
共表达HAC1基因对重组毕赤酵母分泌表达葡萄糖氧化酶的影响
2016-05-23 10:00:59堵国成江南大学生物工程学院江苏无锡214122粮食发酵工艺与技术国家工程实验室江南大学江苏无锡214122江南大学工业生物技术教育部重点实验室江苏无锡214122
食品与生物技术学报 2016年2期
关键词:葡萄糖氧化酶
顾 磊, 张 娟, 堵国成, 陈 坚(1.江南大学生物工程学院,江苏无锡214122;2.粮食发酵工艺与技术国家工程实验室,江南大学,江苏无锡214122;3.江南大学工业生物技术教育部重点实验室,江苏无锡214122)
共表达HAC1基因对重组毕赤酵母分泌表达葡萄糖氧化酶的影响
顾磊1,3,张娟1,3,堵国成1,3,陈坚*2,3
(1.江南大学生物工程学院,江苏无锡214122;2.粮食发酵工艺与技术国家工程实验室,江南大学,江苏无锡214122;3.江南大学工业生物技术教育部重点实验室,江苏无锡214122)
摘要:通过增强不可折叠蛋白质响应(UPR)信号途径以及研究不同培养温度下的影响,来提高重组葡萄糖氧化酶在毕赤酵母中的分泌表达。影响毕赤酵母外源蛋白表达的因素,主要为内质网中的折叠速率以及不可折叠蛋白积累造成的胞内胁迫压力。通过共表达HAC1基因对不可折叠蛋白信号通路进行调控,摇瓶中改造菌株PP-G-HAC1胞外酶活达到161 U/mL,相比于原始重组葡萄糖氧化酶菌株提高了34%。进一步研究不同温度下过量表达HAC1基因对菌株的生长和分泌外源蛋白的影响,菌株PP-G-HAC1在3 L发酵罐中28℃培养,酶活达到1 008 U/mL,胞外重组蛋白质达到14.43 g/L,相比原始菌株在相同条件下提高了3.12倍。
关键词:重组毕赤酵母;葡萄糖氧化酶;不可折叠蛋白响应;HAC1基因
葡萄糖氧化酶(EC 1.1.3.4,简称GOD)是生物领域中重要的工具酶。它是一种黄素蛋白,可以将β-D-葡萄糖通过分子氧传递电子转化为葡萄糖酸和过氧化氢。葡萄糖氧化酶被广泛应用于食品、饲料、医药等行业[1]。其中在医药和食品领域中的作用尤为重要。在医药领域中,可制成试剂盒及电极,用于血浆、尿液及脑脊液中葡萄糖的定量检测[2]。在食品领域中,可用于去除食品和饮料中残留的葡萄糖和氧,从而提高货架期和保质期,同时也是生产葡萄糖酸及其衍生物的重要转化用酶[3]。葡萄糖氧化酶及其产物和产物衍生物均具有广泛的市场和应用前景。大规模生产菌株常采用黑曲霉或者青霉,然而采用霉菌生产有其弊端,大量的杂蛋白质和伴生产物造成下游提取的困难,同时产酶量不高也成为大规模生产的主要瓶颈[4-6]。为了解决这些问题,近年来使用基因工程改造方法进行异源表达受到关注。其中酵母表达系统相较于大肠杆菌等原核表达系统能够较好地异源表达葡萄糖氧化酶[7-11]。酿酒酵母表达系统由于过糖基化修饰,使得重组葡萄糖氧化酶过糖基化情况严重,限制了葡萄糖氧化酶的应用[12]。毕赤酵母表达系统在这方面展现出优势,但国内外相关报导中,黑曲霉来源基因重组葡萄糖氧化酶在毕赤酵母中的最高产率为40 U/mL,仍然不能满足大规模生产的要求[11]。因此,如何提高重组葡萄糖氧化酶的产量成为十分重要的研究方向。
