶K413A,进行后续的实验。
2.3野生型酶和突变酶表达及纯化
根据2.2中的初筛结果,突变酶K413A和突变酶K413I分别在耐热性和酶活力上有一定提高,它们和野生型酶的SDS-PAGE检测结果如图5所示,纯酶的蛋白质分子量大小为56 kDa。
2.4野生型酶和突变酶的热稳定性
考察野生型酶、突变酶K413A和突变酶K413I的热稳定性,结果如图6所示。
由图6可知,将纯化后的野生型酶、突变酶K413A和突变酶K413I于50 ℃下处理不同的时间 (0–120 min),然后置于冰浴中5 min,测定残余酶活。用Origin 8.0软件拟合热失活曲线,可以看出野生型、突变酶K413A和突变酶K413I的热稳定半衰期分别约为50、105、46 min,说明突变酶K413A较野生型热稳定性有了较大提高,而突变酶K413I较野生型热稳定性略微降低 (突变酶K413A>野生型>突变酶K413I),说明在413位点引入丙氨酸可以明显地提高热稳定性,而将赖氨酸替换成异亮氨酸则有热稳定性降低的趋势。
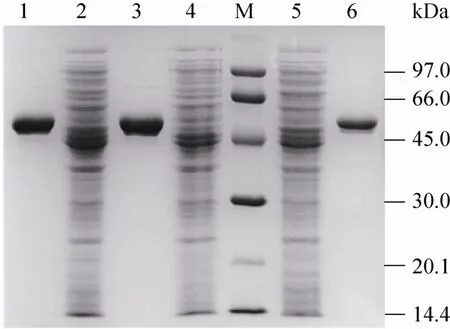
图5 野生型酶和突变酶的SDS-PAGE检测结果图Fig. 5 SDS-PAGE analysis of wide type and mutant GAD. M: protein marker; 1: purified wide type; 2: supernatant of wide type; 3: purified K413A; 4: supernatant of K413A; 5: supernatant of K413I; 6: purified K413I.
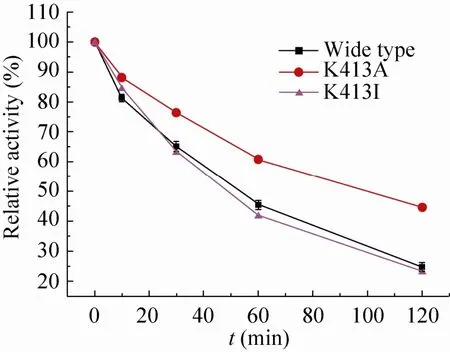
图6 野生型酶和突变酶在50 ℃下的热稳定性Fig. 6 Thermostability of wide type and mutant GAD proteins at 50 ℃. Enzymes were heated at 50 ℃ for increasing time, after which residual activities were determined taking L-MSG as substrate. The error bars show standard deviation calculated for three replicate experiments.
