
乡土珍贵树种钩栗幼苗种源间光合生理特性研究
2016-05-26 02:14王佩兰康华荣李柏海李志辉
中南林业科技大学学报 2016年3期
王佩兰 ,康华荣 ,李柏海 ,李志辉 ,张 斌 ,吴 毅
(1.中南林业科技大学 林学院,湖南 长沙 410004;2. 汨罗市林业局,湖南 汨罗 414400;3. 湖南省森林植物园,湖南 长沙 410116)
钩栗Castanopsis tibetana是壳斗科,栲属的常绿乔木。生长在海拔200~1 600 m的阔叶林中,主要分布在两湖、两广、安徽、浙江、江西和福建等地。果可实,味香甜,富含淀粉。树形优美,是一种优良的园林绿化树种[1]。目前,国内外对钩栗的研究较少,主要集中在钩栗种群生命过程研究、人工育苗技术研究和种子形态及生理特征研究等方面[2-5]。现在钩栗的野生种群数量和面积日趋缩小,人为因素干扰是一方面原因,该物种生物学特性则是另一重要原因。例如,光是影响植物生存和生长最重要的环境因子之一,不同的植物具有不同的光合特性,体现为应对不同的光环境时会有不同的适应策略[6-7]。近年来,许多研究者通过研究植物的光和生理生态特性来分析珍稀树种日趋减少的原因[8-10]。笔者从2013年起,在开展对福建建瓯、湖北建瓯、湖南浏阳、桑植和永顺5个种源进行优良种源初步选择时,就开始综合考虑与幼苗的生长量和光合特性等因素[11]。本研究在该研究的基础上,对1年生钩栗幼苗进行光合和生长的动态监测,掌握钩栗幼苗光合生理特性,为其良种选育与保护提供了重要的理论依据。
1 材料与方法
1.1 试验材料
2012年11月,分别于福建建瓯、湖北恩施、湖南浏阳、桑植、永顺采集钩栗种子,用青苔湿藏,任其自由萌发,无任何层积处理。2013年2月,取出所有萌芽种子进行容器育苗。2013年7月,选取生长较为一致的健壮苗木,测定相应指标。钩栗不同种源地及气象因子见表1。

表1 不同种源钩栗地理位置及气候概况Table 1 Geographic and climate data of different provenances of Castanopsis tibetana
