
草鱼出血病分子流行病学及GCRV多样性研究
2016-06-23 03:01:40黄毅昌杨玉滔闫秀英简纪常吴灶和
安徽农业科学 2016年11期
关键词:多样性
黄毅昌,雷 燕,杨玉滔,闫秀英*,简纪常,吴灶和
(1.广东海洋大学水产学院,广东省水产经济动物病原生物学及流行病学重点实验室/水产经济动物病害控制广东普通高校重点实验室,广东湛江 524088;2.广州利洋水产科技股份有限公司,广东广州 510515)
草鱼出血病分子流行病学及GCRV多样性研究
黄毅昌1,雷 燕2,杨玉滔1,闫秀英1*,简纪常1,吴灶和1
(1.广东海洋大学水产学院,广东省水产经济动物病原生物学及流行病学重点实验室/水产经济动物病害控制广东普通高校重点实验室,广东湛江 524088;2.广州利洋水产科技股份有限公司,广东广州 510515)
摘要[目的]分析不同基因型GCRV分离株与患草鱼出血病草鱼症状的相关性。[方法]采用RT-PCR检测患病草鱼,应用生物信息学方法分析基因型内和基因型间不同分离株间的差异。[结果]对近3年来草鱼出血病分子流行病学分析发现,患草鱼出血病病鱼多为“肠炎型”症状,且检测发现其病原多属于GCRV基因型Ⅲ型。对GCRV的多样性分析表明,不同分离株同源蛋白的同源率高达93%以上,却被给予完全不同的名称,给GCRV多样性研究带来一定的困难;同一基因型不同分离株同源蛋白间的变异位点较少,且功能位点发生变异的更少;不同基因型不同分离株同源蛋白间的保守位点较少,且这些保守位点中只有极少数位点为功能位点;不同基因型分离株同源结构蛋白间存在较大的差异,且同源非结构蛋白间的差异更为显著。[结论]草鱼出血病不同临床症状可能与其病原基因型不同有关。
关键词草鱼呼肠孤病毒;草鱼出血病;分子流行病学;多样性
草鱼(Ctenopharyngodonidellus)是我国淡水养殖的主要经济鱼类之一,但草鱼出血病给草鱼养殖业造成了严重的经济损失[1]。根据患病草鱼出血部位的不同,草鱼出血病的症状可分为3大类型:“红肌肉型”、“红鳍红鳃盖型”和“肠炎型”[2]。迄今为止,对草鱼出血病及其病原草鱼呼肠孤病毒(Grass carp reovirus)的研究已取得很多突破性进展[3-6]。草鱼呼肠孤病毒是分节的dsRNA病毒,具有11个基因节段,且有些分离株间存在较大的差异[4]。不同的选择压影响着病毒的演化[7],目前已分离到多株GCRV分离株,发现已有的GCRV分为4种基因型,在我国分布有3种基因型[8-10]。
控制草鱼出血病的发生、减少草鱼出血病发生率、降低患草鱼出血病草鱼死亡率以及提高草鱼出血病疫苗的免疫效果是目前从事水产养殖业者关心的主要问题。对草鱼出血病及其病原的研究可为降低草鱼养殖业的损失奠定基础,但因为草鱼出血病引起的草鱼死亡率仍然较高,病原多样性是影响因素之一[5,11]。根据已有的研究和提交至GenBank数据库的基因序列及近年来草鱼出血病的症状分析表明,近几年发现的分离株多为以GCRV HZ08为代表的基因型Ⅲ型分离株,且草鱼出血病的症状多为“肠炎型”。因此,推测草鱼呼肠孤病毒的多样性可能与其引起的疾病症状相关,深入研究草鱼呼肠孤病毒的多样性对草鱼出血病的流行及防控治疗有着重要意义。广州利洋水产科技股份有限公司于2012~2015年对草鱼出血病分子流行病学进行了调查,笔者通过对GCRV基因组和蛋白多样性及蛋白特性进行分析,探讨GCRV多样性与草鱼出血病临床症状及GCRV致病机理的相关性,旨在为深入研究GCRV的致病机理、蛋白功能及草鱼出血病的防控等奠定基础。
1材料与方法