毕赤酵母(Pichia pastoris)是一种广泛应用于分泌表达外源蛋白的真核系统。尽管已有多种外源蛋白质在该系统中成功表达,但不意味着所有的案例都达到预期的效果,毕赤酵母系统中表达外源源蛋白存在着很大的差异。最近的研究发现,在毕赤酵母表达系统中,内质网中未折叠蛋白或错误折叠蛋白的积聚对于外源蛋白的分泌表达量有很大的影响[13-14]。经常采用的更换强启动子或者增加外源蛋白基因拷贝数的策略,使得大量属于外源蛋白质的初生肽链在内质网中滞留,不能成功分泌表达,甚至影响了宿主菌株的生长[15-17]。HAC1基因编码的Hac1p是酵母内质网中不可折叠蛋白响应(UPR)机制的激活调控因子,能够调控分泌表达途径中的下游靶基因,这些基因包含了各种分子伴侣,折叠因子,内质网中参与糖基化控制,以及其他胞内胁迫条件下调控因子的编码基因[18-19]。在作者所在研究室前期的研究中,筛选得到的高产葡萄糖氧化酶黑曲霉菌株Aspergillus niger BBE11721,扩增其葡萄糖氧化酶基因,构建了毕赤酵母基因工程菌株Pichia pastoris GS115-pPIC9K-GOD(保藏编号为CCTCC NO:M 2012266)。作者在此基础上,共表达了HAC1基因,并且进一步研究不同培养温度下共表达HAC1基因对菌株的生长和分泌外源蛋白质的影响。
1 材料与方法
1.1材料
1.1.1菌株与质粒菌株与质粒:均为作者所在实验室购买及保存。研究中使用及构建所得菌株和质粒见表1。
1.1.2试剂PrimeSTAR HS DNA聚合酶、T4连接酶、PCR产物纯化试剂盒、限制性内切酶等:大连宝生物公司;SDS-PAGE凝胶试剂盒、蛋白质电泳marker:购自碧云天公司;培养基所用酵母膏、蛋白胨:Oxoid公司;引物及测序:均由上海生工有限公司完成。其他常规试剂为国产分析纯。
1.2培养基
1.2.1 LB培养基葡萄糖20 g/L,蛋白胨20 g/L,酵母膏10 g/L。
1.2.2毕赤酵母培养基毕赤酵母培养用的培养基(YPD、MD、BMGY和BMMY)配方参照Invitrogen公司操作手册。
1.2.3分批发酵BSM培养基85%磷酸26.7 mL/L,CaSO4·H2O 0.93 g/L,K2SO418.2 g/L,MgSO4·7H2O 14.9 g/L,KOH 4.13 g/L,甘油40.0 g/L,PTM1 4.35 mL/L(PTM1配方参照Invitrogen公司操作手册)。
1.2.4补料培养基50%甘油(含12 mL/L PTM1)。

表1 本研究中所使用的菌株和质粒Table 1 Strains and plasmids used in this study
1.2.5诱导培养基100%甲醇(含12 mL/L PTM1)。
1.3培养方法
1.3.1摇瓶发酵大肠杆菌采用LB培养基培养。毕赤酵母采用YPD、BMGY和BMMY等培养。其中毕赤酵母诱导培养,在BMGY中培养至OD值为1.2~1.5时,将酵母细胞转入BMMY培养基中诱导,每12小时加入总体积1%的甲醇进行持续诱导。
1.3.2 3 L发酵罐高密度发酵将YPD中菌液按10%接种体积分数接入3 L全自动发酵罐(LiFlus GM BioTRON,Korea)中。以50%氨水和磷酸溶液控制pH 5.5,温度30℃,调节搅拌转速(500~1 000 r/min)和通气量(2~5 L/min)维持溶氧30%以上。当甘油耗尽(DO迅速上升,且DO>60%时)开始流加补料培养基。当菌体达到一定质量浓度后(60、80、100 g/L),停止补料。待甘油再次耗尽,继续保持基质匮乏状态1 h,DO>60%后,开始流加诱导培养基,并把诱导温度降低至22℃或者28℃诱导表达外源蛋白质。甲醇浓度控制在2%,使用FC2002华东理工大学在线甲醇在线检测与控制器控制。
1.4方法