1.2 实验方法
1.2.1 光合参数测定
于2013年10月(秋季),2014年1月(春季),2014年4月(夏季),2014年7月(夏季)中下旬,选择晴朗的天气,从每个种源中选取3棵长势基本一致的钩栗,采用Li-6400光合仪测定成熟健康叶片的光合参数,从8:00~16:00每2 h测定一次,每个叶片重复记录5组数据,取平均值。测定时采用开放气路,以空气中CO2浓度为气源,测定的参数为:净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(Tr)。
1.2.2 光响应测定
采用Li-6400光合仪,在9:00~11:00对植物进行光响应曲线的测定。设置光强梯度从小到大 依 次 为:0、25、50、75、100、150、200、300、600、900、1 200、1 500、1 800 和 2 000 μmol·m-2s-1,测定各种源钩栗光合速率,流速控制在 500 μmol·m-2s-1。
1.2.3 幼苗叶片叶绿素含量测定
使用紫外可见分光光度计测OD663、OD645吸光值,计算叶绿素a、b以及叶绿素总量。
1.3 数据处理
采用SPSS17.0软件对光合数据进行方差分析,采用Excel和Origin7.5软件进行整理分析和绘图。用Photosyn Assistant1.1.2软件拟合光响应曲线,得到光补偿点(LCP)、光饱和点(LSP)、最大净光合速率(Pnmax)、表观量子效率(AQY)和暗呼吸速率(Rd)。拟合方程为Pn={AQY·PRA+Pnmax-[( AQY·PRA)2-4·AQY·PRA·K·Pnmax]0.5}/2K-Rd[12]。
2 结果与分析
2.1 不同季节环境因子变化
光合有效辐射(PRA)四季的变化规律基本一致,呈现出先升高后降低的走势,全年夏季的PRA 最大达到 2 030 μmol·m-2s-1,然后依次是秋季(1 860Pnmax)、春季(1 350Pnmax)和冬季(1 020Pnmax)。大气温度的季节性变化同PRA变化规律,夏季中午最高温度达到43°,4个季节中大气相对湿度的变化与PRA变化相反,12:00时降到最低点,春季大气相对湿度早上最高可达93%。
