
水分胁迫后复水对糜子根冠功能的补偿效应
2016-06-03 12:06赵伟洁杨清华高小丽冯佰利
西北农林科技大学学报(自然科学版) 2016年1期
关键词:糜子
赵伟洁,李 翠,周 达,周 瑜 ,杨清华,高小丽,冯佰利
(1 西北农林科技大学 农学院,陕西 杨凌 712100;2 四川省宜宾市宜宾县农业局,四川 宜宾 644600)
水分胁迫后复水对糜子根冠功能的补偿效应
赵伟洁1,2,李翠1,周达1,周瑜1,杨清华1,高小丽1,冯佰利1
(1 西北农林科技大学 农学院,陕西 杨凌 712100;2 四川省宜宾市宜宾县农业局,四川 宜宾 644600)
[摘要]【目的】 研究水分胁迫后复水对糜子根、冠生长和光合作用的补偿规律及补偿条件的影响,为糜子抗旱节水栽培和补偿效应研究提供理论依据。【方法】 采用盆栽控水方式,以全生育期正常供水为对照,分别在糜子不同生育时期水分胁迫后复水,对复水后糜子根、冠功能的恢复以及复水前后新老叶片净光合速率等指标进行分析。【结果】 (1)水分胁迫后复水,根冠功能和光合作用的补偿效应与胁迫持续时间、胁迫程度及复水时期有关。(2)三叶一心期水分胁迫复水后,重度胁迫拔节期复水处理的根冠比、根系伤流量和叶面积的恢复最快,分别高于对照和中度胁迫拔节期复水处理;同时,拔节期复水处理各指标的补偿效应高于抽穗期复水的2个处理。拔节期水分胁迫复水后,中度胁迫抽穗期复水处理的根冠比、根系伤流量和叶面积高于对照和重度胁迫复水处理。(3)与对照相比,无论是三叶一心期还是拔节期开始水分胁迫,胁迫前生长出的叶片复水后其净光合速率低,胁迫程度越重净光合速率越小;而复水后长出的新叶,三叶一心期重度水分胁迫拔节期复水处理新叶净光合速率最高,复水20 d时比对照高出14.5%,中度水分胁迫拔节期复水后新叶净光合速率与对照无显著差异。【结论】 三叶一心期重度水分胁迫拔节期复水有利于新生器官生理活性的提高,对糜子根冠功能和净光合速率的补偿效应最佳。
[关键词]糜子;水分胁迫;复水;根冠功能;净光合速率;补偿效应
糜子(PanicummiliaceumL.)是我国重要的粮食作物和经济作物[1],年播种面积70~80万hm2,主要分布在陕西、山西、甘肃、宁夏、内蒙古等长城沿线风沙区,干旱是对糜子生产影响最大的限制因素[2]。因此,加强糜子抗旱生理机制的研究具有重要意义[3]。
不同生育时期作物对水分的需求不同,受到不同程度水分胁迫复水后其反应也不尽相同[4]。根系作为植物水分、养分的吸收器官,其数量、大小、干质量和生理状况等直接影响作物抗旱性的强弱[5]。Ilknur等[6]研究指出,作物生长过程中生殖生长对水分胁迫的敏感程度大于营养生长。高志红等[7]研究指出,水分胁迫抑制小麦根、冠功能的发挥,胁迫愈强影响程度越重,但是开花期复水后可以迅速提高干物质的积累速度。郝树荣等[8-9]研究认为,水稻旱后复水的补偿效果明显,特别是分蘖末期短历时重旱复水后有利于根质量、根冠比以及根系活力的增加,而且旱后复水新生器官活性的提高是水稻产生补偿效应的主要贡献者。Singh等[10]认为,水分胁迫条件下叶片净光合速率显著降低。薛慧云等[11]研究指出,净光合速率能在干旱胁迫初期和持续干旱的过程中反映叶片的水分状况,复水后净光合速率未能恢复到初始状态。而蒙祖庆等[12]研究指出,在复水后1~2周作物净光合速率恢复迅速,在第2周时净光合速率高于对照。目前有关水分胁迫及复水的研究主要集中在小麦[13]、水稻[8]、玉米[14]的生理生态响应、抗旱遗传资源等方面,而有关水分胁迫后复水对糜子根、冠生长影响的研究尚未见报道,尤其是对糜子复水前后新老叶片光合作用的补偿规律、产生补偿的条件、补偿形式更缺乏深层次的研究。因此,本研究通过盆栽试验,对糜子复水后根、冠功能的恢复和复水前后新老叶片净光合速率的影响进行探讨,旨在为糜子抗旱节水栽培和补偿效应研究提供理论依据。
1材料与方法
1.1试验设计
