
SET蛋白对精原细胞株GC-1 spg增殖和凋亡的影响
2016-06-07 01:51:23朱倩徐文丹张蓓许波群高超高莉刘嘉茵崔毓桂
生殖医学杂志 2016年5期
朱倩,徐文丹,张蓓,许波群,高超,高莉,刘嘉茵,崔毓桂*
(1. 南京医科大学第一附属医院生殖医学科,南京 210029;2. 南京医科大学第二附属医院妇科,南京 210036)
·实验研究·
SET蛋白对精原细胞株GC-1 spg增殖和凋亡的影响
朱倩1,徐文丹1,张蓓1,许波群2,高超1,高莉1,刘嘉茵1,崔毓桂1*
(1. 南京医科大学第一附属医院生殖医学科,南京210029;2. 南京医科大学第二附属医院妇科,南京210036)
【摘要】目的探讨SET蛋白对精原细胞株GC-1 spg增殖和凋亡的影响。方法使用含10%胎牛血清的DMEM高糖培养基在37℃、5%CO2条件下培养GC-1 spg细胞,分对照组(转染无关序列腺病毒)和实验组[转染SET干涉腺病毒(AdH1-siRNA/SET)];GC-1 spg接种于96孔板或者6孔板,转染腺病毒48 h、72 h后收集细胞,免疫荧光和共聚焦激光扫描显微镜检测SET蛋白在GC-1 spg细胞中的表达与定位;提取细胞总蛋白,Western Blot检测转染前后SET蛋白的表达;细胞计数试剂盒-8(CCK8)和5-溴脱氧尿嘧啶核苷(BrdU)标记法检测细胞的数目和增殖情况;流式细胞仪检测细胞凋亡的变化。结果SET蛋白表达于GC-1 spg细胞的胞核和胞浆中,且胞浆分布多于胞核;转染SET干涉腺病毒后,干涉组GC-1 spg细胞中的SET蛋白相对表达量为(0.217±0.044)显著低于对照组的(0.629±0.170)(P<0.05),同时GC-1 spg细胞的数目减少,增殖减慢,且细胞凋亡率显著增加[干涉组(21.663±1.287)%,对照组(8.813±0.671)%](P均<0.01)。结论SET蛋白在精原细胞GC-1 spg胞核和胞浆中均有表达,且胞浆分布多于胞核;GC-1 spg细胞中SET蛋白表达降低,能够抑制细胞增殖、促进细胞凋亡,提示SET蛋白可能参与调节精子发生的过程。
【关键词】SET蛋白;精子发生;精原细胞;细胞增殖;细胞凋亡
Objective: To investigate the effects of SET protein on the proliferation and apoptosis of spermatogonia cell line GC-1 spg.
Methods: GC-1 spg cells were cultured in DMEM/HIGH GLUCOSE with 10% fetal bovine serum in 5% CO2at 37℃. The cells in control group were transfected with AdH1-siRNA/NS,and the cells in experimental group were transfected with AdH1-siRNA/SET. The cells were seeded in 96-well plate or 6-well plate and treated with adenovirus for 48 or 72 hours,and then the cells were collected. The expression and cellular location of SET protein were assessed by immunofluorescence and confocal laser scanning microscopy. The total proteins were extracted,and the expressions of SET protein before and after transfection were detected by Western blotting. Cell number and proliferation were tested by CCK8 assay and BrdU incorporation assay,while cell apoptosis was measured by flow cytometry.
Results: It was found that SET protein was expressed both in cytoplasm and nucleus of GC-1 spg cells,and it was mainly expressed in cytoplasm. The expression of SET protein in the GC-1 spg cells transfected with AdH1-siRNA/SET [(0.217±0.044)vs.(0.629±0.170)] was significantly lowered than that in control group(P<0.05);while the apoptosis index was significantly increased [(21.663±1.287)vs.(8.813±0.671)%] (P<0.01),and the cell number and the proliferation rate were decreased (P<0.01).
Conclusions: SET protein was expressed both in cytoplasm and nucleus of GC-1 spg cells,and it was mainly expressed in cytoplasm. Knockdown of SET protein inhibits proliferation,and promotes apoptosis of GC-1 spg cells,which suggests that SET protein may participate in the regulation of spermatogenesis.
