
Optimal timing of autophagy occurrence induced by earle’s balanced salts solution in DLD-1, HCT-116, A2780, CHO, Hep G2 and SMMC7721 cancer cell lines*
2016-06-05 15:19,,,,
中山大学学报(自然科学版)(中英文) 2016年4期
, , , ,
(Department of Biochemistry, Zhongshan School of Medicine,Center for Stem Cell Biology and Tissue Engineering,Sun Yat-sen University, Guangzhou 510080, China)
Optimal timing of autophagy occurrence induced by earle’s balanced salts solution in DLD-1, HCT-116, A2780, CHO, Hep G2 and SMMC7721 cancer cell lines*
ZHANGWenliang,ZHANGMingming,LIUBin,SHIHongshun,WANGHaihe
(Department of Biochemistry, Zhongshan School of Medicine,Center for Stem Cell Biology and Tissue Engineering,Sun Yat-sen University, Guangzhou 510080, China)
Autophagy has emerged as a key effector in tumorgenesis, but its precise function remains elusive. Autophagy flux is too low to be detected in normal culture conditions. Earle’s balanced salts solution (EBSS) is often used to induce autophage with two distinct advantages compared with other inducers: one is only needed to considerate the induction time, rather than the drug concentration; the other is that it exerts only starvation pressure by specifically simulating amino acids deprivation to cells without other undistinguishable cytotoxicity. Given the tolerance threshold of outside stimulus differs in various types of tumor cells, in this study we carried out the autophage induction experiments with several time points in DLD-1 and HCT-116 colon cancer cell lines, A2780 and CHO ovarian cancer cell lines, Hep G2 and SMMC7721 hepatocellular carcinoma cell lines. Our western-blotting and Immunofluorescence staining results demonstrated that EBSS can efficiently induce autophagy in the assayed cells, precisely with 2 hours treatment in DLD-1, HCT-116, Hep G2 and CHO cells, 4 hours in A2780 and 8 hours in SMMC7721 cells, respectively. Our results provide a reference for autophagy induction with EBSS in cancer cells, and would be useful to shorten the time-consuming optimizing process.
autophage induction; EBSS; LC3; SQSTM1/p62; cancer cells
Autophagy is an evolutionarily highly conserved cellular homeostatic process whereby cells maintain their cytoplasmic biomass, organellar abundance and proper distribution, as well as remove potentially harmful protein aggregates and aged organelles[1]. Although the precise function of autophagy remains unclear, more and more evidences have shown that it is an important factor in tumorgenesis and cancer treatment[2]. The current consensus suggests that autophagy may have a dual role in cancer. In normal cells, autophagy functions to prevent the accumulation of damaged or aged organelles and aggregated proteins; by contrast, in cancer cells, autophagy is hijacked to serve as a cell survival mechanism that plays a critical role for established tumors[2]. In normal culture conditions, the detection of autophagy is relatively hard because of its low level[3]. Thus, to elaborate the precise role of autophagy in tumorgenesis and drug resistance, the autophagy activators and inhibitors need to be used to activate and suppress autophagy, respectively. Recently, earle’s balanced salts solution (EBSS) is a saline solution with physiological pH which is often used to induce autophagy[4-10]. At the same time, DLD-1 and HCT-116 as colon cancer[11-13], A2780 and CHO as ovarian cancer[13-15], Hep G2 and SMMC7721 as hepatocellular carcinoma[16-19]have been frequently used to elaborate the mechanism of autophagy in tumorgenesis and drug resistance. However, no detailed information of EBSS starvation duration for autophage induction in those cell lines has been reported yet. In this study, we performed western blotting and Immunofluorescence stainings in DLD-1, HCT-116, A2780, CHO, Hep G2 and SMMC7721 cell lines at several designed time points with EBSS starvation, hoping to find the optimal timing of autophagy occurrence in these tumor cell.
1 Materials and Methods
1.1 Cell culture and treatment
Human colorectal carcinoma cell line DLD-1 and HCT116(ATCC),CHO-K1 cells (ATCC), and human ovarian carcinoma cell line A2780 (ATCC) were maintained in DMEM basic medium(Gbico)supplemented with 10% fetal bovine serum and 1% antibiotics/antimycotics. Hepatocellular carcinoma cell line Hep G2 and SMMC7721 (ATCC) were maintained in DMEM high glucose medium (HyClone) supplemented with 10% fetal bovine serum and 1% antibiotics/antimycotics. All cells were maintained in a 5% CO2incubator at 37 ℃. EBSS was purchased from Life Technologies (Calcium, Magnesium and Phenol Red free, Gibco, 14155-063). Unless otherwise specified, reagents were used at the following final concentrations: chloroquine diphosphate salt (50 μmol/L, C6628, Sigma-Aldrich).