试验于2012-2013年在西北农林科技大学干旱棚进行,采用直径30 cm、高35 cm的聚乙烯塑料桶进行盆栽试验。试验土壤类型为垆土,养分含量为有机质2.4 g/kg,全氮0.03 g/kg,速效磷4.6 mg/kg,速效钾102 mg/kg,田间最大持水量26.20%。土壤经风干、打碎、过筛后与基肥充分混合,每盆装干土12 kg,基肥用量为每千克干土施P2O50.2 g、N素0.25 g,每盆另施尿素6.0 g,磷酸二氢钾4 g。供试糜子品种为榆糜2号,2012-07-05播种,09-24收获;2013-06-18播种,09-08收获;每盆播种30粒,三叶一心期定苗,每盆留苗12株。
试验共设置3个水分梯度:分别为充足供水(占田间最大持水量的75%±5%,对照)、中度水分亏缺(占田间最大持水量的60%±5%)、重度水分亏缺(占田间最大持水量的40%±5%);并结合不同生育期共设置了7个处理:全生育期充足供水(CK)、三叶一心期-拔节期中度水分胁迫后复水(SMB)、三叶一心期-拔节期重度水分胁迫后复水(SSB)、三叶一心期-抽穗期中度水分胁迫后复水(SMC)、三叶一心期-抽穗期重度水分胁迫后复水(SSC)、拔节期-抽穗期中度水分胁迫后复水(BMC)、拔节期-抽穗期重度水分胁迫后复水(BSC)。每个处理重复15盆,各盆随机摆放,每10 d随机调换1次位置排除边际效应。每天早晨用感量2 g 的TCS-A1-30型计重台秤称各盆质量,低于控水下限时补充所失水分。
1.2测定项目及方法
糜子根冠获取:各处理胁迫结束时以及复水后每隔7 d取样,先从茎基部剪下,获得完整的冠,然后将盆栽用桶放在尼龙网筛上,用水冲去泥土,获取完整根系,再根据需要从不同部位取样。
根、冠干质量的测定:在105 ℃下杀青15 min后,置于70 ℃恒温下烘干至恒质量,用1/10 000电子天平称质量。
根系伤流量测定:抽穗期后每隔7 d,采用邱全胜等[15]的改进重量法测定根系伤流强度,以单位时间内单株伤流量的大小计算。
叶面积测定:叶片取样时间为开花期和成熟期,量取每片绿色叶片的长度和最大宽度,按冯冬霞等[16]的叶面积拟合公式计算。
净光合速率(Pn)测定:用LI-6400型便携式光合仪测定,2012年净光合速率在抽穗至成熟期间每隔7 d测定1次,共测定6次;2013年在胁迫结束时及复水后7 d、14 d各测量1次,共测定3次。每次测量保证在相同时间段测定相同叶片的相同部位,测量时新生叶片为复水后长出的叶片,老叶片为胁迫结束时的最上部叶片。
1.3数据处理
采用 Excel 2003,DPS 统计软件进行相关试验数据的处理和分析。
2结果与分析
2.1复水对糜子根、冠功能的补偿效应
2.1.1复水对糜子根冠比的影响由表1可见,与CK比较,短历时水分胁迫复水后有利于糜子根系的生长,根冠比增长速度快;长历时水分胁迫抑制糜子根冠的生长,根冠比降低,且胁迫程度越重,根冠比越小。拔节期复水后7 d,重度水分胁迫处理的根冠比大于中度水分胁迫处理,到复水后14 d,中度水分胁迫处理的根冠比则大于重度水分胁迫处理。复水初期根对水分的反应更敏感,但随着时间的推移复水对冠的生长促进作用更大。与拔节期复水不同,抽穗期复水时,三叶一心期开始胁迫处理表现为中度水分胁迫处理根冠比大于重度水分胁迫处理;而拔节期开始胁迫处理复水后则是重度水分胁迫处理根冠比大于中度水分胁迫;三叶一心期中度、重度水分胁迫处理复水后根冠比较拔节期中度、重度水分胁迫处理复水后低。

表 1 复水后糜子根、冠干质量及根冠比的变化(2013年)

续表 1 Continued table 1
注:同列不同大写字母表示同一复水期不同处理间差异极显著(P<0.01) 。下表同。
Note:Capital letters in each column show significant difference between treatments atP<0.01 level.The same below.