1.1草鱼出血病分子流行病学调查2012年4月至2015年6月,在我国各地草鱼养殖区采集患出血病的草鱼,检查并记录出血病的临床症状,同时应用RT-PCR方法对其病原GCRV进行检测,并对获得的目的片段进行测序。RT-PCR检测的GCRV目的基因为vp2基因,目的片段长度为413 bp,PCR所用引物为UP1(5′-GTTCCTGTCGTGGCTGGTATC-3′)和DW1(5′-GGTAGCTTAGAGTTCCTATCA-3′)、UP2(5′-GTCCCACTCACTGCCGGTATG-3′)和DW2(5′-GGGATTTTCGTGAAGGTTGTCT-3′)。PCR反应体系(25 μL)如下:ddH2O 16.5 μL、cDNA(约25 ng)2 μL、引物(10 μmol/L)各1 μL、dNTPs(2.5 mmol/L)2 μL、10×ExTaqBuffer 2.5 μL、ExTaq酶(1 U/μL)0.125 μL。PCR反应条件为:95 ℃ 5 min;95 ℃ 1 min,52 ℃ 1 min,72 ℃ 1 min,35个循环;72 ℃ 10 min。PCR反应结束后,用1%琼脂糖凝胶电泳检测PCR产物。将电泳产物纯化回收后与pMD18-T载体连接,转化至大肠杆菌 DH5α菌株,筛选阳性克隆并送至上海生工生物有限公司测序鉴定。
所获得的检测毒株序列与已知的GCRV序列进行比对,然后使用MEGA软件构建最大简约树(MP)、最大似然树(ML)和邻接树(NJ),Bootstrip值设为1 000。
1.2GCRV基因组多样性分析从GenBank数据库中下载已知的GCRV分离株基因组序列,应用DNAStar软件分析各分离株的同源性,进而分析GCRV基因组多样性。
1.3GCRV Ⅲ型基因型分离株蛋白的变异特性目前已知GCRV Ⅲ型基因型分离株基因组和基因序列最多,在已知的GCRV基因组中有9株GCRV分离株属于Ⅲ型基因型,1株分离株(GCRV-873)属于Ⅰ型基因型,1株分离株(GCRV 104)属于Ⅳ型基因型。为探讨GCRV的多样性与GCRV的进化,多样性包括基因型间的多样性和基因型内的多样性,同时为了深入探讨GCRV基因型和GCRV蛋白作用位点与疾病症状的关联,对已知的GCRV Ⅲ型基因型分离株(GCRV 106、GCRV JX02、GCRV 918、GCRV 794、GCRV 109、GCRV Henan988、GCRV HZ08和GCRV GD108)蛋白变异特性进行分析。使用DNAStar排列相关氨基酸序列,并统计相应同源蛋白的变异位点。
1.4GCRV HZ08蛋白相互作用基序分析使用iLIR软件(http://repeat.biol.ucy.ac.cy/iLIR/)对HZ08分离株的蛋白质氨基酸序列进行相互作用基序分析。
1.5GCRV HZ08蛋白糖基化位点、金属及二硫键结合位点分析使用NetOGlyc软件(http://www.cbs.dtu.dk/services/NetOGlyc/)分别对HZ08的各个蛋白质进行O-连接糖链的连接位点预测,得到上述蛋白质潜在的糖基化位点。使用METALDETECTER软件对上述蛋白质进行金属结合位点预测,得出其金属结合位点与二硫键结合位点。
1.6GCRV 873 VP1、VP3、VP5、VP6蛋白相互作用位点分析利用蛋白质三维结构数据库Protein Data Base获取GCRV-873的VP1、VP3、VP5、VP6蛋白的三级结构数据,并结合基于序列特征的不同残基溶剂可及性的SPPIDER方法,使用SPPIDER在线数据库(http://sppider.cchmc.org/)对上述蛋白质进行蛋白互作位点预测。
1.7不同基因型代表株VP1、VP3、VP5、VP6蛋白的保守位点分析以GCRV 873、GCRV HZ08和GCRV 104分别为基因型Ⅰ型、Ⅲ和Ⅳ型的代表株,使用DNAStar排列相关的氨基酸序列,分析此3株分离株蛋白的保守位点。
1.8GCRV 873 VP1、VP3、VP5、VP6蛋白的3D结构预测使用蛋白质数据库(Protein Data Bank)查询到的GCRV 873分离株的VP1、VP3、VP5和VP6蛋白的PDB文件[12],预测其3D结构。
2结果与分析