2.2 不同种源钩栗光合作用日进程
如图1所示,5个种源钩栗的净光合速率(Pn)日变化在春季、夏季和秋季3个季节中呈现为“双峰”曲线,第1个峰值出现在10:00,在12:00出现一个最低值,即光合“午休”现象,第2个峰值出现在下午16:00,春季和夏季Pn的第2个峰值比第1个峰值大,秋季Pn的第2个峰值比第1个峰值大。钩栗Pn日变化在冬季则呈现为“单峰”曲线,随着光强的增加,叶片上午的Pn随之升高,14:00达到最高峰,然后随着光照强度减弱,下午叶片的Pn下降。4个季节钩栗的日均Pn值均表现为夏季>秋季>春季>冬季,5个种源钩栗中,恩施种源四季的日均Pn值均为最大,与其他4个种源差异显著,其他4个种源的日均Pn值大小排序在四个季节中有差异,综合四季的日均Pn值得出5个钩栗种源的光合能力强弱顺序为:恩施>桑植>永顺>浏阳>建瓯(见表2)。

图1 不同种源钩栗净光合速率(Pn)日进程季节性变化Fig. 1 Seasonal change in daily process of net photosynthetic rate of Castanopsis tibetana in different provenances
如图2所示,5个种源钩栗的气孔导度(Gs)日变化曲线在春季、夏季和秋季均表现为“N”型,Gs值在10:00时达到峰值,之后逐渐下降, 14:00达到最小值后又开始缓慢上升。钩栗Gs日变化规律在冬季为“单峰”曲线,与Pn的日变化规律一致,峰值出现在12:00。4个季节中钩栗的日均Gs均表现为:夏季>秋季>春季>冬季。
如图3所示,5个种源钩栗在春季、夏季、秋季和冬季胞间CO2浓度(Ci)的日变化规律与净光合速率(Pn)的日变化规律相反,钩栗Ci日变化曲线在春季、夏季和秋季这3个季节中呈现为“W”字形,Ci值先随着光强的增大而减小,10:00时达到第1个低值,之后Ci值随着光强的增大迅速上升,12:00时达到最大值,随后Ci下降,14:00达到第2个低值后又缓慢上升。钩栗Ci日变化曲线在冬季则呈现为“V”字形,Ci值随着光强的增大而减小,在12:00达到最小值后,随着光强增大而增大。4个季节钩栗的日均Ci值均表现为:夏季>冬季>秋季>春季。