2.1.2复水对糜子根系活力的影响从图1可以看出,拔节期复水后糜子根系活力有明显提高,在抽穗后14 d根系伤流量分别比对照高15.6%和 5.1%,表现出超补偿效应,具有较高的根系活力,有利于植株吸收水分和无机矿物质,为糜子的生长提供了充分的后效补偿。而抽穗期复水SMC、SSC 2个处理的根系伤流量则低于对照,对作物的补偿效果不明显。
如图2所示,拔节期水分胁迫使糜子根系活力降低,胁迫程度越重活力越低。抽穗期复水时,中度水分胁迫复水后14 d根系伤流量比对照高9.26%,而重度胁迫复水后根系伤流量比对照低16.29%。这可能是由于拔节期后,糜子的生长重心由地下转向地上,根系抗旱能力低,重度水分胁迫对根系的抑制作用大,因此复水后根系活力的补偿效应较中度水分胁迫差。从图1和图2可以看出,水分胁迫复水后糜子根系活力不仅取决于胁迫历时,还取决于胁迫的时期,三叶一心期重度水分胁迫拔节期复水处理的补偿效果最佳。

图 1 三叶一心期胁迫后复水对糜子根系
2.2复水对糜子叶片及叶面积的补偿效应
从图3可以看出,从开花期到成熟期,糜子绿叶数在不断减少,与对照相比,三叶一心期重度水分胁迫拔节期复水处理和拔节期中度水分胁迫抽穗期复水处理在整个生育期保持更多的绿叶数,提高了“源”的强度,有利于光合产物的积累,为糜子产量的提高提供了良好的冠层条件。拔节期复水,重度水分胁迫复水处理对保持后期叶片数的后效性大于中度水分胁迫处理,而抽穗期复水时则是中度水分胁迫后复水对保持后期叶片数的后效性大于重度水分胁迫处理,同时拔节期胁迫抽穗期复水处理对保持后期叶片数的后效性大于三叶一心期胁迫抽穗期复水处理。
由图4可见,与对照相比,三叶一心期重度水分胁迫拔节期复水处理在开花期和成熟期均保持了更大的叶面积,分别比对照增加了6.22%和15.69%,而三叶一心期中度水分胁迫拔节期复水处理补偿效果不明显,表明三叶一心期重度水分胁迫拔节期复水处理延缓了后期叶片的衰老、延长了叶片的光合时间,有利于穗生长和灌浆。抽穗期复水时,拔节期中度水分胁迫处理的复水效果最佳,其叶面积在开花期、成熟期比对照分别增加了2.46%和13.92%,而拔节期重度水分胁迫和三叶一心期中度、重度水分胁迫处理的后效补偿效果不佳,叶面积均低于对照。
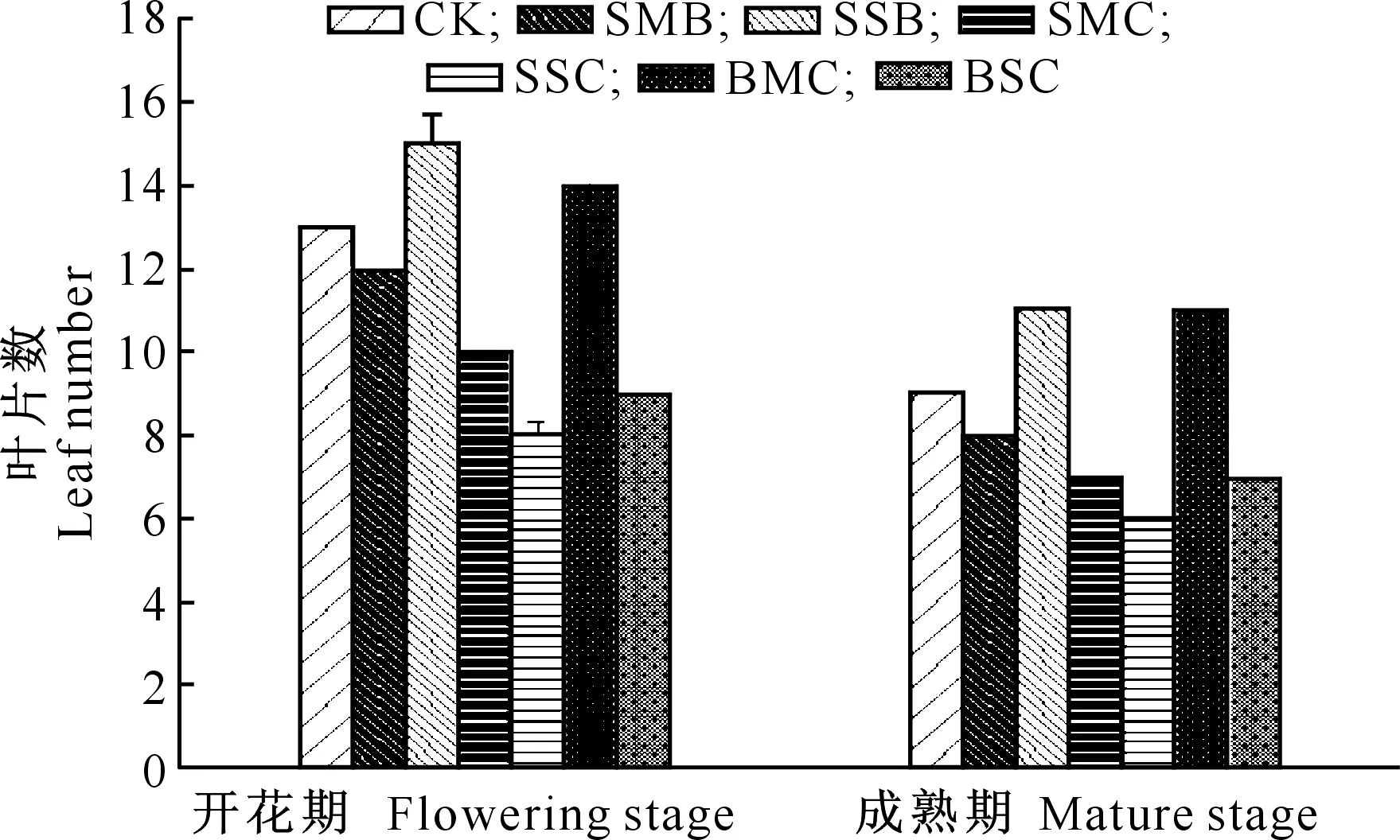
图 3 不同水分胁迫处理下糜子叶片数的变化(2013年)

图 4 不同水分胁迫处理下糜子叶面积的变化(2013年)
2.3复水对糜子光合补偿发生规律的影响