1.2 Immunofluorescence staining
Cells were fixed in 4% formaldehyde for 15 min, washed with cold 1 x PBS, permeabilized with 0.2% Triton X-100(BIOSHARP, Ameresco 0694) for 30 min and blocked with 5% BSA (Sigma-Aldrich, 05470) for 1 h at room temperature. Cells were then incubated with primary antibodies overnight at 4 ℃. Anti-LC3 antibody (3868, Cell Signaling Technology) was used at a dilution of 1∶200. After being washed five times with 1 x PBS, cells were incubated with secondary antibody (1∶500 dilution; Life Technologies, A21207) conjugated with appropriate fluorophore for 30 min at room temperature. Slides were mounted with Prolong Gold Antifade Reagent and analyzed immediately by using an LSM710 confocal microscope (Carl Zeiss AG). The averaged LC3 channel pixel intensity value within the cytoplasmic area per cell was determined by Photoshop (Adobe Systems) to represent the overall amount of LC3 puncta accumulation in each cell. Results were presented as mean ± SD.
1.3 Western blotting
The detailed immunoblotting procedures had been described previously[20]. LC3 (3868), SQSTM1/p62 (5114), Anti-rabbit HRP-labeled secondary antibody (7074) and anti-mouse HRP-labeled secondary antibody (7076S) were purchased from Cell Signaling Technology. GAPDH (CW0101A) was purchased from CW Biotech. The Actin (sc-8432) was purchased from Santa Cruz Biotechnology. Immunoreactive bands were visualized by using chemiluminescent substrate (Millipore, MA01821) and photographed on radiography film. Quantification of band intensity was done by Image J software.
1.4 Statistical analysis
All the statistical analyses were performed by GraphPad Prism version 5.00 software for windows (GraphPad Software, San Diego California USA, www.graphpad.com.).One-way ANOVA with Dunnett’s post test was performed for the comparison of means between groups.P< 0.05 was considered as significant in all tests.
2 Results
2.1 EBSS is an easy-to-use autophagy inducer
EBSS is a saline solution with physiological pH that is often used to induce autophagy[4-10]. We searched and summarized the putative reagents utilized for autophagy induction (Table 1). From the above Table, we found that EBSS as a saline solution have two distinct advantages, one is that comparing to other pharmic inducers, when EBSS is used as an autophage inducer, we only need to considerate the induction time, rather than the drug concentration; the other is that by specifically simulating amino acids deprivation, it just exerts starvation pressure to cells without arising other undistinguishable cell toxicities.
2.2 The optimal timing of autophagy induced by EBSS in cancer cells
Microtubule-associated protein 1 light chain 3 (LC3) is the most widely and reliably monitored autophagy-related protein[3]. A hallmark of autophagy initiation is the conversion of the cytosolic LC3 I into the membrane bound LC3 II that associates with autophagosome formation[21]. Upon the autophagy initiation,the ratio of LC3 II/LC3 I and LC3 II level increase because the LC3 II rapidly conversed from LC3 I is more than the LC3 II degraded by autophgolyosome. However, along with the autophagy induction, the ratio of LC3II/LC3 I and LC3 II level are obviously decreased because LC3 II as an autophage substrate is rapidly degraded in autophgolyosome. In addition to LC3, SQSTM1/p62 becomes incorporated into the completed autophagosome and is also significantly degraded in autolysosomes when autophagy is induced[3]. Collectively, the degradation of LC3 and SQSTM1/p62 can be used to monitor the autophagy induction.