如图4所示,5个种源钩栗的蒸腾速率Tr日变化规律与净光合速率(Pn)日变化规律一致。在春季、夏季和秋季呈现为“双峰”曲线,冬季则呈现为“单峰”曲线。4个季节钩栗的日均Tr值均表现为夏季>秋季>春季>冬季。
2.3 不同种源钩栗光合参数变化
图5为不同种源钩栗四季的光响应曲线图。如图所示,5个种源钩栗在春、夏、秋、冬的Pn-PAR曲线变化趋势基本一致。PAR=0 ~ 200 μmol·m-2s-1,净光合速率呈直线上升,在PAR=200~600 μmol·m-2s-1时,净光合速率缓慢增长,光合速率达到饱和,净光合速率在 PAR=600 ~ 1 500 μmol·m-2s-1区间内,保持较大的光合速率不变,最大净光合速率出现在 1 000 ~ 1 500 μmol·m-2s-1范围内,当 PAR 大于1 500 μmol·m-2s-1后,Pn值略有下降,表现出强光抑制现象。
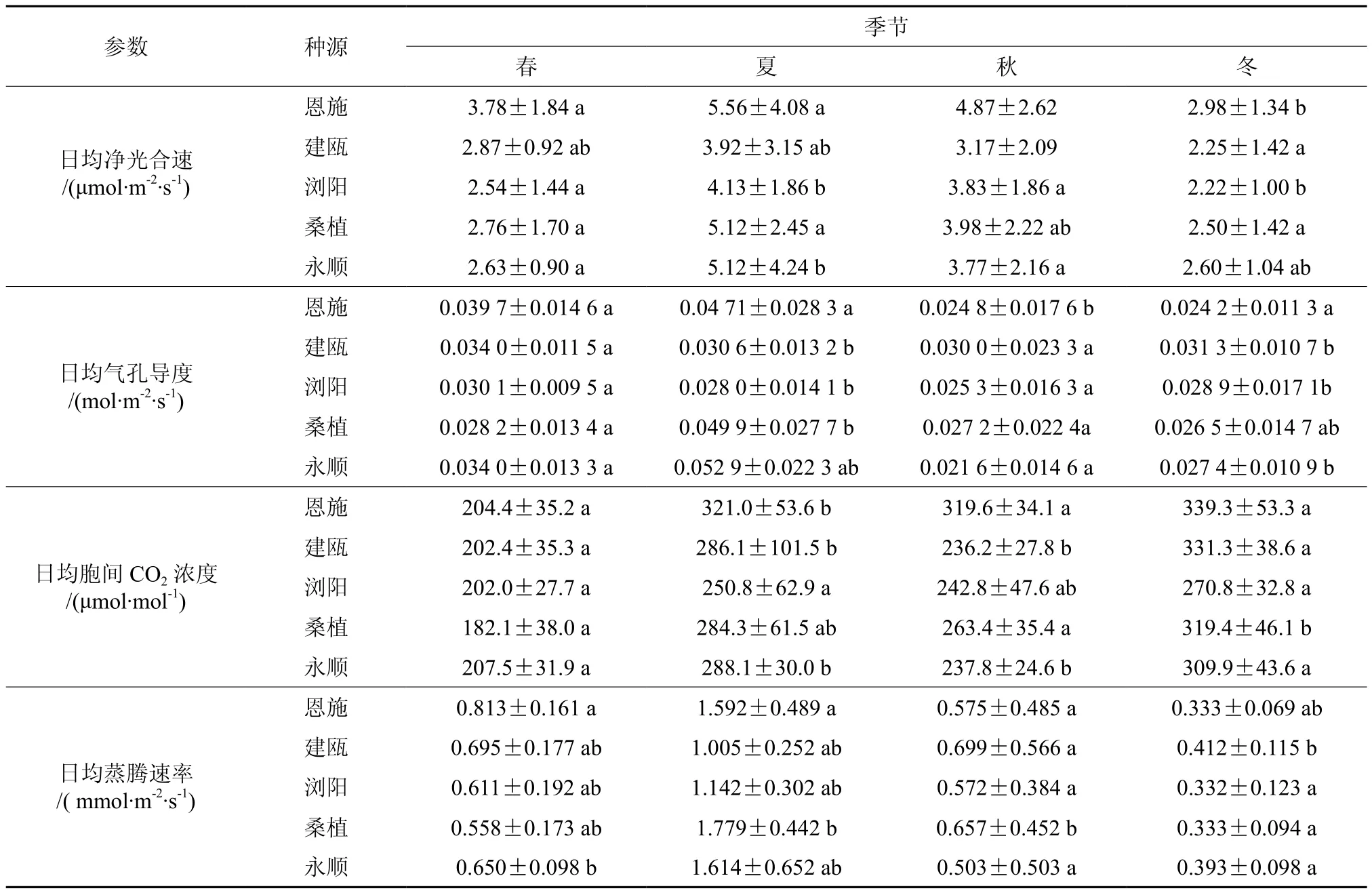
表2 不同种源钩栗日均净光合速率、日均气孔导度、日均胞间 CO2浓度、日均蒸腾速率的季节性变化†Table 2 Seasonal change in daily mean values of net photosynthetic rate, stomatal conductance, intercellular CO2 concentration and transpiration rate of Castanopsis tibetana leaves in different provenances
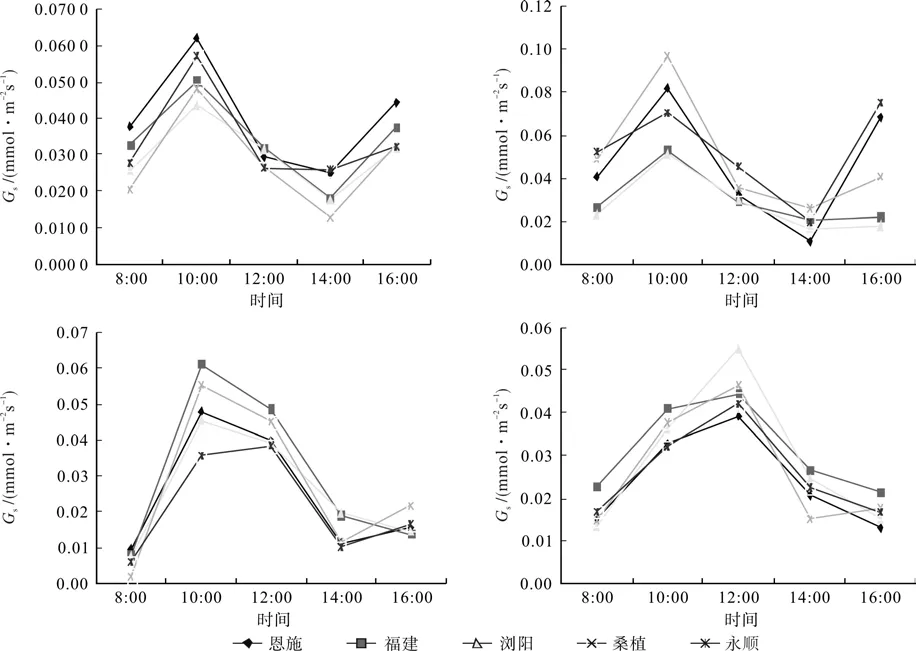
图2 不同种源钩栗气孔导度(Gs)日进程季节性变化Fig. 2 Seasonal change in daily process of stomatal conductance of Castanopsis tibetana in different provenances