2.1草鱼出血病分子流行病学调查情况广州利洋水产科技股份有限公司水产动物疾病研究所于2012年4月至2015年6月从我国各地草鱼养殖区采集疑似患出血病的草鱼440例,检测结果表明其中288例草鱼出血病病毒呈阳性,此288例分布于山东、河南、湖北、湖南、天津、河北、广东、福建、山西等省份。检测出草鱼出血病病毒阳性的草鱼都具有肠道发红的症状,偶尔具有红肌肉的症状,未见到红鳍红鳃盖的症状。利用PCR方法检测到目的片段并进行测序,序列比对结果表明患出血病草鱼均检测出GCRV。同时,对检测到的毒株进行亲缘关系分析,使用MEGA所构建多种系统树的拓扑结构基本一致。从图1可以看出,所检测的毒株(GCRV-Tianjin、GCRV-Guangdong、GCRV-Hubei、GCRV-Qingyu)与GCRV HZ08亲缘关系最近。

注:括号内为已知GCRV分离株vp2基因的GenBank编码。数字为置信度。Note:GenBank codes of the vp2 genes in GCRV isolates are in brackets.Numbers are the confidence.图1 部分检测GCRV分离株的系统发生分析Fig.1 Analysis on the phylogency of some detected GCRV isolates
2.2GCRV基因组多样性分析从GenBank数据库中下载了GCRV 873、AGCRV、GCRV 106、GCRV Henan988、GCRV Hunan794、GCRV JX02、GCRV HZ08、GCRV 918、GCRV GD108、GCRV 109和GCRV 104基因组序列(表1),基因组序列GenBank编码分别为KC201166~KC201187、AF403390~AF40 3397、GQ896334~GQ896337、GU350742~GU350748、HQ231198~HQ231208、KC238676~KC238686、KC847320~KC847330、KF712475~KF712485、KM880065~KM 880073、NC_010584~NC_010594、JQ042805~JQ042808、AF260511~AF2605113、HQ018818和JN967629~JN967639。Ⅰ型基因型代表株GCRV 873、Ⅲ型基因型代表株GCRV HZ08和Ⅳ型基因型代表株GCRV 104相应蛋白相似性分析见表2。
2.3GCRV Ⅲ型基因型分离株蛋白的变异特性分析GCRV 106、GCRV JX02、GCRV 918、GCRV 794、GCRV 109、GCRV Henan988、GCRV HZ08和GCRV GD108编码的蛋白变异特性。从图2可以看出,GCRV基因组片段1编码的VP1间的同源率为99.1%~99.8%,变异位点有39个氨基酸位点;片段2编码的VP2间的同源率为98.0%~99.8%,变异位点有60个氨基酸位点;片段3编码的VP3间的同源率为97.2%~99.0%,变异位点55个个氨基酸位点;片段4编码的NS1或NS79间的同源率为97.9%~99.7%,变异位点有21个个氨基酸位点;片段5编码的VP5间的同源率为97.8%~99.4%,变异位点有31个氨基酸位点;片段6编码的VP4间的同源率为98.2%~100%,变异位点有22个氨基酸位点;片段7编码的NS4或VP56的同源率为93.4%~99.8%,变异位点有44个氨基酸位点;片段8编码的VP6间的同源率为97.1%~100%,变异位点有28个氨基酸位点;片段9编码的NS2间的同源率为97.5%~99.4%,变异位点有25个氨基酸位点;片段10编码的VP38或NS38间的同源率为99.1%~100%,变异位点有5个氨基酸位点;片段11编码的VP35或VP11间的同源率为97.4%~100%,变异位点有11个氨基酸位点。
注:*表示此行数据为该片段长度(bp)/基因长度(bp)/编码蛋白及其长度(aa)。
Note:* stands for that data in the line is the segment length(bp)/the gene length(bp)/the encoded proteins and their length(aa).