Table 1 Reagents used for autophagy induction
2.2.1 In DLD-1 and HCT-116 colon cancer cell lines In colon cancer cells, western blotting results show that upon 1 h EBSS starvation, the ratio of LC3 II/LC3 I has been increased compared with the control group (under normal culture condition) in DLD-1 (Fig.1 A, green box, lane 2 and Fig.1 B) and HCT-116 (Fig.1 E, green box, lane 2 and Fig.1 F), respectively, which means autophagy is initiated by EBSS after one hour starvation. Then, with the starvation time extended to 2 h, the ratios of LC3 II/LC3 I, LC3 II/GAPDH and SQSTM1/p62/GAPDH have both been reduced significantly compared with the control group in DLD-1 ( Fig.1 A, red box, lane 3 and Fig.1 B) and HCT-116 (Fig.1 E, red box, lane 3 and Fig.1 F). In addition, we choose LC3 protein as a marker to trace autophagosome and perform Immunofluorescence (IF) staining. Chloroquine (CQ) is a lysosomal acidification inhibitor that blocks autophagosome related degradation. In colon cancer cell lines, we noticed that DLD-1 and HCT-116 cells treated with EBSS + CQ at 2 h (Fig.1 C, b’ and G, b’) contained more endogenous LC3 puncta per cell than DLD-1 and HCT-116 cells from respective control group (Fig.1 C, a’ and G, a’). Quantification of LC3 puncta intensity confirmed a significantly higher LC3 puncta frequency per cell in EBSS + CQ treated DLD-1 and HCT-116 cells at 2 h compared with normally cultured DLD-1 and HCT-116 cells, respectively (Fig.1 D and H;P<0.001). Collectively, our results suggest that autophage is efficiently induced by EBSS in DLD-1 and HCT-116 colon cancer cells after only 2 hours.
2.2.2 In A2780 and CHO ovarian cancer cell lines Noteworthily, upon 2 h EBSS starvation in A2780 cell line, the ratios of LC3 II/LC3 I and LC3 II/GAPDH have been obviously increased compared with the control group (Fig.2 A, green box, lane 2 and Fig.2 B). The increased ratios of LC3 II/LC3 I and LC3 II/GAPDH indicate that autophagy is initiated by EBSS starvation at 2 h in A2780. When the starvation reaches 4 h, the ratios of LC3 II/GAPDH and SQSTM1/p62/GAPDH are both obviously decreased compared with the control group in A2780 (Fig.2 A, red box, lane 3 and Fig.2 B). In addition, the IF results show that A2780 cells treated with EBSS + CQ at 4 h (Fig.2 C, b’) contained more endogenous LC3 puncta per cell than A2780 cells from control group (Fig.2 C, a’). Quantification of LC3 puncta intensity confirmed a significantly higher LC3 puncta frequency per cell in EBSS + CQ treated A2780 cells at 4 h compared with normally cultured A2780 cells, respectively (Fig.2 D;P<0.001 ). Our results suggest that EBSS can efficiently induce autophage in A2780 when the starvation time is extended to 4 h. However, in CHO cell line, upon 2 h EBSS starvation, the ratios of LC3II/LC3 I, LC3 II/GAPDH and SQSTM1/p62/GAPDH have been significantly reduced compared with the control group (Fig.2 E, red box, lane 2 and Fig.2 F). Those results demonstrate that the autophgy is induced in CHO by EBSS starvation after only 2 hours. In addition, the IF results showed that CHO cells treated with EBSS + CQ at 2 h (Fig.2 G, b’) contained more endogenous LC3 puncta per cell than CHO cells from control group (Fig.2 G, a’). Quantification of LC3 puncta intensity confirmed a significantly higher LC3 puncta frequency per cell in EBSS + CQ treated CHO cells at 2 h compared with normally cultured CHO cells, respectively (Fig.2 H;P<0.001 ). Thus, our results confirm that 2 h EBSS starvation can efficiently induce autophage in CHO cells.