1.4.1基因PCR扩增葡萄糖氧化酶GOD基因通过GeneBank登录号ACB30370.1设计引物:GODF:TCCTACGTAAATGGCATTGAAGCCAGCCT;GODR:TTGCGGCCGCTCACTGCATGGAA GCATAATCT TC(下划线处碱基为酶切位点)。以黑曲霉A. niger BBE11721的cDNA为模版。PCR扩增条件:95℃预变性5 min;98℃变性10 s,57℃退火30 s,72℃延伸2 min(30个循环);72℃延伸10 min。
HAC1基因以GeneBank公布的基因编号8196642设计引物:HAC1 -F:GGGTTCGAAAT GCCCGTAGATTCTTCTC;HAC1-R:TTGCGGCCGCC TATTCCTGGAAGAATACAAAGTC(下划线处碱基为酶切位点)。以毕赤酵母P. pastoris GS115的cDNA为模版。PCR扩增条件:95℃预变性5 min;98℃变性10 s,55℃退火30 s,72℃延伸1 min(30个循环);72℃延伸10 min。PCR扩增产物纯化参照试剂盒说明书。
1.4.2重组质粒构建将上述PCR片断纯化后连接到pMD19-T克隆载体上,转化E. coli DH5α,提取质粒,将筛选得到的阳性克隆子进行测序鉴定。葡萄糖氧化酶GOD克隆载体采用SnaBI和NotI进行双酶切,其产物纯化后与表达载体pPIC9k相连接,筛选阳性克隆验证,获得重组质粒pPIC9k/ GOD。HAC1基因克隆载体采用AsuII和NotI进行双酶切,其产物纯化后与表达载体pGAPZ相连接,筛选阳性克隆验证,获得重组质粒pGAPZ/HAC1。
1.4.3酵母转化和重组子筛选将构建的表达载体pPIC9k/GOD使用BglII进行酶切线性化,pGAPZ/HAC1使用SacI进行酶切线性化。pPIC9k/ GOD线性化质粒电转入毕赤酵母GS115感受态细胞,使用电转仪(Gene Pulser electroporator,Bio-Rad),电转条件:2000 V,200 Ω,25 μF。使用RT-qPCR方法测定挑选出的阳性克隆中的GOD拷贝数,获得PP-GOD重组菌株。将pGAPZ/HAC1线性化质粒电转入毕赤酵母PP-GOD感受态细胞,使用RT-qPCR方法测定挑选出的阳性克隆中的HAC1基因拷贝数,获得PP-G-HAC1重组菌株。
1.4.4 TRIzol法提取RNA离心收集菌体并置于液氮终止反应,RNA的抽提使用RNA抽提试剂盒(Trizol regent),参照试剂盒说明书进行。
1.4.5 RT -qPCR将获得的RNA使用cDNA Synthesis kit(Takara)进行反转录,然后将反转录cDNA作为模版。荧光定量PCR所用试剂盒为SYBR○R
Premix Ex TaqTMkit(Takara),反应仪器为LightCycler 480 II Real-time PCR instrument(Roche Applied Science,Mannheim,Germany)。反应条件为95℃30 s,40个循环。95℃5 s,60℃20 s最后50 ℃30 s冷却。基因转录变化计算方法为2-△△CT,内参基因为GAPDH。选取3个生物学平行样,取平均值,所用定量PCR引物见表2。

表2 本研究中所使用定量引物Table 2 Oligonucleotides used in this study
1.4.6葡萄糖氧化酶活性测定测定方法参照文献[6]。活力定义单位:每分钟转化1 μmol葡萄糖为葡萄糖酸和过氧化氢所需的酶量为一个单位,以U/mL表示。