表2Ⅰ型基因型GCRV 873、Ⅲ型基因型GCRV HZ08和Ⅳ型基因型GCRV 104相应蛋白的相似性分析
Table 2Similarity analysis of homologous proteins in GCRV 873 attibuted to genotype Ⅰ,in GCRV HZ08 attibuted to genotype Ⅲ and in GCRV 104 attibuted to genotype Ⅳ
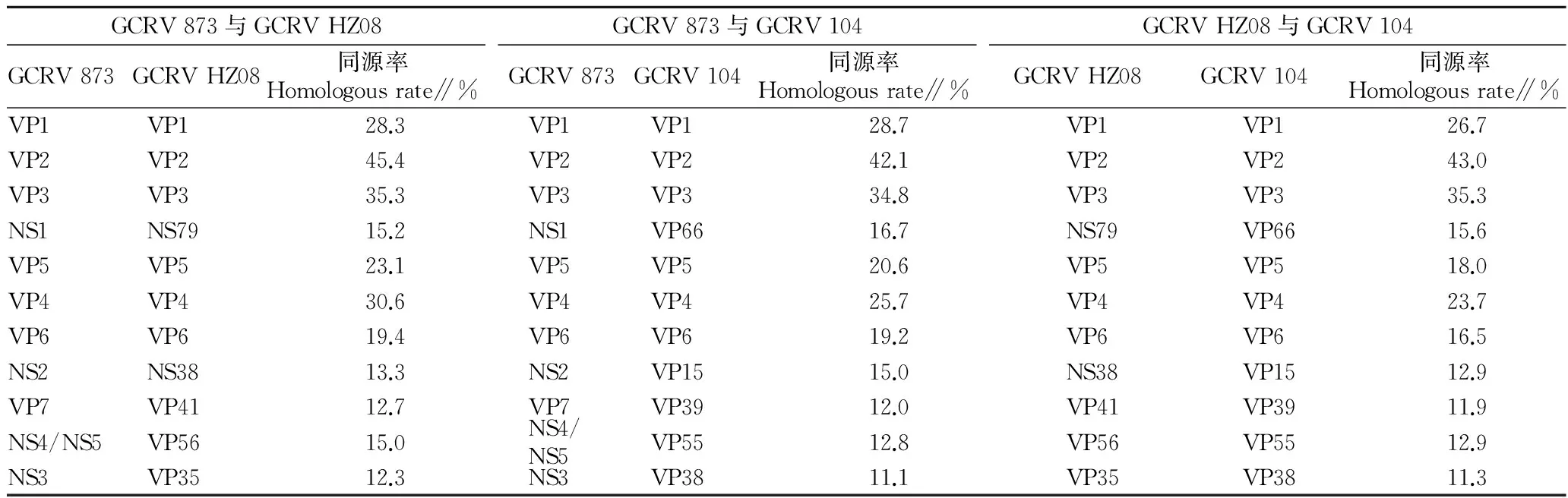
GCRV873与GCRVHZ08GCRV873GCRVHZ08同源率Homologousrate∥%GCRV873与GCRV104GCRV873GCRV104同源率Homologousrate∥%GCRVHZ08与GCRV104GCRVHZ08GCRV104同源率Homologousrate∥%VP1VP128.3VP1VP128.7VP1VP126.7VP2VP245.4VP2VP242.1VP2VP243.0VP3VP335.3VP3VP334.8VP3VP335.3NS1NS7915.2NS1VP6616.7NS79VP6615.6VP5VP523.1VP5VP520.6VP5VP518.0VP4VP430.6VP4VP425.7VP4VP423.7VP6VP619.4VP6VP619.2VP6VP616.5NS2NS3813.3NS2VP1515.0NS38VP1512.9VP7VP4112.7VP7VP3912.0VP41VP3911.9NS4/NS5VP5615.0NS4/NS5VP5512.8VP56VP5512.9NS3VP3512.3NS3VP3811.1VP35VP3811.3



图2 GCRV蛋白特性分析Fig.2 Analysis on the characterizations of GCRV proteins
2.4GCRV HZ08蛋白互作基序分析以GCRV HZ08为代表,分析其蛋白互作基序。从图2可以看出,VP1、VP2、VP3、NS79、VP5、VP4、VP56、VP41、VP6、NS38和VP35分别有168、156、120、66、90、60、42、36、78、24和30个氨基酸位点位于蛋白互作基序。
2.5GCRV HZ08蛋白糖基化位点、金属及二硫键结合位点分析通过预测发现VP1、VP2、VP3、VP4、VP5、VP6、NS79、VP35、NS38、VP41和VP56潜在的糖基化位点为分别有15、21、37、11、6、7、36、6、6、14、4个氨基酸位点(图2)。VP1、VP2、VP3、VP4、VP5、VP6、VP35、NS38、VP56、NS79和VP41分别含有39、49、40、10、30、16、21、16、13、37和23个金属结合位点(图1),其中VP1、VP2、VP3、VP6、VP35和NS38还分别含有2个二硫键结合位点(图2)。
2.6GCRV 873 VP1、VP3、VP5、VP6相互作用位点分析因为在PDB数据库中只登录有GCRV 873 VP1、VP3、VP5和VP6的PDB文件,因此只对此4个蛋白进行了蛋白互作位点分析。VP1、VP3、VP5和VP6的蛋白互作位点分别有243、273、136和114个氨基酸位点(图2)。
2.7不同基因型代表株VP1、VP3、VP5和VP6蛋白的保守位点分析基因型Ⅰ型、Ⅲ型和Ⅳ型基因型代表株GCRV 873、GCRV HZ08和GCRV 104分离株蛋白的保守位点如下:VP1、VP3、VP5和VP6间的保守位点分别有227、269、77和18个氨基酸位点(图2)。