由表2可见,三叶一心期水分胁迫拔节期复水时,重度水分胁迫处理叶片的净光合速率比对照和中度水分胁迫复水处理恢复得更快、恢复程度也更高,到开花期时其净光合速率分别比对照、中度水分胁迫高10.3%和9.4%;三叶一心期水分胁迫抽穗期复水时,则是中度水分胁迫开花期叶片的净光合速率恢复程度高于重度水分胁迫处理,但其恢复程度均低于对照。拔节期水分胁迫抽穗期复水时,与对照相比,开花期时中度水分胁迫复水后叶片净光合速率恢复较快,其净光合速率比对照高5.1%;而重度水分胁迫复水后叶片的净光合速率则低于对照。

表 2 胁迫后复水对糜子叶片净光合速率的影响(2012年)
如表3所示,三叶一心期重度水分胁迫拔节期复水和拔节期中度水分胁迫抽穗期复水处理的糜子叶片净光合速率恢复迅速,复水后14 d分别比对照增加11.2%和15.08%,表现出超补偿效应,与2012年结论一致。

表 3 胁迫后复水对糜子叶片净光合速率的影响(2013年)
2.4复水对糜子新老叶片净光合速率补偿效应的影响
由表4可见,与CK相比,无论是三叶一心期还是拔节期开始水分胁迫,胁迫前生长出的叶片复水后其净光合速率低,胁迫程度越重净光合速率越小。说明受到胁迫抑制生长的叶片复水后不能恢复到原有的光合生产能力。

表 4 胁迫后复水对糜子老叶片净光合速率的影响(2013年)
从图5可以看出,三叶一心期重度水分胁迫拔节期复水处理糜子叶片的净光合速率均大于对照,复水后20 d时净光合速率达到最大,比对照高出14.5%,产生了超补偿效应。同时,三叶一心期中度水分胁迫拔节期复水后糜子叶片的净光合速率则与对照差异不显著。
由图6可见,拔节期中度水分胁迫抽穗期复水后10 d时,新生叶片的净光合速率最大,比对照高10.8%,表现出超补偿效应,但与三叶一心期胁迫拔节期复水不同的是随着复水时间的延长,其补偿作用不断下降。说明抽穗期复水的补偿能力不如拔节期,拔节期胁迫抽穗期复水时,只在复水初期补偿效果表现明显,后期补偿效果有限。同时,拔节期中度水分胁迫抽穗期复水后光合补偿能力大于重度水分胁迫。由此可见,糜子叶片光合补偿效应是受多因素影响的,它不仅与水分胁迫程度有关,还与胁迫的生育阶段以及胁迫时间有关。

图 5 三叶一心期胁迫拔节期复水后对糜子
3讨论