1.4.7 AOX氧化酶活性测定测定方法参照文献[20]。活力定义单位:相同菌浓下,每分钟产生1 μmol过氧化氢所需的酶量为一个单位,以U/gDCW表示。
2 结果与讨论
2.1葡萄糖氧化酶重组菌株及HAC1共表达重组菌株的构建
分别以A. niger BBE11721为模版,配合以GOD-F/GOD-R为引物;以毕赤酵母P. pastoris GS115的cDNA为模版,配合以HAC1-F/HAC1-R和为引物,按照方法1.4.1进行基因克隆,分别PCR扩增得到1 800左右和1 000左右的特异性条带,见图1。测序结果表明,PCR扩增所得GOD基因全长1 749 bp,编码583个氨基酸;PCR扩增所得HAC1基因全长996 bp,编码332个氨基酸,两段基因大小和序列均与预期相符。
为在毕赤酵母表达体系中顺利将GOD表达并分泌至胞外,采用毕赤酵母表达载体上的信号肽αfactor以实现在毕赤酵母表达体系中的分泌。并使用带有AOX1强启动子的pPIC9k表达载体,按照方法1.4.2构建得到pPIC9k/GOD重组载体。同时,为在毕赤酵母表达体系中胞内表达调控子Hac1p,作者采用了带有GAP启动子的表达载体pGAPZα,并通过使用AsuII和NotI双酶切方式去除了质粒上的信号肽α-factor,按照方法1.4.2构建得到了pGAPZ/HAC1重组载体。此后,将pPIC9k/GOD线性化质粒电转入毕赤酵母GS115感受态细胞,获得PP-GOD重组菌株。将pGAPZ/HAC1线性化质粒电转入毕赤酵母PP-GOD感受态细胞,获得PP-GHAC1重组菌株。重组菌中的葡萄糖氧化酶基因GOD和HAC1基因拷贝数使用RT-qPCR的方法测定,参照文献[21]。本研究中所使用菌株中基因拷贝数见表3。因为AOX1与GAP基因同时存在于转化的质粒和毕赤酵母的基因组中,因此表中所示拷贝数为测得拷贝数减去基因组中已存在的一个拷贝数。
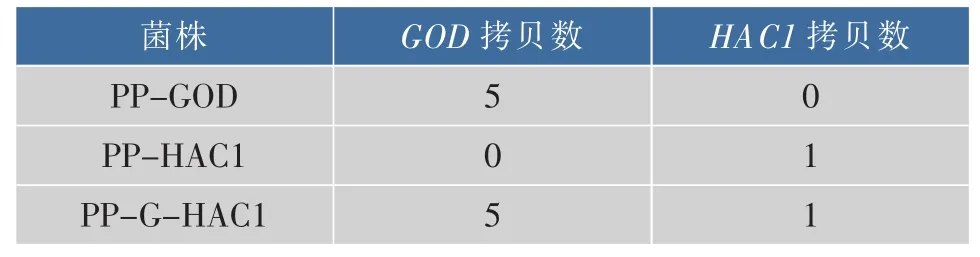
菌株 GOD拷贝数 HAC1拷贝数PP-GOD 5 0 PP-HAC1 0 1 PP-G-HAC1 5 1
2.2共表达HAC1基因对葡萄糖氧化酶GOD表达的影响
在毕赤酵母中过量表达外源蛋白质会带给分泌途径负担,大量的未折叠肽链或错误折叠蛋白质是影响外源分泌表达的主要原因[22]。在本研究中,作为UPR效应的激活因子Hac1p的编码基因HAC1,通过过量共表达HAC1基因考察了该基因对葡萄糖氧化酶GOD在P. pastoris中表达的影响。如图2所示,改造重组菌株PP-G-HAC1在摇瓶中能够分泌更多的胞外GOD,培养120 h后胞外GOD活性达到161 U/mL,相比于原始菌株PP-GOD提高了37%。同时通过过量共表达HAC1基因,使得GOD的单位生产率从PP-GOD菌株的0.42 mg/(g·h)提高到PP-G-HAC1菌株的0.52 mg/(g·h)。如表4所示,改造重组菌PP-G-HAC1在胞外GOD蛋白质质量浓度和GOD酶得率(重组酶量/细胞干质量DCW)均有所提高,分别从原始菌株的1 059 mg/L 与60.2 mg/g提高到1 439 mg/L与74.2 mg/g。结果表明,共表达调控因子编码HAC1基因能够有效的改善GOD在毕赤酵母中的分泌表达。
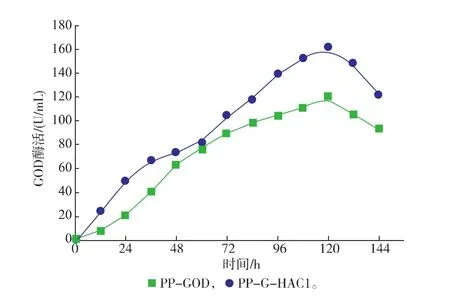
图2 摇瓶中过量表达HAC1对发酵生产葡萄糖氧化酶的发酵过程曲线Fig. 2 Effects of Co-Expression of HAC1 on the GOD secretion in P. pastoris in shake flasks