2.8GCRV 873 VP1、VP3、VP5、VP6蛋白的3D结构预测应用DeepView预测GCRV 873 VP1、VP3、VP5、VP6蛋白的3D结构如图3所示。
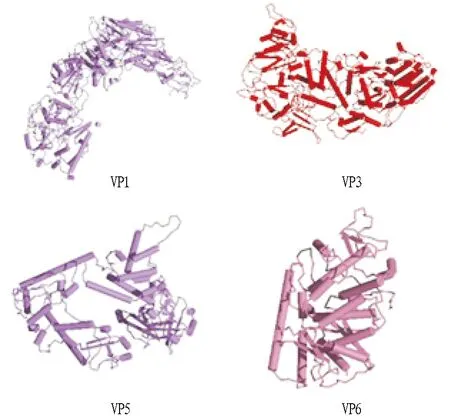
图3 GCRV 873 VP1、VP3、VP5、VP6蛋白的三级结构Fig.3 Tertiary structures of the proteins VP1,VP3,VP5 and VP6 in GCRV 873
3讨论与结论
对草鱼出血病分子流行病学进行调查及对GCRV多样性进行分析,有助于草鱼出血病的防治及GCRV致病机理的阐释。该试验结果表明在不同基因型分离株间片段1、2、3、5、6、9编码的结构蛋白VP1、VP2、VP3、VP4、VP5和VP6比较保守,但有些编码蛋白的基因所在片段不同。GCRV 873和AGCRV片段8编码VP6蛋白,而GCRV 106、GCRV Henan988、GCRV Hunan794、GCRV JX02、GCRV HZ08、GCRV 918、GCRV GD108、GCRV 109片段9编码VP6蛋白。非结构蛋白与结构蛋白相比变异较大,但同一基因型分离株间蛋白的同源性很高[13]。此外,即使上述蛋白同源性高达93%以上,不同分离株同源蛋白却给予不同的名称,如NS1与NS79、VP7与VP41、NS2与NS38、VP38、NS3与VP35等,这给GCRV多样性分析带来了一定的困难,由此建议同源性高的蛋白采用相同的名称给予命名。
在基因型Ⅲ型分离株间,蛋白间的变异位点分析表明,VP1蛋白39个变异位点中有9个氨基酸位点预测为蛋白互作位点,即73、130、378、379、445、654、905、1 088和1 120 aa。VP1的糖基化位点未发生变异,在金属结合位点中只有378 aa发生变异。在预测的VP1蛋白互作基序中,25、290、1 153、1 165和1 212 aa发生变异。VP2的50个变异位点中,2、279、315、564、765、946、1 062和1 250 aa位于VP2的互作基序,177和1 028 aa为糖基化位点,563 aa为金属结合位点。VP3的55个变异位点中,142、147、291、295和299 aa位于互作基序,57和103 aa为潜在的糖基化位点,4和884 aa为金属结合位点,还有17个位点为互作位点。NS1/NS79的21个变异位点中,401 aa位于互作基序,418 aa为潜在的糖基化位点。NS38/VP38的糖基化位点、金属结合位点、互作基序和蛋白互作位点未发生变异。VP4的22个变异位点中,70和418 aa位于互作基序,349 aa为金属结合位点。NS4/VP35的44个变异位点中,152、275、287、290和291 aa位于互作基序。VP5的31个变异位点中,554、699和715 aa位于互作基序,608 aa为糖基化位点,512 aa为金属结合位点,且41、238、421、554和618 aa为蛋白互作位点。VP6的28个变异位点中,111和302 aa位于互作基序,101和318 aa为潜在的糖基化位点,48、75、116和178 aa为金属结合位点,116 aa为二硫键结合位点。VP7/VP41的20个变异位点中,9、336和337 aa位于互作基序,69 aa为糖基化位点,155和
199 aa为金属结合位点。VP35/VP11/NS3/NS26的11个变异位点中,195 aa位于互作基序,167 aa为金属结合位点。由此可见,同一基因型分离株蛋白间的变异位点较少,且功能位点发生变异的更少。