Key words:SET protein;Spermatogenesis;Spermatogonia cells;Cell proliferation;Cell apoptosis
(J Reprod Med 2016,25(5):449-454)
不育症已成为影响人类健康与社会发展的一个全球性医学和社会问题,全球约15%的育龄夫妇存在不育问题,在引起不育症的因素中男性因素占50%左右[1]。引起男性不育的原因有很多,其中精子发生障碍是男性不育常见的原因之一,占所有男性不育原因的20%~25%[1],临床表现为少、弱、畸形精子症。诸多激素、细胞因子调控精子发生过程,其中可能也包括SET蛋白[2]。SET基因是原癌基因,定位于染色体9q34着丝粒的c-abl。SET蛋白是细胞内多任务因子,参与调控包括细胞周期、细胞增殖凋亡、DNA修复、基因转录以及表观遗传等多个生物过程[3-4]。目前关于SET蛋白在生殖系统中的作用研究相对较少。本实验室前期研究发现,SET蛋白表达于卵巢中的卵泡膜细胞、卵母细胞以及睾丸中的间质细胞、精原细胞和精母细胞[2,5]。已有研究证实SET参与调节卵母细胞的发育[6-7],但是SET蛋白在精子发生中的作用及其机制鲜有报道。本研究采用SET干涉腺病毒转染体外培养的精原细胞株GC-1 spg,观察SET蛋白对GC-1 spg细胞增殖和凋亡的影响。
材料和方法
一、材料与试剂
小鼠精原细胞株GC-1 spg(南京医科大学生殖医学国家重点实验室惠赠),DMEM高糖培养基(HyClone,美国),胎牛血清(Gibco,美国),兔SET抗体(Santa Cruze,美国),兔beta-actin抗体(abcam,美国),二喹啉甲酸(BCA)蛋白浓度测定试剂盒和细胞组织快速裂解液RIPA(上海碧云天),辣根过氧化酶(HRP)标记山羊抗兔IgG和二氨基联苯胺(DAB)显色液(北京中杉金桥),细胞计数试剂盒-8(CCK-8)检测试剂盒(北方生物研究所),5-溴脱氧尿嘧啶核苷(BrdU)ELISA试剂盒(Roche,美国),四甲基异硫氰酸罗丹明(TRITC)标记的羊抗兔二抗(IgG-TRITC)(invitrogen,美国),酶标仪(Thermo,美国),共聚焦激光扫描显微镜(Nikon,日本),APC Annexin V/7-AAD流式凋亡试剂盒和流式细胞仪(BD Biosciences,美国)。SET干涉腺病毒载体由本课题组构建[8],应用于前期多项研究中[9-10]。
二、研究方法
1. 细胞分组与转染:使用含10%胎牛血清的DMEM高糖培养基常规培养GC-1 spg细胞。实验分为两组:对照组:转染无关序列腺病毒;干涉组(实验组):转染SET干涉腺病毒(AdH1-siRNA/SET)。GC-1 spg以5×104个/孔的密度接种于6孔板中,37℃、5%CO2条件下培养24 h,每孔转染2 μl的空载腺病毒或者干涉腺病毒载体,转染后48 h和72 h收集细胞进行后续试验。各实验重复3~5批次。
2. 蛋白质印迹(Western Blot)检测目标分子蛋白表达:转染腺病毒72 h后收集细胞提取总蛋白,BCA法测定蛋白浓度,12%的十二烷基磺酸钠-聚丙烯酰胺凝胶(SDS-PAGE)分离蛋白,湿转法转膜250 mA 2 h,5%脱脂牛奶37℃摇床封闭1~2 h,一抗(抗SET蛋白多克隆抗体1∶500,β-actin 1∶10 000作为内参)4℃过夜,用TBST洗膜3次,每次10 min,HRP标记的二抗(抗IgG 1∶1 000)37℃摇床孵育1 h,再用TBST洗膜3次,每次10 min,洗膜后使用增强化学发光(ECL)检测试剂盒(Millipore,美国)显影特异蛋白条带。采用天能Gel Image System分析软件进行灰度值分析,以目的蛋白与内参蛋白条带灰度值的比值均数作为各蛋白的相对表达量。