2.3 3 L发酵罐中不同培养温度下共表达HAC1基因对发酵产GOD的影响
外源蛋白质表达分泌的过程中,环境因素对宿主的生理和蛋白质的生产率有重要影响。在毕赤酵母表达体系中,温度被认为直接影响蛋白质生产能力和宿主代谢功能。不同的培养温度能够改变蛋白质的合成、折叠和分泌速率,同时对宿主内部蛋白质分泌途径中造成胁迫应激反应也有重要的影响。温度对蛋白质分泌的影响是基于宿主生长情况,蛋白质合成和蛋白质在胞内积聚造成的胁迫压力等各因素的综合体现[23]。毕赤酵母的最适生长温度为28~30℃,而研究表明,降低培养温度有利于提高部分外源蛋白质的分泌[24]。虽然温度的降低使得毕赤酵母细胞的生长代谢速率降低,但能够减少由于蛋白质过量合成分泌过程引起的细胞衰亡,见表4。作者采用3 L发酵罐进行培养,进一步对不同改造菌株在不同培养温度下的发酵情况进行研究。目的在于研究共表达HAC1基因在不同温度下对外源分泌蛋白质的影响,以及在改进培养温度的发酵条件下进一步提高葡萄糖氧化酶GOD的酶活。
2.3.1重组菌PP-GOD在3 L发酵罐中不同培养温度下对发酵产GOD的影响考察了3 L发酵罐中不同培养温度(22℃和28℃)、不同重组菌发酵产GOD的影响。如图3所示,重组菌PP-GOD在22℃时DCW最高达到179 g/L,然而在28℃DCW最高仅能达到134 g/L。同时,重组菌PP-GOD在22℃最高GOD活性达到427.6 U/mL,相较于28℃培养条件GOD活性323 U/mL提高了25%。这些结果表明,甲醇诱导期的低温诱导能够有效提高重组菌PP-GOD的GOD产量。
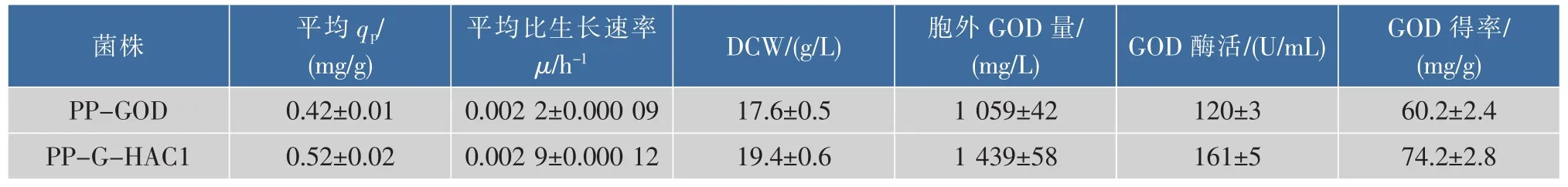
表4 摇瓶中过量表达HAC1对工程菌发酵参数的影响Table 4 Effects of co-expression of HAC1 on heterologous protein production of GOD in shake flasks
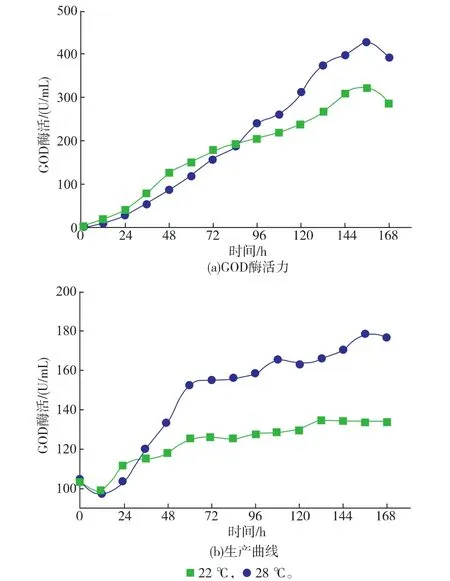
图3 3 L发酵罐中不同培养温度下重组菌PP-GOD产GOD的发酵曲线Fig. 3 Fermentation properties of the GOD over -expressing strain PP-GOD in 3 L fermentor
2.3.2重组菌PP-G-HAC1在3 L发酵罐中不同培养温度下对发酵产GOD的影响考察3 L发酵罐中改造重组菌PP-G-HAC1菌株在过量共表达HAC1基因发酵产GOD的影响,在22℃培养条件下,产GOD的能力并没有因为降低温度而有所提高。如图4所示,经过156 h的诱导,改造重组菌PP-G-HAC1在28℃时胞外GOD酶活能够达到1 008 U/mL,同时比酶活70 kU/g,菌体干重DCW最高达到300 g/L。而在3 L发酵罐中,22℃下GOD酶活最高达到699 U/mL,胞外GOD酶量12.02 g/L,比28℃培养条件下有所降低。
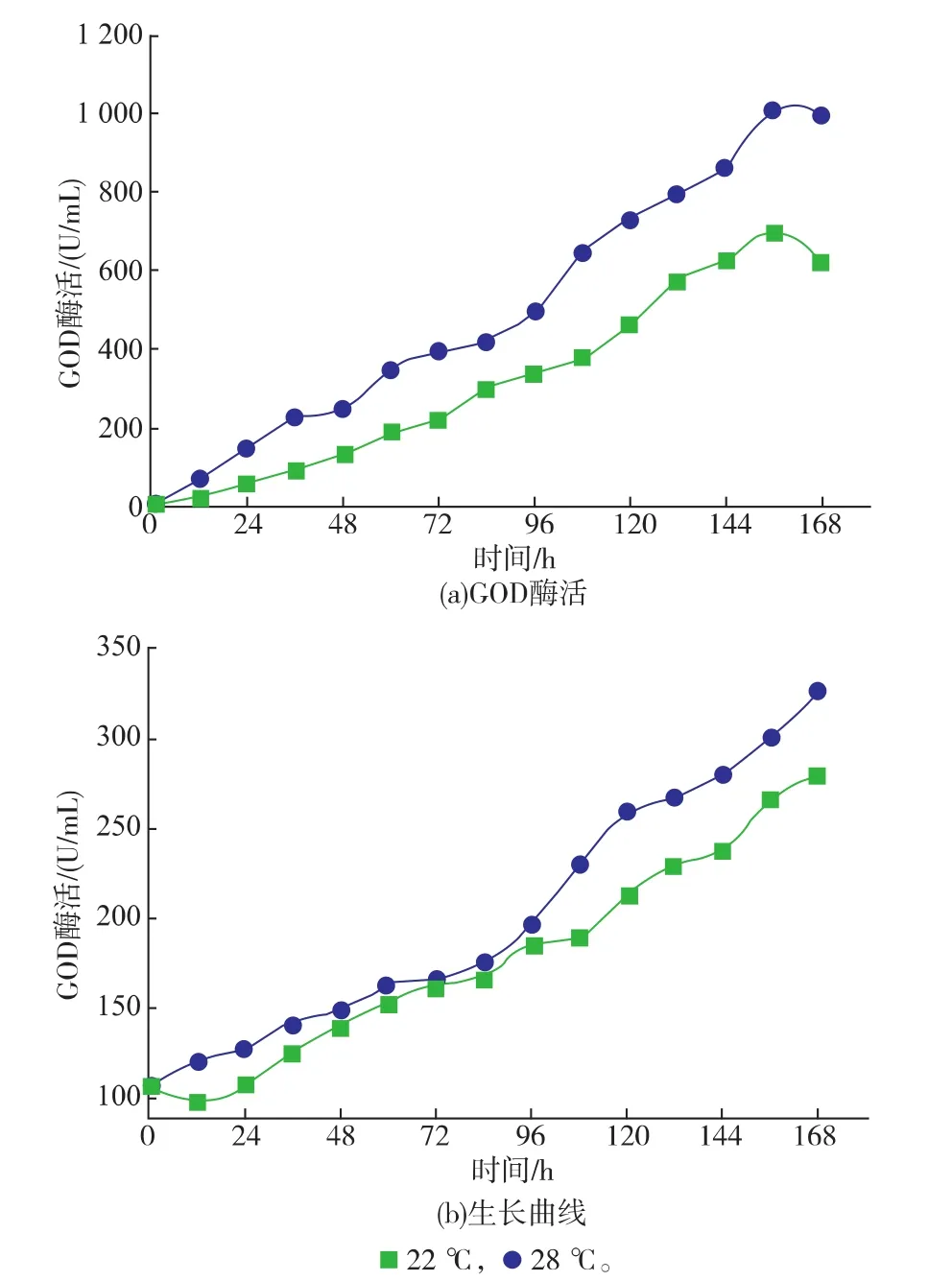
图4 3 L发酵罐中不同温度下重组菌PP-G-HAC1产GOD的发酵曲线Fig. 4 Effect of different cultivation temperatures on GOD secretion in engineered secretion strain PPG-HAC1 in 3 L fermentor