图3 不同种源钩栗胞间CO2浓度(Ci)日进程季节性变化Fig.3 Seasonal change in daily process of intercellular CO2 concentration of Castanopsis tibetana in different provenances

图4 不同种源钩栗胞间蒸腾速率(Tr)日进程季节性变化Fig. 4 Seasonal change in daily process of transpiration rate of Castanopsis tibetana in different provenances
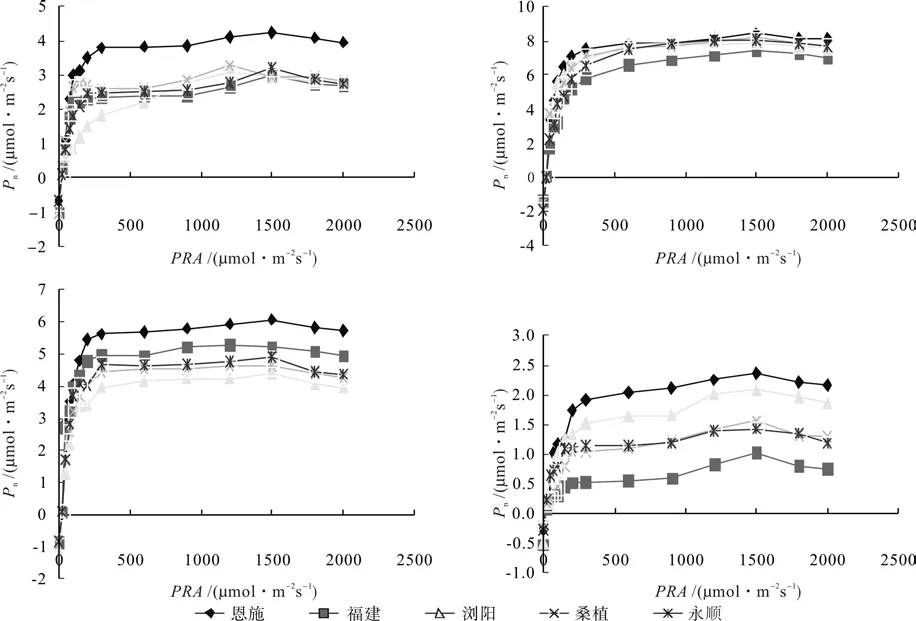
图5 不同种源钩栗叶片四季的光合作用-光响应曲线Fig.5 Seasonal photosynthesis-light response curves in leaves of different provenances of Castanopsis tibetana
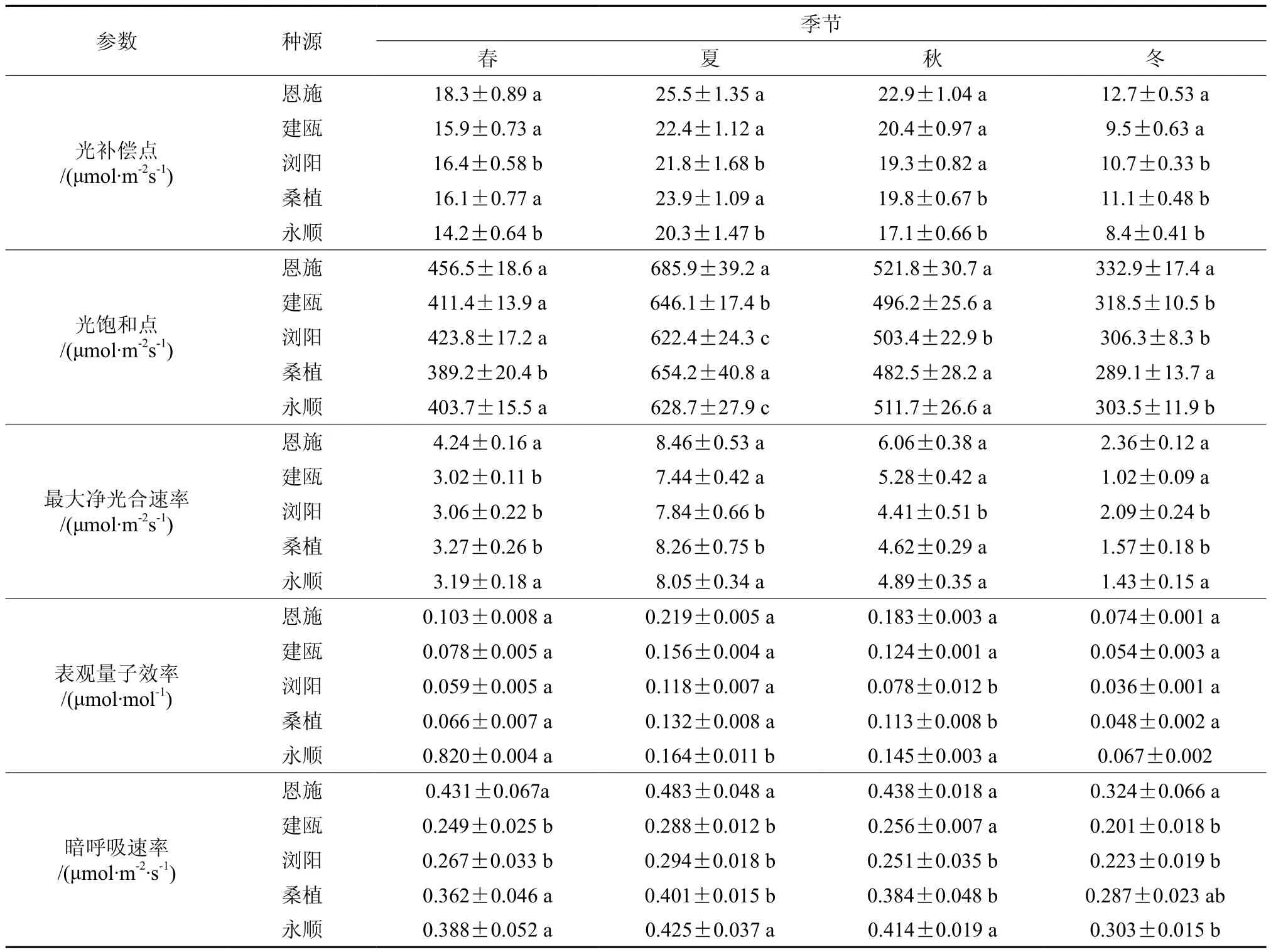
表3 不同种源钩栗光合参数季节性变化Table 3 Seasonal change in photosynthesis paramenter of Castanopsis tibetana of different provenances
由表3可知,5个种源钩栗的光补偿点(LCP)、光饱和点(LSP)、表观量子效率(AQY)、暗呼吸速率和最大净光合速率(Pnmax)均表现为夏季>秋季>春季>冬季,夏季的3个参数显著高于秋季、冬季和春季,4个季节的参数均差异显著。恩施种源的LCP值在4个季节中均表现为最大,永顺种源的LCP值最小,其他3个种源之间差异不显著,综合考虑5个种源的LCP值大小排序为:恩施>桑植>浏阳=建瓯>永顺。