3. 荧光免疫和激光共聚焦检测SET蛋白在细胞中的定位:消过毒的小圆片置于24孔板中,GC-1 spg以2.5×104个/孔的密度接种于24孔板中,37℃、5%CO2条件下培养,细胞密度达到60%~70%时,PBS洗2次,每次5 min,4%多聚甲醛固定60 min,PBS洗4次,每次5 min,0.4%的triton-100细胞打孔5 min,PBS洗3次,室温下5%BSA湿盒内封闭1~2 h,1∶50稀释SET一抗,4℃湿盒内过夜,PBS洗膜,1∶500稀释TRITC荧光二抗,37℃湿盒内1 h,PBS洗膜3次,4,6-联脒-2-苯基吲哚(DAPI)染核1 min,PBS洗膜3次,Debico封片,在共聚焦显微镜下进行拍摄。阴性对照,是用无抗体活性的IgG替代SET一抗,其余操作与实验组一致。
4.CCK8检测细胞活率:GC-1 spg以1 250个/孔的密度接种于96孔板中,37℃、5%CO2条件下培养24 h,转染干涉腺病毒72 h后,96孔板每孔加10 μl的CCK8溶液,孵育2 h,用酶标仪测定450 nm处的吸光度值。
5.BrdU ELISA试剂盒检测细胞增殖:GC-1 spg以1 250个/孔的密度接种于96孔板中,37℃、5%CO2条件下培养24 h,转染干涉腺病毒24 h后,每孔加入10 μl的BrdU标记液,再37℃孵育24 h,去掉标记液,60℃放置1 h,每孔加入200 μl的FixDenat,室温孵育30 min,去掉FixDenat,拍板,每孔加入100 μl抗BrdU-POD工作液(1∶25稀释),室温孵育2 h,倒掉抗体工作液,用每孔200 μl洗液洗板3次,倒掉洗液,拍板使孔内没有残余,每孔加入100 μl的底物溶液,室温孵育30 min,读取370 nm处的光密度(OD)值,OD值越高,说明细胞增殖能力越强。
6. 流式细胞仪检测细胞凋亡:GC-1 spg以5×104个/孔的密度接种于6孔板中,37℃、5%CO2条件下培养24 h,转染干涉腺病毒72 h后消化、收集各组细胞,PBS洗涤后离心,PBS重悬后转移至流式管中1 500 rpm/min,离心5 min,倒掉PBS,加入400 μl的1×结合缓冲液及5 μl的APC Annexin V和5 μl的7-AAD,震荡混匀后,避光室温孵育15 min,流式细胞仪检测细胞凋亡。
三、统计学分析
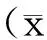
结果
一、SET蛋白在GC-1 spg细胞中的表达及定位
激光共聚焦结果显示,SET蛋白在GC-1 spg细胞核和胞浆中都有表达,且胞浆表达高于胞核(图1)。
A:阴性对照组DAPI染细胞核;B:阴性对照组TRITC标记的二抗荧光染色;C:阴性对照组细胞核和SET的合并荧光染色图;D:实验组DAPI染细胞核;E:实验组TRITC标记的SET一抗荧光染色;F:实验组细胞核和SET的合并荧光染色图。图1 共聚焦激光扫描显微镜检测SET蛋白在GC-1 spg细胞中的表达与定位(×600)
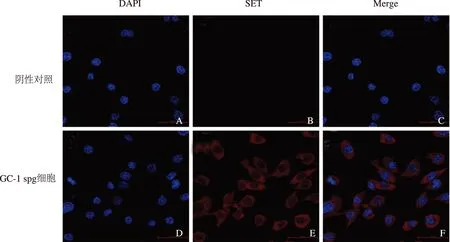
二、GC-1 spg转染SET干涉腺病毒后SET蛋白水平的变化
GC-1 spg转染SET干涉腺病毒72 h后,SET蛋白表达水平(0.217±0.044)较对照组(0.629±0.170)显著降低,干涉率为65%,差异具有统计学意义(P<0.05)(图2)。
三、SET蛋白干涉后GC-1 spg细胞数目和细胞增殖的变化
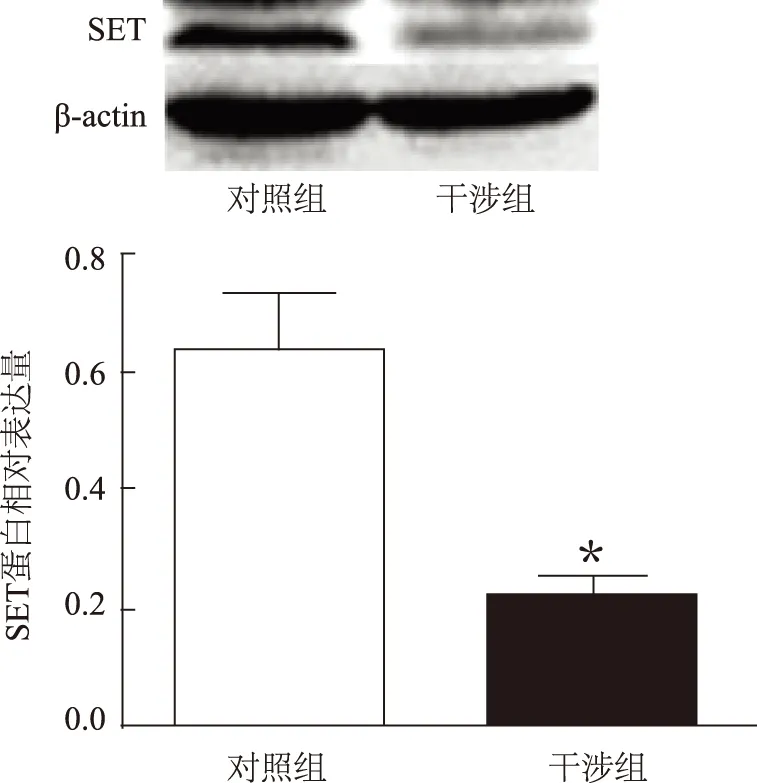
GC-1 spg转染SET干涉腺病毒72 h后,CCK8检测细胞数目结果表明,SET蛋白干涉组OD值为(0.281±0.068)较对照组的(1.193±0.005)明显降低,差异有统计学意义(P<0.01)(图3A)。
为了研究细胞数目减少是由于增殖减少还是凋亡增加引起的,GC-1 spg转染SET干涉腺病毒48 h后,采用BrdU ELISA法检测细胞增殖情况。结果表明,SET蛋白干涉组的OD值(0.080±0.004)较对照组(0.098±0.005)明显下降(P<0.01)(图3B)。