2.2.3 In HepG2 and SMMC7721 hepatocarcinoma cell lines In HepG2 cell line, the western blotting result indicates that upon 1 h EBSS starvation, the ratios of LC3 II/LC3 I and LC3 II/GAPDH have been both increased compared with the control group (Fig.3 A, boxed lane 2 and Fig.3 B). The increased ratios of LC3 II/LC3 I and LC3 II/GAPDH also confirm that autophagy is initiated by 1 h EBSS starvation in Hep G2. With the starvation extended to 2 h, the ratios of LC3 II/GAPDH and SQSTM1/p62/GAPDH are both significantly reduced compared with the control group (Fig.3 A, boxed lane 3 and Fig.3 B). It should also be noted that 2 h EBSS + CQ treatment clearly increase the LC3 II level compared with the control group in Hep G2 (Fig.3 C, boxed lane 3). Thus, the above results suggest that the autophage is induced by only 2 h EBSS starvation in Hep G2. However, in SMMC7721 cell line, upon 4 h EBSS starvation, the LC3II/GAPDH and LC3 II / LC3 I are not reduced compared with the control group (Fig.3 D, boxed lane 2 and Fig.3 E). When the starvation is extended to 8 h in SMMC7721, the ratios of LC3 II/GAPDH and SQSTM1/p62/GAPDH rapidly decrease compared with the respective control group (Fig.3 D, boxed lane 3 and Fig.3 E). Similarly, we noticed that 8 h EBSS + CQ treatment significantly increased the LC3 II level compared with the control group in SMMC7721 (Fig.3 F, boxed lane 3). Our results show that the efficient induction time of SMMC7721 by EBSS starvation should be 8 h. Collectively, EBSS can also efficiently induce autophagy in hepatocarcinoma cells and it’s possible that the EBSS optimal induction time of autophagy is dependent on specific tumor cell types.
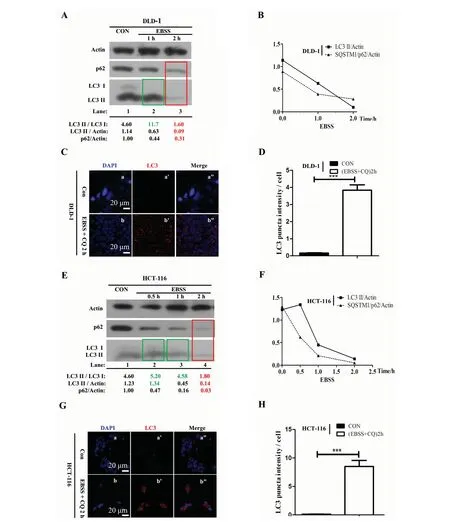
Fig.1 The optimal timing of autophagy induction by EBSS in DLD-1 and HCT-116 colon cancer cell lines

Fig.2 The optimal timing of autophagy induction by EBSS in A2780 and CHO ovarian cancer cell lines
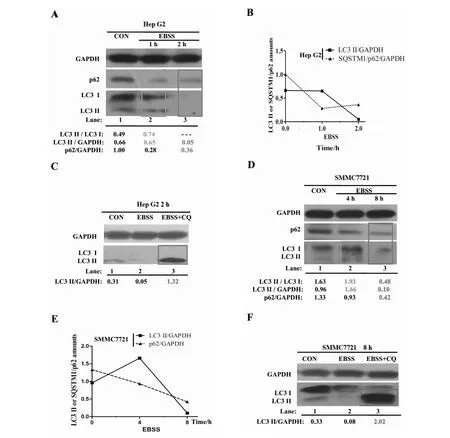
Fig.3 The optimal timing of autophagy induction by EBSS in Hep G2 and SMMC7721 hepatocarcinoma cell lines
3 Discussions
Autophagy has emerged as a key effecter in tumorgenesis and cancer treatment, although its precise function remains elusive[2].The DLD-1, HCT-116, A2780, CHO, Hep G2 and SMMC7721 cell lines are often used as models to elaborate the precise role of autophagy in tumorgenesis and drug resistance[11-19]. EBSS is often used as an autophage inducer in related studies[4-10], but its usage in DLD-1, HCT-116, A2780, CHO, Hep G2 and SMMC7721 for autophagy researches has not been explored yet. In this study, firstly we analyzed several common inducers of autophagy and pointed out that EBSS is easier to use in tumor cells than others, secondly we demonstrated that tumor cells can be efficiently induced by EBSS starvation in just a few hours. We also found that the optimal timing of autophagy occurrence varies in different cell types. Most importantly, we have obtained the optimal induction time of DLD-1, HCT-116, A2780, CHO, Hep G2, and SMMC7721 cells through western blotting and IF results acquired from several specific induction time points. To sum up, our results provide practical usage data of EBSS as an autophagy inducer in several cancer cell lines, which would possibly be useful for other researchers to optimize their autophagy experiments and spare the time-consuming process of exploring the proper inducer and finding the appropriate induction time.
[1] DERETIC V. Autophagosome and phagosome[J]. Methods in Molecular Biology, 2008, 445:1-10.