恩施种源的LSP值在春季和夏季明显大于其他4个种源,其他4个种源无显著差异,在秋季和冬季,恩施种源的LSP值仍然保持最大,但除了和LSP值最小的桑植种源差异显著外,与其他3个种源无显著差异,综合考虑5个种源的LSP值大小排序为:恩施>建瓯>浏阳>永顺>桑植。钩栗的AQY值在夏秋季节显著高于春冬季节,5个种源Rd值排序为:恩施>永顺>桑植>浏阳>建瓯。钩栗的Rd值为冬季显著低于其他季节,夏季为最大,春秋两季无显著差异,5个种源Rd值大小排序同AQY值。5个种源间最大净光合速率存在一定差异, 但未达到显著水平,恩施种源的Pnmax值优势明显,综合虑5个种源的Pnmax值大小排序为:恩施>浏阳=桑植=永顺>建瓯。
2.4 不同种源钩栗叶片叶绿素含量变化
在植物光合作用中叶绿素起着吸收光能的作用,叶绿素含量的高低直接影响植物光合能力的强弱[13]。由表4可知,5个种源钩栗的叶绿素a、b和a+b含量均以冬季最低,叶绿素a和a+b,冬季与其他季节均差异显著,但是叶绿素b,各个季节差异不显著。在4个季节中,恩施、桑植和永顺3个种源的叶绿素a、b和a+b含量明显高于建瓯和浏阳种源,但是彼此间差异不大。
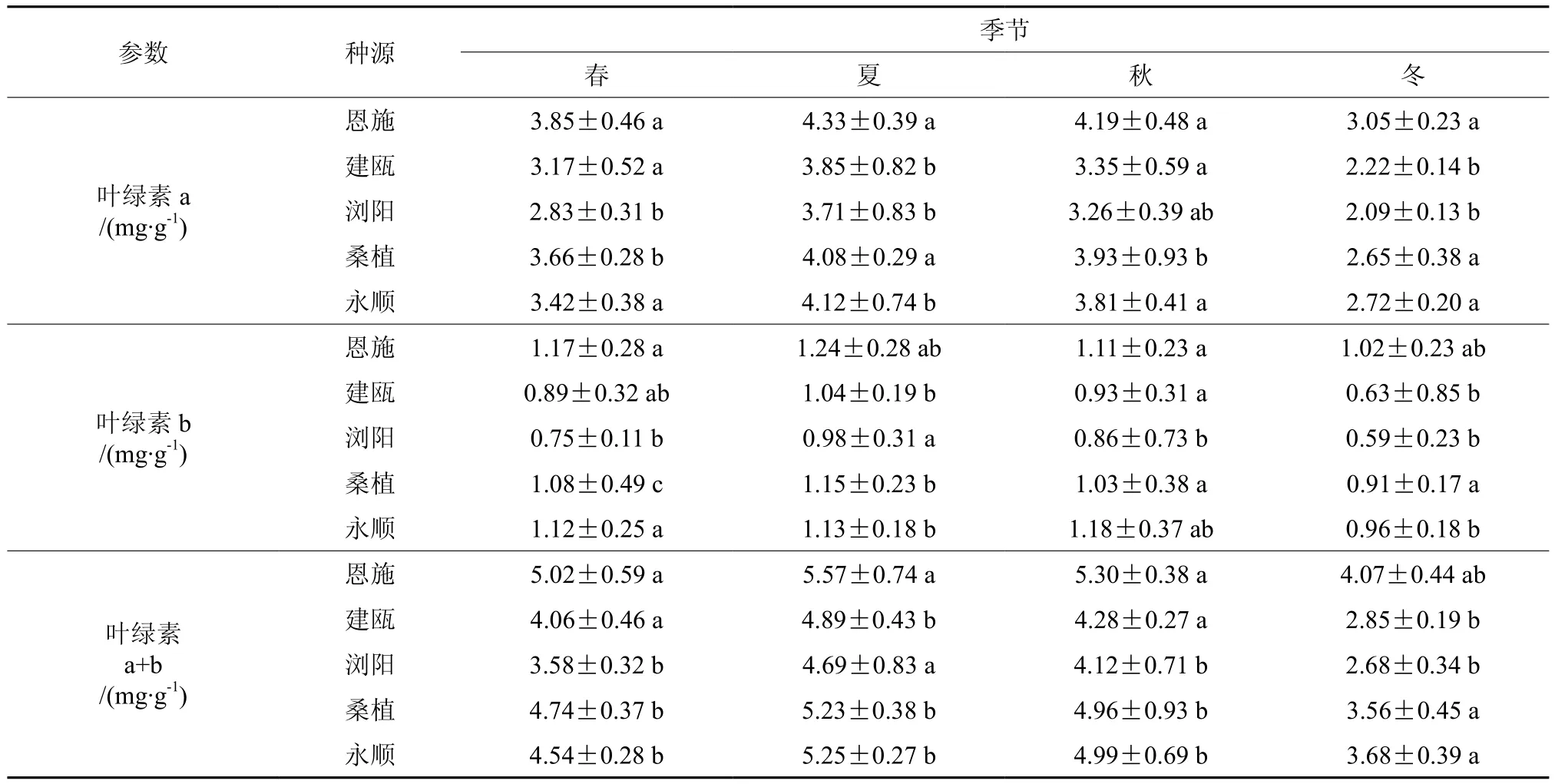
表4 不同种源钩栗叶绿素含量季节性变化Table 4 Seasonal change in the chlorophyll content of Castanopsis tibetana of different provenances
3 结论与讨论
(1)5个种源钩栗的净光合速率(Pn)日变化在春季、夏季和秋季3个季节中呈现为“双峰”曲线,在冬季则呈现为“单峰”曲线, 4个季节钩栗的日均Pn值均表现为夏季>秋季>春季>冬季,5个钩栗种源的光合能力强弱顺序为:恩施>桑植>永顺>浏阳>建瓯。Gs日变化曲线在春季、夏季和秋季均表现为“N”型,在冬季与Pn的日变化规律一致,Ci日变化规律与Pn日变化规律相反,Tr日变化规律与Pn日变化规律一致。有研究表明,如果Ci和Pn的变化方向一致,且Gs降低时Pn值也降低,则主要是由气孔因素造成;如果Ci和Pn变化方向相反,且随着Gs值的增大Pn值反而降低,则主要是由非气孔因素造成的,即叶肉细胞同化能力的降低引起的[14]。可见钩栗的光合“午休”现象主要是由非气孔因素造成的。