作物对水分胁迫的响应有一个从“伤害”到“适应”的过程,一定程度的水分胁迫后复水往往会产生生长、生理和产量上的补偿[17]。根冠比是作物根冠所具备的吸收、合成、分配等基本功能的结构体现[18]。Magnani等[19]指出,根冠结构与功能处于均衡状态时,其资源利用效率最高,但环境改变时平衡即被打破。前人研究表明,旱长根水长冠,水分胁迫使根冠比增大[20],而本研究结果显示,水分胁迫抑制糜子根冠的生长,且对根的抑制作用大于冠,胁迫使根冠比降低,且胁迫程度越重,根冠比越小。从生育时期看,拔节期复水根较冠对水分的反应更敏感,但随着时间的推移复水对冠的生长促进作用更大,使复水后根冠比处于减小趋势。由此可见,根冠比的大小,不仅与水分胁迫程度,还与胁迫时期和胁迫历时都有着紧密的联系。
根系是糜子吸收水分的主要器官,并能迅速产生化学信号向上传递以促使气孔关闭,减少水分散失[21];根系还可以通过自身形态和生理生化特征的调整来适应变化后的水分环境。本研究发现,不同程度水分胁迫复水后对根系吸水恢复能力的影响依不同胁迫时期而有所变化,起始于三叶一心期的水分胁迫,重度水分胁迫拔节期复水的吸水效果反而比中度水分胁迫拔节期复水好,这可能是因为三叶一心期重度水分胁迫后复水产生的根系活性大于中度水分胁迫,但拔节期或三叶一心期长历时水分胁迫对根系吸水的不利影响则随胁迫程度加重而加大。同时,不同程度水分胁迫下根系吸收水分的方式也有差异,重度水分胁迫复水后根系吸水恢复主要取决于新生根的增加,对于失活根系的激发占次要地位;而中度水分胁迫复水后根系吸水功能的恢复主要依赖于对原有失活根系活性的激发,新根的生长占次要地位。这与刘晓英等[22]对冬小麦的研究结果一致,但有关新生根和原有根系复水后的功能还有待进一步研究。
干旱胁迫通过气孔限制和非气孔限制来抑制植物的光合作用,使得净光合速率、光合产物的量下降[23],但复水后,在一定范围内能解除干旱胁迫对光合作用的抑制,产生相应的后效补偿[24]。本研究发现,水分胁迫复水后糜子净光合速率的增加,是由复水后新生器官生理活性的提高而产生的补偿效应,并不是受到胁迫的老叶光合机能的恢复,直接遭受胁迫的老叶净光合速率无论在胁迫期间还是复水后均呈降低趋势。
4结论
复水对水分胁迫条件下糜子根、冠生长的补偿效应明显,尤以三叶一心期重度胁迫拔节期复水的补偿效应最佳。复水后糜子根、冠生物量的积累快,新生器官生理活性高,有利于作物的生长,能够准确地反映作物旱后复水对水分的需求状况。因此,糜子生产过程中,三叶一心期水分胁迫后复水可作为节水高产栽培技术措施的理论参考。
[参考文献]
[1]张雄,王立祥.小杂粮在黄土高原旱作农业中的地位和作用 [J].西北农业学报,2008,17(5):333-336.