四、SET蛋白干涉后GC-1 spg细胞凋亡结果
GC-1spg转染SET干涉腺病毒72 h,流式细胞仪检测细胞凋亡。结果表明,干涉组细胞凋亡率[(21.663±1.287)%]较对照组[(8.813±0.671)%]显著升高(P<0.01)(图4)。
讨论
本文初步研究SET蛋白在精子发生中的作用及其作用机制,采用SET干涉腺病毒转染精原细胞株GC-1 spg,免疫荧光及共聚焦实验发现SET蛋白在GC-1 spg胞核和胞浆中都有表达,且胞浆多于胞核;利用Western Blot检查发现GC-1 spg转染SET干涉腺病毒后,SET蛋白的干涉效率为65%。免疫荧光可以进行细胞亚定位观察,并由此推导其潜在的生物学功能;结合Western Blot结果,探讨了SET蛋白在细胞增殖和细胞凋亡中的作用。诸多研究提示,随细胞状态改变,SET蛋白可分别定位于细胞核、细胞膜、细胞质及内质网。如在HeLa和HOS细胞、HeLa细胞的分裂间期,SET蛋白主要定位于细胞核内,亚细胞定位可能与其分子伴侣功能相关[11-12]。在大脑皮质神经元细胞,SET蛋白也主要定位于细胞核内,而当毒性诱导神经元凋亡时,SET蛋白在细胞核内减少而在细胞质内增多[13]。我们的实验未能显示RNAi后GC-1 spg细胞内SET蛋白免疫荧光亚定位的改变,可能与干涉效率65%不足以形成荧光强度、显微镜图像处理、所用荧光抗体与AdH1-siRNA/SET携带绿色荧光蛋白(GFP)相互干扰等原因有关。
本研究结果发现,降低GC-1 spg细胞的SET蛋白表达水平,能抑制GC-1 spg细胞的增殖,同时促进细胞凋亡。本课题组前期研究发现SET蛋白表达于精原细胞和精母细胞,以及睾丸间质细胞,提示SET蛋白一方面可能调节睾丸间质细胞雄激素合成,另一方面可能作用于生精细胞调节精子发生。研究发现SET过表达可以降低细胞的解毒作用,使细胞毒性物质和致癌物质积聚于细胞内,导致细胞死亡或肿瘤发生[14]。SET蛋白积聚在神经元胞浆中会使细胞对DNA损伤更加敏感,容易诱发细胞死亡[13]。不存在氧化应激时,聚集的SET蛋白在人源胚胎肾细胞HEK293T细胞的胞核和胞浆中均等分布,抑制细胞凋亡;而当细胞处于轻度氧化应激状态时,SET蛋白则主要聚集在胞浆中,促进细胞凋亡[15]。在氧化应激状态下,SET蛋白通过改变叉头框蛋白O1(FoxO1)的乙酰化状态正向调节细胞凋亡[16]。
SET蛋白通过多条通路调节细胞增殖,其中磷脂酰肌醇3-激酶/蛋白激酶B(PI3K/AKT)和细胞外调节蛋白激酶(ERK)信号通路是SET调节细胞增殖的重要通路。在非小细胞肺癌中,敲除SET蛋白,上调蛋白磷酸酶2A (PP2A)的活性,从而抑制AKT和ERK信号通路,抑制细胞增殖[17]。在前列腺癌中也发现SET蛋白通过PI3K/AKT信号通路调节细胞增殖[18]。在头颈部鳞状细胞癌细胞系HN12和Cal27中,SET蛋白通过ERK信号通路调节细胞增殖和存活[19]。有研究发现,SET蛋白通过不依赖caspase途径和依赖caspase途径来调节细胞凋亡[20-21]。刘雁峰等[22]研究发现,干涉SET蛋白能够通过激活外源性细胞凋亡通路引起人早幼粒细胞白血病维甲酸耐药细胞株NB4-R1凋亡。在HEK293T细胞中超表达的SET蛋白能够抑制caspase 9和caspase 3的活性,抑制内源性凋亡通路[15]。Cristobal等[23]也发现SET蛋白能够抑制PP2A激活的caspase凋亡途径。SET蛋白调节GC-1 spg细胞增殖和凋亡的机制,尚有待于进一步研究。
综上所述,SET蛋白表达于精原细胞GC-1 spg胞核和胞浆中,且胞浆多于胞核。降低SET蛋白表达水平,可以抑制GC-1 spg细胞增殖、促进细胞凋亡,表明SET蛋白可能在精子发生中发挥着重要的作用。深入研究SET蛋白在精子发生中的作用机制,有利于我们进一步了解男性少、弱、畸形精子症的发生,可能为男性不育症的治疗提供新线索。