[2] ZHI X, ZHONG Q. Autophagy in cancer[J]. F1000prime Reports, 2015, 7: 18.
[3] KLIONSKY D J, ABDALLA F C, ABELIOVICH H, et al. Guidelines for the use and interpretation of assays for monitoring autophagy[J]. Autophagy, 2012, 8(4): 445-544.
[4] LUO T, FU J, XU A, et al. PSMD10/Gankyrin Induces Autophagy to Promote Tumor Progression through Cytoplasmic Interaction with ATG7 and Nuclear Transactivation of ATG7 Expression[J]. Autophagy, 2015,11(4):1-17.
[5] MAURO-LIZCANO M, ESTEBAN-MARTINEZ L, SECO E, et al. New method to assess mitophagy flux by flow cytometry[J]. Autophagy, 2015, 11(5): 833-843.
[6] MIN Y, XU W, LIU D, et al. Earle’s balanced salts solution and rapamycin differentially regulate the Bacillus Calmette-Guerin-induced maturation of human dendritic cells[J]. Acta Biochimica et Biophysica Sinica, 2013, 45(3): 162-169.
[7] NAMKOONG S, LEE K I, LEE J I, et al. The integral membrane protein ITM2A, a transcriptional target of PKA-CREB, regulates autophagic flux via interaction with the vacuolar ATPase[J]. Autophagy, 2015, 11(5):756-768.
[8] NI Z, GONG Y, DAI X, et al. AU4S: a novel synthetic peptide to measure the activity of ATG4 in living cells[J]. Autophagy, 2015, 11(2):403-415.
[9] RUI Y N, XU Z, CHEN Z, et al. The GST-BHMT assay reveals a distinct mechanism underlying proteasome inhibition-induced macroautophagy in mammalian cells[J]. Autophagy, 2015, 11(5):812-832.
[10] WANG Z, HU J, LI G, et al. PHF23 (plant homeodomain finger protein 23) negatively regulates cell autophagy by promoting ubiquitination and degradation of E3 ligase LRSAM1[J]. Autophagy, 2014, 10(12):2158-2170.
[11] CHOI J H, YOON J S, WON Y W, et al. Chloroquine enhances the chemotherapeutic activity of 5-fluorouracil in a colon cancer cell line via cell cycle alteration[J]. APMIS, 2012, 120(7):597-604.
[12] COKER-GURKAN A, ARISAN E D, OBAKAN P, et al. Inhibition of autophagy by 3-MA potentiates purvalanol-induced apoptosis in Bax deficient HCT 116 colon cancer cells[J]. Experimental Cell Research, 2014, 328(1):87-98.
[13] HUANG Y H, AL-AIDAROOS A Q, YUEN H F, et al. A role of autophagy in PTP4A3-driven cancer progression[J]. Autophagy, 2014, 10(10):1787-1800.
[14] BAO L J, JARAMILLO M C, ZHANG Z B, et al. Nrf2 induces cisplatin resistance through activation of autophagy in ovarian carcinoma[J]. International Journal of Clinical and Experimental Pathology, 2014, 7(4):1502-1513.
[15] FONG M Y, JIN S, RANE M, et al. Withaferin A synergizes the therapeutic effect of doxorubicin through ROS-mediated autophagy in ovarian cancer[J]. PloS One, 2012, 7(7):e42265.
[16] DING Z B, HUI B, SHI Y H, et al. Autophagy activation in hepatocellular carcinoma contributes to the tolerance of oxaliplatin via reactive oxygen species modulation[J]. Clinical Cancer Research, 2011, 17(19):6229-6238.
[17] GOU X, RU Q, ZHANG H, et al. HAb18G/CD147 inhibits starvation-induced autophagy in human hepatoma cell SMMC7721 with an involvement of Beclin 1 down-regulation[J]. Cancer Science, 2009, 100(5):837-843.
[18] TONG Y, HUANG H, PAN H. Inhibition of MEK/ERK activation attenuates autophagy and potentiates pemetrexed-induced activity against HepG2 hepatocellular carcinoma cells[J]. Biochemical and Biophysical Research Communications, 2015, 456(1):86-91.
[19] YUAN H, LI A J, MA S L, et al. Inhibition of autophagy signi fi cantly enhances combination therapy with sorafenib and HDAC inhibitors for human hepatoma cells[J]. World Journal of Gastroenterology, 2014, 20(17):4953-4962.