Zhang X,Wang L X.The status and role of minor food crops in dry farming of the Loess Plateau [J].Acta Agriculturae Boreali-Occidentalis Sinica,2008,17(5):333-336.(in Chinese)
[2]Chai Y,Feng B L.Advances in broomcorn billet research:Proceedings of the 1st international symosium on broomcorn millet [M].Yangling,Shaanxi:Northwest A&F University Press,2012.
[3]林汝法,柴岩.中国小杂粮 [M].北京:中国农业科技出版社,2002:68-77.
Lin R F,Chai Y.Minor grain crops in China [M].Beijing:China Agricultural Science and Technology Press,2002:68-77.(in Chinese)
[4]赵长星,程曦,王月福,等.不同生育时期干旱胁迫对花生生长发育和复水后补偿效应的影响 [J].中国油料作物学报,2012,34(6):627-632.
Zhao C X,Cheng X,Wang Y F,et al.Effects of drought stress on peanut growth during different growth stages and compensatory effect after water recovery [J].Chinese Journal of Oil Crop Sciences,2012,34(6):627-632.(in Chinese)
[5]Yousfi N,Slama I,Ghnaya T,et al.Effects of water deficit str-ess on growth,water relations and osmolyte accumulation inMedicagotruncatulaandM.laciniatapopulations [J].Comptes Rendus Biologies,2010,333:205-213.
[6]Ilknur K,Elman B,Alain C.Growth and yield responses of cv.Merlot(VitisviniferaL.) to early water stress [J].African Journal of Agricultural Research,2011,29(6):6281-6288.
[7]高志红,陈晓远,罗远培.不同土壤水分条件下冬小麦根、冠平衡与生长稳定性研究 [J].中国农业科学,2007,40(3):540-548.
Gao Z H,Chen X Y,Luo Y P.Winter wheat root and shoot:Equilibrium and growth stability under different soil and water conditions [J].Scientia Agricultura Sinica,2007,40(3):540-548.(in Chinese)
[8]郝树荣,郭相平,张展羽.水分胁迫及复水对水稻冠层结构的补偿效应 [J].农业机械学报,2010,41(3):52-55,61.
Hao S R,Guo X P,Zhang Z Y.After effects of rewatering after water stress on the rice growth [J].Transactions of the Chinese Society for Agricultural Machinery,2010,41(3):52-55,61.(in Chinese)
[9]郝树荣,郭相平,张展羽.水稻根冠功能对水分胁迫及复水的补偿响应 [J].农业机械学报,2010,41(5):52-55.
Hao S R,Guo X P,Zhang Z Y.Compensation effects of water stress and re-watering on the function of root shoot [J].Transactions of the Chinese Society for Agricultural Machinery,2010,41(5):52-55.(in Chinese)
[10]Singh S K,Reddy K R.Regulation of photosynthesis,fluorescence,stomatal conductance and water-use efficiency of cowpea under drought [J].Journal of Photochemistry and Photobiology B:Biology,2011,105(1):40-50.
[11]薛慧云,张永江,刘连涛,等.干旱胁迫与复水对棉花叶片光谱、光合和荧光参数的影响 [J].中国农业科学,2013,46(11):2386-2393.