注:与对照组比较,*P<0.05图2 Western Blot检测转染SET干涉腺病毒后GC-1 spg细胞中SET蛋白的表达情况
A:干涉SET蛋白表达后,GC-1 spg细胞数目变化;B:干涉SET蛋白表达后,GC-1 spg细胞增殖情况。注:与对照组比较,**P<0.01图3 干涉SET蛋白表达对GC-1 spg细胞数目和细胞增殖的影响
【参考文献】
[1]Wang Z,Jin B,Zhang X,et al. Yangjing capsule extract promotes proliferation of GC-1 Spg cells[J]. Evid Based Complement Alternat Med,2014,2014:640857.
[2]Dai XN,Liu S,Shao L,et al. Expression of the SET protein in testes of mice at different developmental stages[J]. Asian J Androl,2014,16:689-693.
[3]朱倩,崔毓桂. SET蛋白及其对雄激素合成的调节作用[J]. 国际生殖健康/计划生育杂志,2014,33:423-427.
[4]许波群,刘嘉茵,崔毓桂. SET/TAF-1β的研究进展[J]. 癌变畸变突变,2009,21:482-485.
[5]Boqun X,Xiaonan D,Yugui C,et al. Expression of SET protein in the ovaries of patients with polycystic ovary syndrome[J]. Int J Endocrinol,2013,2013:367956.
[6]Qi ST,Wang ZB,Ouyang YC,et al. Overexpression of SETbeta,a protein localizing to centromeres,causes precocious separation of chromatids during the first meiosis of mouse oocytes[J]. J Cell Sci,2013,126:1595-1603.
[7]Chambon JP,Touati SA,Berneau S,et al. The PP2A inhibitor I2PP2A is essential for sister chromatid segregation in oocyte meiosis II[J]. Curr Biol,2013,23:485-490.
[8]许波群,李瑛,薛凯,等. 人SET基因小分子干扰RNA重组腺病毒载体的构建与鉴定[J]. 医学研究生学报,2010,23:117-122.
[9]Xu B,Gao L,Cui Y,et al. SET protein up-regulated testosterone production in the cultured preantral follicles[J]. Reprod Biol Endocrinol,2013,11:9.
[10]Gao LL,Liu XQ,Xu BQ,et al. SET/PP2A system regulates androgen production in ovarian follicles in vitro[J]. Mol Cell Endocrinol,2013,374:108-116.
[11]Adachi Y,Pavlakis GN,Copeland TD. Identification and characterization of SET,a nuclear phosphoprotein encoded by the translocation break point in acute undifferentiated leukemia[J]. J Biol Chem,1994,269:2258-2262.