2.5野生型酶和突变酶的动力学参数测定
以L-MSG为底物,在pH 4.8、48 ℃的条件下,分别测定野生型酶、突变酶K413A和突变酶K413I的米氏动力学常数。按照1.6所述,在酶含量为1 mg/mL,底物浓度为1−100 mmol/L的条件下测定催化速率,根据米氏方程,采用非线性拟合对数据进行分析,计算获得酶动力学参数,结果见表2。
从表2中可知,突变酶K413A和突变酶K413I的米氏常数Km值与野生型酶的Km值基本相同,而kcat值均有一定提高,突变酶K413A和突变酶K413I的kcat值分别是野生型酶的133%和167%,此外,酶的催化效率kcat/Km也有不同程度的提升,突变酶K413A和突变酶K413I的kcat/Km分别是野生型酶的139%和156%,其中突变酶K413I的kcat值比野生型酶的kcat值提高了67%,这说明突变酶K413I是通过催化效率的提高而获得较大的比活力。

表2 野生型酶和突变酶酶学参数比较Table 2 Kinetic analysis of wide type and mutant GAD proteins
3 讨论
本研究通过构建并分析了GAD1407的结构拉氏图,发现413位点Lys在MODELLER和Swiss Model两种建模方法得到的结构中均处于构象不稳定区,说明Lys在该位点的构象中非共价键合原子间的距离小于极限距离,斥力很大,构象能量高,结构不稳定,所以对该位点进行了定点突变,成功地筛选到了突变酶K413A和突变酶K413I。
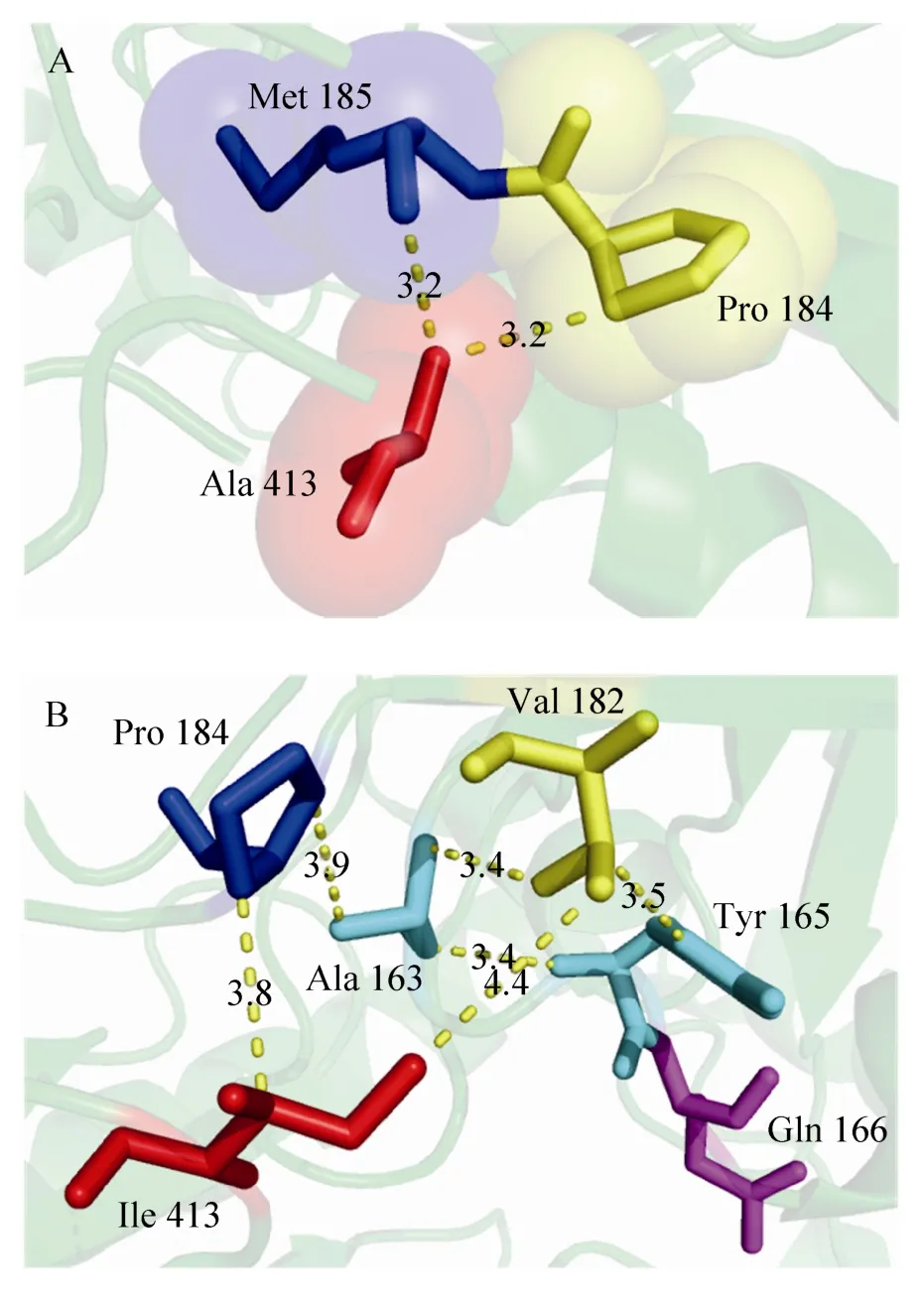
图7 K413A (A) 和K413I (B) 的三维结构模拟图Fig. 7 Three-dimensional structures of K413A and K413I by homology modelling. (A) In the mutant K413A, substituted Ala was shown to involve in hydrophobic interaction with the neighbouring hydrophobic amino acids M185 and P184. (B) In the mutant K413I, a view of the hydrophobic interaction among the substituted Ile, V182, P184, A163 and Y165 was shown.