Xue H Y,Zhang Y J,Liu L T,et al.Responses of spectral reflectance,photosynthesis and chlorophyll fluorescence in cotton during drought stress and re-watering [J].Scientia Agricutural Sinica,2013,46(11):2386-2393.(in Chinese)
[12]蒙祖庆,宋丰萍,刘振兴,等.干旱及复水对油菜苗期光合及叶绿素荧光特性的影响 [J].中国油料作物学报,2012,34(1):40-47.
Meng Z Q,Song F P,Liu Z X,et al.Effects of drought and rewatering at seedling stage on photosynthesis and chlorophyll fluorescence characteristics in rapeseed [J].Chinese Journal of Oil Crop Sciences,2012,34(1):40-47.(in Chinese)
[13]马富举,李丹丹,蔡剑,等.干旱胁迫对小麦幼苗根系生长和叶片光合作用的影响 [J].应用生态学报,2012,23(3):724-730.
Ma F J,Li D D,Cai J,et al.Responses of wheat seedlings root growth and leaf photosynthesis to drought stress [J].Chinese Journal of Applied Ecology,2012,23(3):724-730.(in Chinese)
[14]刘洪展,郑伟,郑风荣,等.复水对海水浇灌的玉米幼苗根系补偿效应的影响 [J].农业工程学报,2012,28(3):101-106.
Liu H Z,Zheng W,Zheng F R,et al.Influence of rewatering on compensatory effect of maize seedling roots with diluted seawater irrigation [J].Chinese Society of Agricutural Engineering,2012,28(3):101-106.(in Chinese)
[15]邱全胜,李琳,梁厚果,等.水分胁迫对小麦根细胞质膜氧化还原系统的影响 [J].植物生理学报,1994,20(2):145-151.
Qiu Q S,Li L,Liang H G,et al.Effect of water stress on the redox system of the plasma membrane of wheat roots [J].Acta Phytophysiol Sin,1994,20(2):145-151.(in Chinese)
[16]冯冬霞,施生锦.叶面积测定方法的研究效果初报 [J].中国农学通报,2005,21(6):150-155.
Feng D X,Shi S J.Research on night measurement methods of leaf area [J].China Agricultural Science Bulletin,2005,21(6):150-155.(in Chinese)
[17]丁红,张智猛,戴良香,等.干旱胁迫对花生根系生长发育和生理特性的影响 [J].应用生态学报,2013,24(6):1586-1592.
Ding H,Zhang Z M,Dai L X,et al.Effects of drought stress on the root growth and development and physiological characteristics of peanut [J].Chinese Journal of Applied Ecology,2013,24(6):1586-1592.(in Chinese)
[18]冯烨,郭峰,李宝龙,等.单粒精播对花生根系生长、根冠比和产量的影响 [J].作物学报,2013,39(12):2228-2237.
Feng Y,Guo F,Li B L,et al.Effects of single-seed sowing on root growth,root-shoot ratio,and yield in peanut [J].Acta Agronomica Sinica,2013,39(12):2228-2237.(in Chinese)
[19]Magnani F,Mencuccini M,Grace J.Age-related decline in sta-nd productivity:The role of structural acclimation under hydraulic constraints [J].Plant,Cell and Environment,2000,23:251-263.
[20]刘水,李伏生,韦翔华,等.分根区交替灌溉对玉米水分利用和土壤微生物量碳的影响 [J].农业工程学报,2012,28(8):71-76.
Liu S,Li F S,Wei X H,et al.Effects of alternate partial root-zone irrigation on maize water use and soil microbial biomass carbon [J].Chinese Society of Agricultural Engineering,2012,28(8):71-76.(in Chinese)
[21]Jia W S,Zhang J H.Stomatal movements and long distance signaling in plants [J].Plant Signaling and Behavior,2008,3:772-777.
[22]刘晓英,罗远培.水分胁迫后复水冬小麦根系吸收的恢复 [J].中国生态农业学报,2002,10(4):16-23.
Liu X Y,Luo Y P.Recovery of root water uptake of winter wheat after water stress [J].Chinese Journal of Eco-Agriculture,2002,10(4):16-23.(in Chinese)
[23]Lawlor D W,Cornic G.Photosynthetic carbon assimilation and associated metabolism in relation to water deficits in higher plants [J].Plant,Cell and Environment,2002,25:275-294.