[12]Karetsou Z,Martic G,Sflomos G,et al. The histone chaperone SET/TAF-Ibeta interacts functionally with the CREB-binding protein[J]. Biochem Biophys Res Commun,2005,335:322-327.
[13]Qu D,Zhang Y,Ma J,et al. The nuclear localization of SET mediated by impalpha3/impbeta attenuates its cytosolic toxicity in neurons[J]. J Neurochem,2007,103:408-422.
[14]Almeida LO,Goto RN,Pestana CR,et al. SET overexpression decreases cell detoxification efficiency:ALDH2 and GSTP1 are downregulated,DDR is impaired and DNA damage accumulates[J]. FEBS J,2012,279:4615-4628.
[15]Leopoldino AM,Squarize CH,Garcia CB,et al. Accumulation of the SET protein in HEK293T cells and mild oxidative stress:cell survival or death signaling[J]. Mol Cell Biochem,2012,363:65-74.
[16]Chae YC,Kim KB,Kang JY,et al. Inhibition of FoxO1 acetylation by INHAT subunit SET/TAF-Ibeta induces p21 transcription[J]. FEBS Lett,2014,588:2867-2873.
[17]Liu H,Gu Y,Wang H,et al. Overexpression of PP2A inhibitor SET oncoprotein is associated with tumor progression and poor prognosis in human non-small cell lung cancer[J]. Oncotarget,2015,6:14913-14925.
[18]Farrell AS,Allen-Petersen B,Daniel CJ,et al. Targeting inhibitors of the tumor suppressor PP2A for the treatment of pancreatic cancer[J]. Mol Cancer Res,2014,12:924-939.
[19]Sobral LM,Sousa LO,Coletta RD,et al. Stable SET knockdown in head and neck squamous cell carcinoma promotes cell invasion and the mesenchymal-like phenotype in vitro,as well as necrosis,cisplatin sensitivity and lymph node metastasis in xenograft tumor models[J]. Mol Cancer,2014,13:32.
[20]Fan Z,Beresford PJ,Oh DY,et al. Tumor suppressor NM23-H1 is a granzyme A-activated DNase during CTL-mediated apoptosis,and the nucleosome assembly protein SET is its inhibitor[J]. Cell,2003,112:659-672.
[21]Chakravarti D,Hong R. SET-ting the stage for life and death[J]. Cell,2003,112:589-591.
[22]刘雁峰,贺鹏程,刘锋,等. RNA干扰I2PP2A基因表达对NB4-R1细胞增殖及凋亡的影响[J]. 中华血液学杂志,2014,35:732-736.
[23]Cristobal I,Rincon R,Manso R,et al. Deregulation of the PP2A inhibitor SET shows promising therapeutic implications and determines poor clinical outcome in patients with metastatic colorectal cancer[J]. Clin Cancer Res,2015,21:347-356.
[编辑:辛玲]
Effects of SET protein on proliferation and apoptosis of spermatogonia cell line GC-1 spg
ZHU Qian1,XU Wen-dan1,ZHANG Bei1,XU Bo-qun2,GAO Chao1,GAO Li1,LIU Jia-yin1,CUI Yu-gui1*
1.Clinical Center of Reproductive Medicine,the First Affiliated Hospital of Nanjing Medical University,Nanjing210029
2.The Second Affiliated Hospital of Nanjing Medical University,Nanjing210036
【Abstract】
DOI:10.3969/j.issn.1004-3845.2016.05.012
【收稿日期】2015-11-13;【修回日期】2016-01-14
【基金项目】国家自然科学基金(81370754,81170559,31301182);江苏省卫生厅项目(ZX201110,FXK201221)
【作者简介】朱倩,女,江苏镇江人,硕士研究生,生殖医学专业.(*通讯作者,Email:cuiygnj@njmu.edu.cn)
猜你喜欢
畜牧兽医学报(2023年7期)2023-07-31 08:32:22
右江医学(2021年9期)2021-10-22 05:15:28
昆明医科大学学报(2021年1期)2021-02-07 01:06:12
健康必读·下旬刊(2020年9期)2020-09-13 22:55:19
实用口腔医学杂志(2017年6期)2017-09-19 02:51:42
中南医学科学杂志(2017年1期)2017-01-12 20:25:49
右江医学(2016年4期)2017-01-05 16:26:48
医学信息(2016年29期)2016-11-28 09:54:46
癌变·畸变·突变(2016年5期)2016-08-22 05:55:20
课程教育研究·学法教法研究(2016年6期)2016-04-26 10:06:57