Ⅰ型基因型GCRV 873、Ⅲ型基因型GCRV HZ08和Ⅳ型基因型GCRV 104比较保守的相应蛋白保守位点分析表明(以GCRV 873蛋白序列为模式序列进行相应氨基酸位点分析),VP1的227个保守位点中70、531、574、581和831氨基酸位点为潜在的糖基化位点,197、229、238和525氨基酸位点为金属结合位点,还有29个氨基酸位点同为预测的蛋白互作位点。VP3的269个保守位点中有29个氨基酸位点位于蛋白互作基序,609氨基酸位点为糖基化位点,119、122、135、140、410、617、836和874氨基酸位点为金属结合位点,119和122氨基酸位点同为二硫键结合位点,此外还有31个位点为蛋白互作位点。VP577个保守位点中有8个氨基酸位点位于互作基序,522氨基酸位点为金属结合位点,还有13个位点为蛋白互作位点。VP6的18个保守位点中,280氨基酸位点位于互作基序,111、165、169和179氨基酸位点为蛋白互作位点。由此可见,不同基因型分离株蛋白间的保守位点较少,且其中少数保守位点为功能位点,多数功能位点发生了变异。
结构蛋白是病毒颗粒的组成部分,非结构蛋白虽然不是病毒颗粒的组成部分,但在病毒感染和复制等过程中对病毒增殖起着极其重要的作用。目前的研究已表明病毒非结构蛋白参与调控病毒增殖、复制、感染致病等生命过程[14-15]。例如,禽呼肠孤病毒(Avian reovirus,ARV)的非结构蛋白δNS在ARV复制的起始阶段起着重要作用,非结构蛋白μNS在病毒装配中起着关键作用,且δNS还可提高或补充μNS的活性[13,16]。
不仅草鱼呼肠孤病毒不同基因型分离株结构蛋白间存在着比较大的差异,而且非结构蛋白间的变异更加显著,据此推测草鱼出血病不同症状可能与其病原分属不同基因型有关。GCRV病毒蛋白及宿主蛋白参与病毒复制的作用机制及GCRV致病机制仍未阐释清楚,草鱼出血病的症状到底与GCRV的基因型间的关系仍有待深入调查和研究。
参考文献
[1] 毛树坚,邵建忠,杭绮,等.草鱼出血病的病原研究[J].水产学报,1989,13(1):1-4.
[2] 王方华,李安兴.草鱼病毒性出血病研究进展[J].南方水产,2006,2(3):66-71.
[3] 殷亮,王庆,曾伟伟,等.水生呼肠孤病毒基因组研究进展[J].动物医学进展,2013,34(1):80-85.
[4] 李军,王铁辉,陆仁后,等.草鱼出血病病毒的研究进展[J].海洋与湖沼,1999,78(4):445-453.
[5] 张盛权.注射草鱼出血病疫苗免疫效果的观察报告[J].中国水产,2006,7(2):85-86.
[6] 郑树城,谢吉国,王雅,等.草鱼呼肠孤病毒096vp7基因生物信息学分析及其酵母表达载体的构建[J].广东海洋大学学报,2014,34(6):31-37.
[7] 崔治中.病毒准种多样性及其在免疫选择压作用下的演变[J].生命科学,2013,25(9):843-852.
[8] YAN X Y,XIONG L F,WANG Y,et al.Phylogenetic analysis of newly isolated grass carp reovirus[J].Springer plus,2014,3(4):190-199.
[9] 李永刚,曾伟伟,王庆,等.草鱼呼肠孤病毒分子生物学研究进展[J].动物医学进展,2013,34(4):97-103.
[10] WANG Q,ZENG W,LIU C,et al.Complete genome sequence of a reovirus isolated from grass carp,indicating different genotypes of GCRV in China[J].Journal of virology,2012,86(22):12466.