[24]杨文权,顾沐宇,寇建村,等.干旱及复水对小冠花光合及叶绿素荧光参数的影响 [J].草地学报,2013,21(6):1130-1135.
Yang W Q,Gu M Y,Kou J C,et al.Effects of drought and rewatering on the photosynthesis and chlorophyll fluorescence ofCoronillavaria[J].Acta Agrestia Sinica,2013,21(6):1130-1135.(in Chinese)
Compensation effects of rewatering on root and shoot functions of broomcorn millet after water stress
ZHAO Wei-jie1,2,LI Cui1,ZHOU Da1,ZHOU Yu1,YANG Qing-hua1,GAO Xiao-li1,FENG Bai-li1
(1CollegeofAgronomy,NorthwestA&FUniversity,Yangling,Shaanxi712100,China;2BureauofAgricultureinYibinCountyYibinCitySichuan,Yibin,Sichuan644600,China)
Abstract:【Objective】 Effects of rewatering on compensation mechanism and condition of root and shoot growth as well as shoot photosynthesis rate (Pn) of broomcorn millet were studied to provide basis for water saving cultivation of broomcorn millet.【Method】 Taking adequate water supply as CK,pot experiment was conducted to rewater after water stress at different growth periods.The root and shoot characteristics and Pn of new and old leaves before and after rewatering were measured and analyzed.【Result】 (1)Compensation effects of root and shoot functions as well as Pn correlated with the duration and intensity of water stress and rewatering time.(2)When water control was conducted from 3-leaf stage and rewatering was implemented from jointing stage,root to shoot ratio,root extrudes volume and leaf area of broomcorn millet under severe water stress increased faster than that of the CK and the treatment under moderate water stress.Besides,compensation effects of tested items were stronger compared with the two treatments when rewatering was conducted from heading stage.When water control was implemented from jointing stage and rewatering was conducted from heading stage,root to shoot ratio,root extrudes volume and leaf area of broomcorn millet under moderate water stress were larger than that of the CK and the treatment under severe water stress.(3)When water control was conducted from 3-leaf stage or from jointing stage,Pn of original leaves before water control was lower than that of the CK,and fiercer stress led to lower Pn.While for the new leaves after rewatering,Pn after rewatering from jointing stage under severe water stress from 3-leaf stage was the highest,14.5% higher than that of the CK 20 days after rewatering.When water stress was moderate,no significant difference was found between Pn of new leaves and that of the CK.【Conclusion】 Rewatering after early stage severe water stress was beneficial for the enhancement of physiological activities of the new organs,and the compensation effects for root and shoot growth as well as shoot photosynthesis were optimum.
Key words:broomcorn millet;water stress;rewatering;root and shoot functions;Pn;compensation
[文章编号]1671-9387(2016)01-0045-08
[中图分类号]S516.01
[文献标志码]A
[作者简介]赵伟洁(1989-),男,四川宜宾人,硕士,主要从事作物栽培生理研究。E-mail: jieweizhao1989@163.com[通信作者]冯佰利(1966-),男,陕西耀县人,教授,博士生导师,主要从事作物高产生态生理技术及小杂粮栽培、育种研究。
[基金项目]国家自然科学基金项目(31371529);国家谷子糜子产业技术体系项目(CARS-07-12.5-A9)
[收稿日期]2014-04-18
DOI:网络出版时间:2015-12-0214:2510.13207/j.cnki.jnwafu.2016.01.008
网络出版地址:http://www.cnki.net/kcms/detail/61.1390.S.20151202.1425.016.html
E-mail:7012766@163.com
猜你喜欢
中国粮油学报(2022年6期)2022-07-23
今日农业(2022年8期)2022-07-05
现代农村科技(2021年12期)2021-12-06
现代农村科技(2021年12期)2021-12-06
北方农业学报(2021年3期)2021-08-26
种子(2020年7期)2020-10-09
河北农业科学(2020年6期)2020-04-07
现代农村科技(2019年12期)2019-01-06
科学种养(2018年8期)2018-11-22
中国种业(2017年9期)2017-09-21