[20] RUHUA Z, SHUANGBING X, YUANZHONG W, et al. Construction and identification of hSSB1 retrovirus expressing vector and screening of stable transfected cells[J]. Acta Scientiarum Naturalium Universitatis Sun Yat-seni, 2012 (2):73-76,85.
[21] KABEYA Y, MIZUSHIMA N, UENO T, et al. LC3, a mammalian homologue of yeast Apg8p, is localized in autophagosome membranes after processing[J]. The EMBO Journal, 2000, 19(21):5720-5728.
[22] MARCILLA-ETXENIKE A, MARTIN M L, NOGUERA-SALVA M A, et al. 2-Hydroxyoleic acid induces ER stress and autophagy in various human glioma cell lines[J]. PloS One, 2012, 7(10):e48235.
[23] TSVETKOV A S, MILLER J, ARRASATE M, et al. A small-molecule scaffold induces autophagy in primary neurons and protects against toxicity in a Huntington disease model[J]. Proceedings of the National Academy of Sciences of the United States of America, 2010, 107(39):16982-16987.
[24] JOO J H, DORSEY F C, JOSHI A, et al. Hsp90-Cdc37 chaperone complex regulates Ulk1- and Atg13-mediated mitophagy[J]. Molecular Cell, 2011, 43(4):572-585.
[25] MAIURI M C, LE TOUMELIN G, CRIOLLO A, et al. Functional and physical interaction between Bcl-X(L) and a BH3-like domain in Beclin-1[J]. The EMBO Journal, 2007, 26(10):2527-2539.
[26] PEDRO J M, WEI Y, SICA V, et al. BAX and BAK1 are dispensable for ABT-737-induced dissociation of the BCL2-BECN1 complex and autophagy[J]. Autophagy, 2015,11(3):452-459.
[27] DECUYPERE J P, BULTYNCK G, PARYS J B. A dual role for Ca(2+) in autophagy regulation[J]. Cell Calcium, 2011, 50(3):242-250.
[28] YANG S Y, KIM N H, CHO Y S, et al. Convallatoxin, a dual inducer of autophagy and apoptosis, inhibits angiogenesis in vitro and in vivo[J]. PloS One, 2014, 9(3):e91094.
[29] KLIONSKY D J, ABELIOVICH H, AGOSTINIS P, et al. Guidelines for the use and interpretation of assays for monitoring autophagy in higher eukaryotes[J]. Autophagy, 2008, 4(2):151-175.
1)12∶00~14∶00随着太阳角度和高度的变化逐时刻模拟太阳日照,是个积分过程,时刻划分越细,运算次数越多,非常繁琐。为了简便起见,只计算3个时刻(即 12∶00 、 13∶00和14∶00)的日照情况,近似模拟该时间段内的阴影范围,判断建筑的遮挡情况,计算量大大简化[5]。
[30] GARCIA-MARTINEZ J M, MORAN J, CLARKE R G, et al. Ku-0063794 is a specific inhibitor of the mammalian target of rapamycin (mTOR)[J]. The Biochemical Journal, 2009, 421(1):29-42.
[31] SARKAR S, KRISHNA G, IMARISIO S, et al. A rational mechanism for combination treatment of Huntington's disease using lithium and rapamycin[J]. Human Molecular Genetics, 2008, 17(2):170-178.
[32] PEREIRA G J, HIRATA H, FIMIA G M, et al. Nicotinic acid adenine dinucleotide phosphate (NAADP) regulates autophagy in cultured astrocytes[J]. The Journal of Biological Chemistry, 2011, 286(32):27875-27881.
[33] LIU T J, KOUL D, LAFORTUNE T, et al. NVP-BEZ235, a novel dual phosphatidylinositol 3-kinase/mammalian target of rapamycin inhibitor, elicits multifaceted antitumor activities in human gliomas[J]. Molecular Cancer Therapeutics, 2009, 8(8):2204-2210.
[34] SERRA V, MARKMAN B, SCALTRITI M, et al. NVP-BEZ235, a dual PI3K/mTOR inhibitor, prevents PI3K signaling and inhibits the growth of cancer cells with activating PI3K mutations[J]. Cancer Research, 2008, 68(19):8022-8030.
[35] WONG V K, WU A G, WANG J R, et al. Neferine attenuates the protein level and toxicity of mutant huntingtin in PC-12 cells via induction of autophagy[J]. Molecules, 2015, 20(3):3496-3514.