进一步考察了3 L发酵罐中不同培养温度对不同重组菌PP-GOD与PP-G-HAC1发酵产GOD的影响。综合各情况下发酵参数的比较,如表5所示,重组菌PP-G-HAC1在28℃下对产GOD更有利,比酶活69 854 U/g,GOD生产率6.46 U/(mL·h),胞外GOD酶量14.43 g/L,均相较于22℃培养条件下有明显的提高(比酶活58 153 U/g,GOD生产率4.48 U/(mL·h),胞外GOD酶量12.02 g/L)。如图5所示,PP-G-HAC1菌株在不同温度下,诱导156 h胞外蛋白SDS-PAGE图,显示28℃条件下的GOD条带要较22℃更为明显。
同时,从表5中发酵参数发现,重组菌PP-GOD在28℃下的发酵参数要明显低于重组菌PP-GHAC1的发酵参数。通过对AOX醇氧化酶活性的检测,发现在相同条件下不同的改造菌株中差异较大。P. pastoris对外源蛋白质的表达受到AOX1启动子调控,AOX醇氧化酶是甲醇代谢途径中第一个关键酶,受甲醇的诱导启动[25]。如图6所示,PP-GHAC1菌株中的AOX活性曲线高于PP-GOD菌株,并且最大AOX活性达到8.8 U/g,相较于PP-GOD菌株中的5.1 U/g有明显提高。
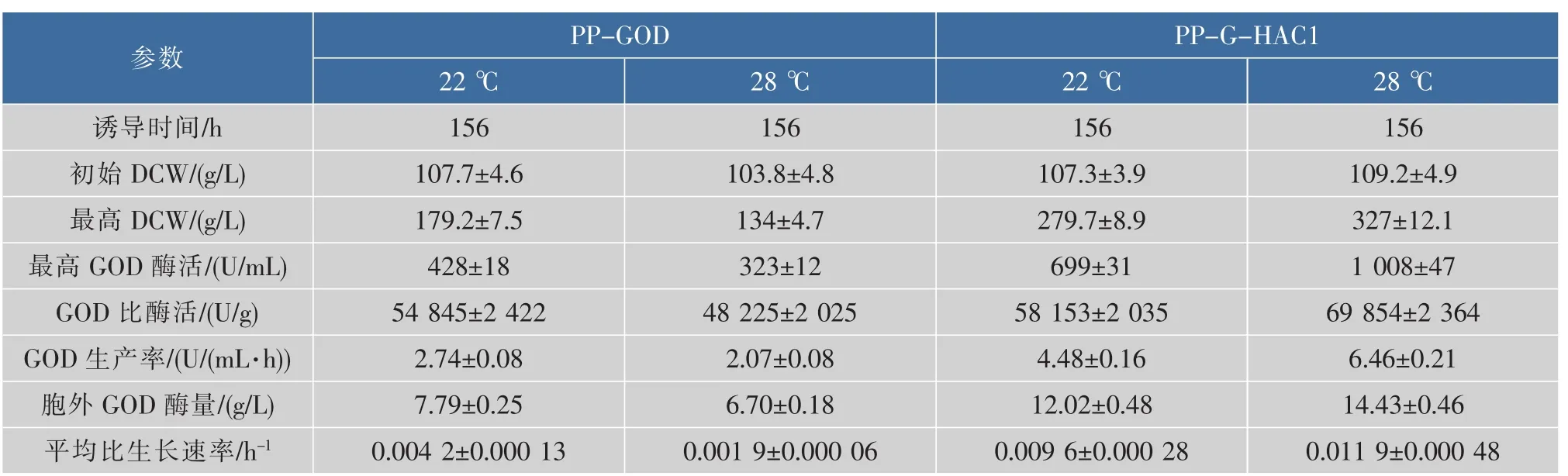
表5 不同重组菌在不同培养温度下发酵产GOD的参数Table 5 Comparison of parameters for GOD production under different cultivation temperatures
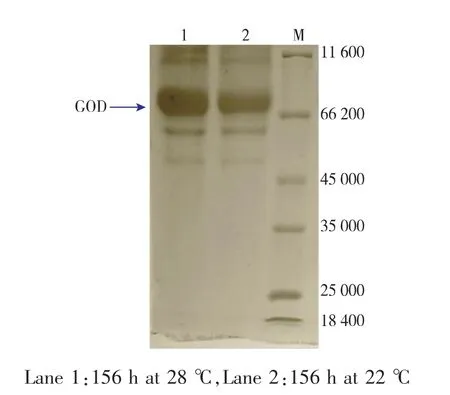
图5 不同培养温度下PP-G-HAC1菌株156 h时胞外蛋白SDS-PAGE图Fig. 5 SDS -PAGE of fermentation supernatant of the engineered secretion strain PP-G-HAC1 in 3 L fermentor at different temperatures.
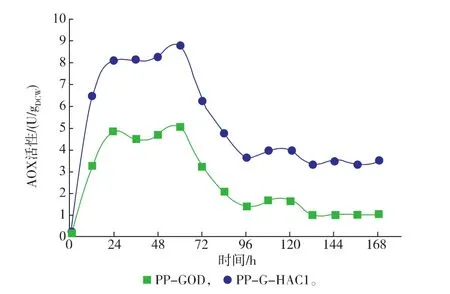
图6 3 L发酵罐中28℃下重组菌PP-G-HAC1与PPGOD产GOD的AOX活性曲线Fig. 6 Effects of co-expression of HAC1 on AOX activity in 3 L fermentor at 28℃
通过在不同温度下不同重组菌株发酵GOD影响的研究,发现在原始菌株PP-GOD中,降低诱导期培养温度至22℃能够有效地提高GOD的产量,菌体生长较28℃时有明显提高,说明较高的培养温度使得蛋白质过量合成分泌过程引起的胁迫反应影响了细胞的生长,导致GOD产量的降低。而在共表达HAC1基因的菌株PP-G-HAC1中,28℃培养条件较22℃培养条件下GOD的产量有进一步的提高,两种菌株在22、28℃下的产外源蛋白质差异,说明了共表达HAC1基因能够有效减缓蛋白质过量表达情况下胞内的胁迫压力,增强宿主细胞的生长情况,在毕赤酵母最适生长温度28℃下进一步提高GOD的产量。
2.4 28℃培养温度下UPR下游靶基因基因转录水平变化
HAC1基因编码的Hac1p作为UPR途径的激活调节因子,能够调控UPR途径中的许多功能基因。毕赤酵母内的UPR典型下游靶基因包含了折叠基因(ERO1,KAR2),转运基因(SEC18,SEC53,SSA4,VPS17),ERAD基因(HRD1,UBC1)和胁迫调控基因(GCN4,MSN2)等典型编码基因。为了进一步考察这些编码基因在28℃培养条件下不同菌株中的转录水平变化,作者采用RT-qPCR技术对这些基因的表达量进行了进一步分析,结果见图7。