[11] JIANG Y L.Hemorrhagic disease of grass carp:Status of outbreaks,diagnosis,surveillance,and research[J].The israeli journal of aquaculture-bamidgeh,2009,61(3):188-192.
[12] CHENG L,ZHU J,HUI W H,et al.Backbone model of an aquareovirus virion by cryo-electron microscopy and bioinformatics[J].J Mol Biol,2010,397(2):852-863.
[13] FERNANDO T O,JOSE M C,VIKRAM N V,et al.Avian reovirus nonstructural protein δNS forms viroplasm-like inclusions and recruits protein δNS to structures[J].Virology,2004,319(1):94-106.
[14] 张丹,冯国和.丙型肝炎病毒非结构蛋白与宿主细胞蛋白相互作用的研究进展[J].世界华人消化杂志,2011,19(2):161-169.
[15] 边葶苈,周继勇,廖敏.冠状病毒非结构蛋白的研究进展[J].中国动物传染病学报,2013,21(4):67-74.
[16] FERNANDO T O,MARCELO C S M,JOSE M C,et al.Avian reovirus morphogenesis occurs within viral factories and begins with the selective recruitment of δNS and γA to δNS inclusions[J].J Mol Biol,2004,341(2):361-374.
Molecular Epidemiology of Hemorrhage Disease of Grass Carp and the Diversity Research of GCRV
HUANG Yi-chang1, LEI Yan2, YANG Yu-tao1, YAN Xiu-ying1*et al
(1. Guangdong Key Laboratory of Pathogenic Biology and Epidemiology for Aquatic Economic Animals, and Key Laboratory of Control for Diseases of Aquatic Economic Animals of Guangdong Higher Education Institues, Fisheries College of Guangdong Ocean University, Guangdong Ocean University, Zhanjiang, Guangdong 524088; 2. Guangzhou Liyang Aqua-Technology Co. Ltd, Guangzhou, Guangdong 510515)
Abstract[Objective] The aim was to analyze the correlation between different genotypes GCRV isolates and symptoms of grass carp with hemorrhage disease. [Method] Using RT-PCR detection, the bioinformatics method was adopted to analyze differences of isolates within and between various genotypes. [Result] The molecular epidemiology of hemorrhage disease of grass carp in recent three years was analyzed. It was found that the symptom of grass carp with hemorrhage disease was “the enteritis type”, with the GCRV mostly attributed to genotype Ⅲ. It was indicated that the homologous rates of homologous proteins in diferrent isolates were more than 93%, but those proteins were named with the completely different names. It (highly homologous proteins with different names) brought some difficulties to the research of GCRV diversity. Variable sites were few among homologous proteins in different isolates attributed to the same genotype, and variable function sites were fewer in these variable sites. Conserved sites among homologous proteins in different isolates attributed to different genotypes were few, and the fewer sites among them were functional sites. There were great differences among homologously structural proteins in different isolates attributed to different genotype, and the variation of homologous non-structural proteins was more significant. [Conclusion] It is suggested that different clinical symptoms of hemorrhage disease of grass carp could be related to different pathogen genotypes.
Key wordsGrass carp reovirus (GCRV); Hemorrhage disease of grass carp; Molecular epidemiology; Diversity
基金项目国家重点基础研究发展计划项目(2009CB118704)。
作者简介黄毅昌(1993- ),男,广东佛山人,本科生,专业:水产养殖学。* 通讯作者,副教授,博士,硕士生导师,从事水产动物病害防控。
收稿日期2016-03-12
中图分类号S 941
文献标识码A
文章编号0517-6611(2016)11-120-06
猜你喜欢
小学教学参考(语文)(2016年12期)2017-01-04 06:02:23
未来英才(2016年20期)2017-01-03 09:55:11
理论导刊(2016年12期)2016-12-27 17:37:25
甘肃教育(2016年22期)2016-12-20 22:15:31
新教育时代·教师版(2016年26期)2016-12-06 11:52:19
求知导刊(2016年30期)2016-12-03 10:48:27
戏剧之家(2016年22期)2016-11-30 15:13:39
散文百家·下旬刊(2016年9期)2016-11-23 22:23:14
散文百家·下旬刊(2016年10期)2016-11-17 21:22:22
科技资讯(2016年19期)2016-11-15 10:39:12