实验结果表明,在50 ℃处理条件下,突变酶K413A的半衰期为105 min,是野生型酶 (50 min) 的2.1倍,这个单一氨基酸位点突变成功地提高了GAD的热稳定性。图1和图7A分别是野生型酶和突变酶K413A的局部三维结构模拟图,当亲水氨基酸K (Lys) 替换成疏水氨基酸A (Ala)之后,将野生型酶和突变酶K413A的PDB文件提交至在线服务器 (http://pic.mbu.iisc.ernet.in/),选取疏水相互作用选项 (距离设为5 Å) 进行计算[19],可以发现突变酶K413A主要增加了蛋白质分子内的疏水作用,例如野生型酶413位点是赖氨酸,并不能与其他相邻的氨基酸残基形成疏水相互作用,但当赖氨酸替换成丙氨酸后,蛋白质分子内增加了A413与A163、P184、M185、P412和M415之间的疏水相互作用,其中A413与P184、M185之间的空间距离小于4 Å,并且上述3个氨基酸的空间上出现了重叠区域。Kellis等[20]和Pace等[21]等通过系统地分析22个蛋白质中和疏水相互作用有关的148个突变体发现,疏水相互作用对蛋白质稳定性的贡献约为60%±4%,而氢键对蛋白质稳定性的贡献约为40%±4%,这明确表明疏水相互作用是蛋白质折叠过程中最重要的驱动力,所以预测很可能是增加的蛋白质分子内疏水相互作用使得突变酶K413A的热稳定性得到了提高。
突变酶K413I热稳定性虽然没有明显的提高,但是在最适pH 4.8条件下的催化活力约为野生型的1.6倍。根据GAD1407的三维结构模型,如图1和图7B所示,K413位点位于C端的一个β-转角上,并且远离活性中心,并不直接参与催化反应,提高酶活力的原因可能是:当413位点的赖氨酸替换成异亮氨酸之后,碱性氨基酸被替换成疏水氨基酸,突变后的I413 与V182 (4.5 Å)、P184 (3.8 Å) 之间存在疏水相互作用,P184与A163 (3.9 Å),V182与Y165 (3.5 Å)、A163 (3.4 Å) 均有相互疏水作用,这使得该区域的空间结构发生轻微的变化,进而影响到了与Y165相连的Q166,Q166直接参与GAD的催化反应[16],故猜测K413替换成I413能改善酶的活力是由于疏水相互作用。
以上位点突变对于GAD1407催化性能的改善作用均为推测性分析,需要后续的研究,尤其是获得GAD1407的晶体结构,来确定分析的准确性。
为了能够有效提高突变概率,常通过理性设计预测重要的氨基酸突变位点,用于节约时间,提高实验效率及可行性。本文建立了一种基于拉氏图获取结构信息,从而预测有效突变位点的方法,并且结合定点突变技术,筛选得到了催化活力或热稳定性优于野生型酶的突变酶。由于没有确切的GAD1407蛋白质结构信息,所以我们还未能说明某一位点突变引起酶学性质改变的确切机制,只能通过模拟的蛋白结构进行猜测,但是本研究基于蛋白质拉氏图的理性设计的策略为谷氨酸脱羧酶酶学性质的提高提供了新的思路。
REFERENCES
[1] Liang HY, Deng LK, Lin HL, et al. Selection of lactic acid bacteria for producting γ-amino butyric acid and optimization of fermentation. Food Res Dev, 2008, 24(1): 36–40 (in Chinese).梁恒宇, 邓立康, 林海龙, 等. 新资源食品-γ-氨基丁酸 (GABA) 的研究进展. 食品研究与开发, 2008, 24(1): 36–40.