[37] PUISSANT A, AUBERGER P. AMPK- and p62/SQSTM1-dependent autophagy mediate resveratrol-induced cell death in chronic myelogenous leukemia[J]. Autophagy, 2010, 6(5):655-657.
[38] VINGTDEUX V, GILIBERTO L, ZHAO H, et al. AMP-activated protein kinase signaling activation by resveratrol modulates amyloid-beta peptide metabolism[J]. The Journal of Biological Chemistry, 2010, 285(12):9100-9113.
[39] WENG Z, LUO Y, YANG X, et al. Regorafenib impairs mitochondrial functions, activates AMP-activated protein kinase, induces autophagy, and causes rat hepatocyte necrosis[J]. Toxicology, 2015, 327:10-21.
[40] VINGTDEUX V, CHANDAKKAR P, ZHAO H, et al. Novel synthetic small-molecule activators of AMPK as enhancers of autophagy and amyloid-beta peptide degradation[J]. FASEB journal : official publication of the Federation of American Societies for Experimental Biology, 2011, 25(1):219-231.
[41] CASAREJOS M J, SOLANO R M, GOMEZ A, et al. The accumulation of neurotoxic proteins, induced by proteasome inhibition, is reverted by trehalose, an enhancer of autophagy, in human neuroblastoma cells[J]. Neurochemistry International, 2011, 58(4):512-520.
[42] SARKAR S, DAVIES J E, HUANG Z, et al. Trehalose, a novel mTOR-independent autophagy enhancer, accelerates the clearance of mutant huntingtin and alpha-synuclein[J]. The Journal of Biological Chemistry, 2007, 282(8):5641-5652.
[43] ZHANG X, YUAN Y, JIANG L, et al. Endoplasmic reticulum stress induced by tunicamycin and thapsigargin protects against transient ischemic brain injury: Involvement of PARK2-dependent mitophagy[J]. Autophagy, 2014, 10(10):1801-1813.
2016-01-11
国家自然科学基金资助项目 (81171947,31271481)
张文亮(1991年生),男;研究方向:细胞自噬与肿瘤发生;通信作者:王海河;E-mail:wanghaih@mail.sysu.edu.cn
Q-331
A
0529-6579(2016)04-0108-10
Earle’s盐平衡液饥饿诱导DLD-1, HCT-116, A2780, CHO,Hep G2和 SMMC7721肿瘤细胞系自噬发生的时间优化
张文亮, 张明明,刘 斌,石宏顺,王海河
(中山大学中山医学院生物化学系∥中山大学干细胞与组织工程中心,广东 广州 510080)
肿瘤细胞在正常培养条件下自噬水平非常低,很难被监测到。与药物性自噬诱导剂相比,Earle’s盐平衡液(EBSS)应用简便,除产生饥饿诱导自噬外,无其他细胞毒性,有利于研究细胞自噬的分子机制和功能。本文应用蛋白免疫印记和细胞免疫荧光技术探讨了EBSS对DLD-1和HCT-116结直肠癌、A2780和CHO卵巢癌、HepG2和SMMC7721肝癌细胞系的自噬诱导效果和有效诱导时间。结果显示:EBSS短短几小时处理,能够非常有效地诱导上述6株肿瘤细胞发生自噬,但不同细胞系之间的有效诱导时间却明显不同。其中,DLD-1、HCT-116、CHO和HepG2细胞系的有效诱导时间仅需2 h;A2780细胞需要稍微延长到4 h;然而SMMC7721却要延长到8 h左右。本研究为其他研究者在应用EBSS诱导肿瘤细胞自噬时提供了实践依据和参考。
细胞自噬诱导;Earle’s盐平衡液;LC3; SQSTM1/p62;癌细胞
10.13471/j.cnki.acta.snus.2016.04.018
猜你喜欢
海河水利(2021年6期)2021-12-28
金桥(2021年9期)2021-11-02
金桥(2021年9期)2021-11-02
金桥(2021年9期)2021-11-02
建材发展导向(2021年23期)2021-03-08
中国生殖健康(2020年2期)2021-01-18
军事文摘(2020年22期)2021-01-04
历史地理研究(2020年2期)2020-08-10
学苑创造·A版(2020年6期)2020-07-07
当代水产(2019年1期)2019-05-16