(2)5个种源钩栗的LCP、LSP、AQY、Rd和Pnmax值均表现为夏季>秋季>春季>冬季,且夏季光合优势明显。恩施种源的LCP、LSP、AQY、Rd和Pnmax在4个季节中均表现为最大。光是影响植物光合作用的重要因子,植物对光能利用能力的高低影响其光合速率,LCP和LSP分别体现了植物对弱光和强光的利用能力,直接反应了植物对光照条件的要求,AQY和Rd则体现了植物对光强变化的可塑性。不同季节光强的变化对钩栗光合特性有较大的影响。夏季PAR 最大,在4 个季节中钩栗叶片的 LSP、LCP、AQY、Rd和Pnmax均为夏季最高,且其他季节差异显著。可看出钩栗适宜在光照比较强的环境生长。而湖南冬季光强较弱,低的 LCP值能是钩栗更好的利用弱光进行光合作用,高的AQY能提高钩栗叶片对光能的利用效率,低的Rd也有助于减少呼吸的消耗,从而使钩栗在光合速率较低的时候积累较多有机物,增强其对弱光环境的适应,这说明钩栗的光合特性在一定程度上能适应光强的变化[15]。通常情况下,耐荫树种的Pnmax随着 PAR 的降低仅有略微的降低,表现出生理上的可塑性较差,因而光合能力较弱[16]。综上所述,钩栗光合作用的能力较强,随着 PAR 的降低,Pnmax值下降明显,表明其光合能力在生理上具有一定的可塑性,由此得出钩栗是阳性树种。
(3)5个种源钩栗的叶绿素含量均以冬季最低,夏季最高。同时恩施、桑植和永顺3个种源的叶绿素含量明显高于建瓯和浏阳种源。叶绿素可以将捕获的光能转化为化学能,是光合作用的光敏催化剂。而叶绿素a、b的含量和比例则是植物适应环境的重要指标[17]。夏季钩栗叶片叶绿素含量最高,表明夏季的强光环境促进了钩栗叶绿素的生物合成和积累,从而使叶绿素含量上升,这也是夏季钩栗光合能力最强的原因之一。同时,光合能力优势明显的恩施种源叶绿素含量也最大,这可能是因为在同样的环境下,植物通过增加叶绿素的量来适应环境,更好的捕获光能。
参考文献:
[1]祁承经,汤庚国.树木学(南方本)[M].北京:中国林业出版社,2010:224-225.