[2] Schuller HM, Al-Wadei HAN, Majidi M. Gammaaminobutyric acid, a potential tumor suppressor for small airway-derived lung adenocarcinoma. Carcinogenesis, 2008, 29(10): 1979–1985.
[3] Takahashi C, Shirakawa J, Tsuchidate T, et al. Robust production of gamma-amino butyric acid using recombinant Corynebacterium glutamicum expressing glutamate decarboxylase from Escherichia coli. Enzyme Microb Technol, 2012, 51(3): 171–176.
[4] Lammens TM, Franssen MCR, Scott EL, et al. Synthesis of biobased N-methylpyrrolidone by one-pot cyclization and methylation of γaminobutyric acid. Green Chem, 2010, 12(8): 1430–1436.
[5] Huang J, Mei LH, Sheng Q, et al. Optimization of γ-aminobutyric acid liquid fermentation conditions and its fed-batch fermentation. J Chem Eng Chin Univ, 2008, 22(4): 618–623 (in Chinese).黄俊, 梅乐和, 盛清, 等. γ-氨基丁酸液体发酵过程的条件优化及补料研究. 高校化学工程学报, 2008, 22(4): 618–623.
[6] Lu YM, Zhang WG. Selection of lactic acid bacteria for producing γ-amino butyric acid and optimization of fermentation. Food Machinery, 2008, 24(1): 36–40 (in Chinese).卢彦梅, 张伟国. γ-氨基丁酸产生菌的选育及发酵条件优化. 食品与机械, 2008, 24(1): 36–40.
[7] Kim H, Kashima Y, Ishikawa K, et al. Purification and characterization of the first archaeal glutamate decarboxylase from Pyrococcus horikoshii. Biosci Biotechnol Biochem, 2009, 73(1): 224–227.
[8] Lin L. Enhancing the activity of glutamate decarboxylase by directed evolution and semirational evolution[D]. Hangzhou: Zhejiang University, 2010 (in Chinese).林玲. 利用定向进化及半理性设计提高谷氨酸脱羧酶催化活性的研究[D]. 杭州: 浙江大学, 2013.
[9] Fan EY, Huang J, Hu S, et al. Cloning, sequencing and expression of a glutamate decarboxylase gene from the GABA-producing strain CGMCC 1306. Ann Microbiol, 2012, 62(2): 689–698.
[10] Tian LZ, Xu MJ, Rao ZM, et al. Construction of a recombinant Escherichia coli BL21/pET-28alpgad and the optimization of transformation conditions for the efficient production of γaminobutyric acid. Chin J Biotech, 2012, 28(1): 65–75 (in Chinese).田灵芝, 徐美娟, 饶志明, 等. 一株重组大肠杆菌/pET-28a-lpgad的构建及其高效生产γ-氨基丁酸转化条件的优化. 生物工程学报, 2012, 28(1): 65–75.
[11] Lin L, Hu S, Yu K, et al. Enhancing the activity of glutamate decarboxylase from Lactobacillus brevis by directed evolution. Chin J Chem Eng, 2014, 22(11/12): 1322–1327
[12] Shi F, Xie YL, Jiang JJ, et al. Directed evolution and mutagenesis of glutamate decarboxylase from Lactobacillus brevis Lb85 to broaden the range of its activity toward a near-neutral pH. Enzyme Microb Technol, 2014: 35–43.
[13] Jun CH, Joo JC, Lee JH, et al. Thermostabilization of glutamate decarboxylase B from Escherichia coli by structure-guided design of its pH-responsive N-terminal interdomain. J Biotechnol, 2014, 174: 22–28
[14] Hollingsworth SA, Karplus PA. A fresh look at the ramachandran plot and the occurrence of standard structures in proteins. Biomol Concepts, 2010, 1(3/4): 271–283.