[2]林 敏,黄宗安.钩栗种群生命表分析[J].福建林业科技,2003, (2):9-13.
[3]王佩兰,许德禄,张 斌,等.钩栗种子形态特征及主要生理指标测定分析[J].种子,2013,(8):7-10,16.
[4]黄荣林,王 勇,刘晓蔚,等.钩栗不同基质和生根剂处理容器育苗对比试验研究[J].林业实用技术,2014,(7):41-44.
[5]陈 养.钩栗人工育苗技术研究[J].林业科技开发,2007,(3):89-90.
[6]Senevirathna A M W K, Stirling C M, Rodrigo V H L. Growth,photosynthetic performance and shade adaptation of rubber(Hevea brasiliensis) grown in natural shade[J]. Tree Physiology,2003, 23(10): 705-7l2.
[7]王路红,袁德义,沈广宁,等.花期喷硼对板栗光合特性和结实率的影响[J].经济林研究,2014,(2):58-61.
[8]王 强,金则新,郭水良,等.濒危植物长叶榧(Torreya jackii)的光合生理生态特性[J].生态学报,2014,(22):15-18.
[9]张 舟,吕芳德,王 森.不同枣品种光合特性的比较研究[J].中南林业科技大学学报,2014,34(8):78-81.
[10]谢 玲,刘卫东,冯斌义,等.不同株型观赏桃的生长及光合特性比较研究[J].经济林研究,2014,(4):103-109.
[11]王佩兰, 许德禄, 张 斌, 等. 钩栗种源间幼苗生长状况和光合特性比较研究[J].北方园艺,2014,(5):20-25.
[12]朱 慧, 马瑞君.入侵植物马缨丹(Lantana camara)及其伴生种的光合特性[J].生态学报,2009,(5):2701-2709.
[13]何维明, 董 鸣. 毛乌素沙地旱柳生长和生理特征对遮荫的反应[J].应用生态学报,2003,(2):175-178.
[14]Farquhar S P, Sharkey T D. Stomatal conductance and photosynthesis[J]. Annual Review of Plant Physiology, 1982, 33:317-345.
[15]王 荣.亚热带常绿阔叶林两个不同功能组树种幼苗对不同光环境的响应与适应[D].北京:中国林业科学研究院,2007.
[16]Reich P B, Walters M B, Tjoelker M G,et al. Photosynthesis and respiration rates depend on leaf and root morphology and nitrogen concentration in nine boreal tree species differing in relative growth rate[J]. Functional Ecology, 1998, 12, 395-405.
[17]刘悦秋,孙向阳,王 勇,等.遮荫对异株荨麻光合特性和荧光参数的影响[J].生态学报,2007,(8):3457-3464.
猜你喜欢
特产研究(2022年6期)2023-01-17
大众文艺(2022年14期)2022-08-09
绿色科技(2021年21期)2021-11-26
小小说月刊(2021年10期)2021-10-21
辽宁林业科技(2021年1期)2021-03-16
农家科技中旬版(2020年4期)2020-07-06
中学物理·高中(2016年12期)2017-04-22
商业文化(2016年32期)2016-06-05
考试周刊(2016年34期)2016-05-28
商业文化(2016年23期)2016-05-15