[15] Mahdieh NR, Khajeh K, Mahdi A, et al. Enhancement of thermal stability of chondroitinase ABC I by site-directed mutagenesis: an insight from ramachandran plot. BBA-Proteins Proteom, 2013, 1834(2): 479–486.
[16] Fan EY. Cloning and expression of glutamate decarboxylase gene from Lactobacillus brevis[D]. Hangzhou: Zhejiang University, 2010 (in Chinese).范恩宇. 短乳杆菌谷氨酸脱羧酶基因的克隆与表达[D]. 杭州: 浙江大学, 2010.
[17] Huang J, Mei LH, Sheng Q, et al. Purification and characterization of glutamate decarboxylase of Lactobacillus brevis CGMCC 1306 isolated from fresh milk. Chin J Chem Eng, 2007, 15(2): 157–161.
[18] Yu K. Improved glutamate decarboxylase by computer-aided enzyme design[D]. Hangzhou: Zhejiang University, 2013 (in Chinese).郁凯. 以计算机辅助分子设计方法改造谷氨酸脱羧酶的研究[D]. 杭州: 浙江大学, 2013.
[19] Tina KG, Bhadra R, Srinivasan N. PIC: protein interactions calculator. Nucleic Acids Res, 2007, 35(2): W473-W475.
[20] Kellis JT, Nyberg K, Sali D, et al. Contribution of hydrophobic interactions to protein stability. Nature, 1988, 333(6175): 784–786.
[21] Pace CN, Fu H, Fryar K, et al. Contribution of hydrophobic interactions to protein stability. J Mol Biol, 2011, 408(3): 514–528.
(本文责编 陈宏宇)
Enhancing glutamate decarboxylase activity by site-directed mutagenesis: an insight from Ramachandran plot
Piyu Ke1, Jun Huang2, Sheng Hu3, Weirui Zhao1, Changjiang Lü1, Kai Yu1, Yinlin Lei3, Jinbo Wang3, and Lehe Mei1,3
1 College of Chemical and Biological Engineering, Zhejiang University, Hangzhou 310027, Zhejiang, China
2 School of Biological and Chemical Engineering, Zhejiang University of Science and Technology, Hangzhou 310023, Zhejiang, China
3 School of Biotechnology and Chemical Engineering, Ningbo Institute of Technology, Zhejiang University, Ningbo 315100, Zhejiang, China
Abstract:Glutamate decarboxylase (GAD) can catalyze the decarboxylation of glutamate into γ-aminobutyrate (GABA) and is the only enzyme of GABA biosynthesis. Improving GAD activity and thermostability will be helpful for the highly efficient biosynthesis of GABA. According to the Ramachandran plot information of GAD1407 three-dimensional structure from Lactobacillus brevis CGMCC No. 1306, we identified the unstable site K413 as the mutation target, constructed the mutant GAD by site-directed mutagenesis and measured the thermostability and activity of the wide type and mutant GAD. Mutant K413A led to a remarkably slower inactivation rate, and its half-life at 50 ℃ reached 105 min which was 2.1-fold higher than the wild type GAD1407. Moreover, mutant K413I exhibited 1.6-fold higher activity in comparison with the wide type GAD1407, although it had little improvement in thermostability of GAD. Ramachandran plot can be considered as a potential approach to increase GAD thermostability and activity.
Keywords:glutamate decarboxylase, protein engineering, γ- aminobutyric acid, Ramachandran plot
DOI:10.13345/j.cjb.150127
Corresponding author:Lehe Mei. Tel: +86-571-8795316; Fax: +86-571-87951982; E-mail: meilh@zju.edu.cn.
猜你喜欢
中国食品(2021年17期)2021-09-27
现代畜牧科技(2021年6期)2021-07-16
现代食品(2021年2期)2021-03-11
中国粮油学报(2019年4期)2019-07-12
现代盐化工(2018年1期)2018-03-29
中国酿造(2017年12期)2017-12-28
食品安全导刊(2017年24期)2017-09-16
磁共振成像(2015年5期)2015-12-23
饲料博览(2015年2期)2015-04-04
茶叶学报(2014年2期)2014-03-06