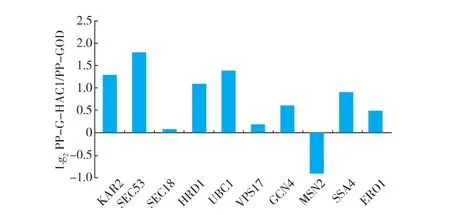
图7 28℃培养温度下UPR下游靶基因基因转录水平变化Fig. 7 Comparison of marker genes transcriptional levels
PP-G-HAC1菌株相较于PP-GOD菌株明显上调的基因包括:内质网折叠基因KAR2,上调1.3倍;参与内质网蛋白修饰与糖基化与转运基因SEC53,上调1.7倍;参与内质网蛋白降解基因HRD1,上调1.1倍;调节选择性降解非正确折叠蛋白基因UBC1,上调1.4倍。这些直接参与分泌途径的基因的上调有效地说明了PP-G-HAC1菌株在过量共表达HAC1基因的情况下胞内转运分泌外源蛋白GOD的能力更强。然而胁迫调控基因MSN2有所下调,同样属于胁迫调控基因GCN4基因却较PP-GOD菌种有所上调。GCN4基因编码的调控因子能够调控一些与氨基酸合成相关基因的表达。GCN4编码蛋白由内质网上胁迫环境激活,被认为是由HAC1基因调控的UPR应答机制中关键下游调控蛋白。GCN4的同源基因作为内质网胁迫与氧化胁迫的调控桥梁,能够调节还原型谷胱甘肽的生物合成,用来改善细胞由于内质网和内质网降解蛋白机制中造成的氧化蛋白折叠积累的活性氧形成的活性降低现象[26]。PP-G-HAC1菌株中GCN4基因的上调说明了胞内由于外源蛋白GOD大量分泌表达的情况下,为抵御活性氧(ROS)的形成进行了更有效的抵御。
在前期的实验中,发现PP-GOD菌株28℃培养的GOD产量较22℃下有明显的降低,造成这一现象的原因是胞内胁迫压力引起宿主细胞生长能力和蛋白质合成速率的下降。胁迫压力由内质网中未折叠和错误折叠蛋白质的大量积累造成,一旦发生这种情况会造成3种常见的结果[27]:1)新生肽链向内质网的转运减少;2)内质网折叠能力的加强;3)当前两者不能解决胁迫压力,就会造成宿主功能性失常,宿主菌体就会死亡。宿主细胞对应过量表达蛋白质的胁迫情况下,自身的UPR途径会被激活,作为一种防御机制来抵御胞内的非正常胁迫环境。通过对UPR途径下游典型编码基因的转录水平考察,发现在PP-G-HAC1中过量共表达HAC1基因能够显著增强UPR信号途径的操纵能力。在相同培养条件下,同原始菌株PP-GOD相比,内质网中处理新生肽链折叠的编码基因明显上调,并且内质网降解途径ERAD等功能基因也显著上调,这些功能基因转录水平的增强减轻了内质网胁迫压力。同时调节胞内ROS水平的氧化胁迫调控基因GCN4的转录水平也明显上调。这说明了在28℃培养温度条件下共表达HAC1基因,能够有效地提高宿主细胞的蛋白质折叠和分泌能力,同时增强胞内胁迫压力的抵御能力,从而提高外源GOD的产量。
3 结语
作者分析了毕赤酵母共表达中UPR中关键基因HAC1对于GS115菌株发酵产葡萄糖氧化酶GOD的影响,改造菌株PP-G-HAC1在3 L发酵罐中28℃培养,酶活达到1 008 U/mL,胞外重组蛋白质达到14.43 g/L,相比原始菌株在相同条件下提高了3.12倍。为了进一步提高葡萄糖氧化酶在毕赤酵母中的发酵生产,解析毕赤酵母系统中合成分泌外源蛋白质通路的代谢网络,以及通过代谢工程改造增强原生肽链修饰折叠,降低未折叠蛋白质对内质网的胁迫,减少细胞负担的同时增强外源蛋白质分泌,将成为我们日后继续深入研究的重点。
参考文献:
[ 1 ] Bankar S B,Bule M V,Singhal R S,et al. Glucose oxidase-an overview[J]. Biotechnol Adv,2009,27:489-501.
[ 2 ] Kim S H,Lee S M,Kim D U,et al. Enzyme-based glucose biosensor using a dye couple system [J]. Dyes Pigments,2001,49:103-108.
[ 3 ] Klein J,Rosenberg M,Markoš J,et al. Biotransformation of glucose to gluconic acid by Aspergillus niger-study of mass transfer in an airlift bioreactor[J]. Biochem Eng J,2002,10:197-205.
[ 4 ] Petruccioli M,Federici F,Bucke C,et al. Enhancement of glucose oxidase production by Penicillium variabile P16 [J]. Enzyme Microb Tech,1999,24:397-401.
[ 5 ] Fiedurek J,Gromada A. Production of catalase and glucose oxidase by Aspergillus niger using unconventional oxygenation of culture[J]. J Appl Microbiol,2000,89:85-89.
[ 6 ] Bankar S B,Bule M V,Singhal R S,et al. Optimization of Aspergillus niger fermentation for the production of glucose oxidase[J]. Food Bioprocess Technol,2009,2:344-352.
[ 7 ] Malherbe D,Du Toit M,Otero RC,et al. Expression of the Aspergillus niger glucose oxidase gene in Saccharomyces cerevisiae and its potential applications in wine production[J]. Appl Microbiol Biotechnol,2003,61:502-511.
[ 8 ] Rocha SN,Abrahão-Neto J,Cerdán ME,et al. Heterologous expression of glucose oxidase in the yeast Kluyveromyces marxianus [J]. Microb Cell Fact,2010,9:4-12.
[ 9 ] Yamaguchi M,Tahara Y,Nakano A,et al. Secretory and continuous expression of Aspergillus niger glucose oxidase gene in Pichia pastoris[J]. Protein Expr Purif,2007,55:273-278.
[10] Crognale S,Pulci V,Brozzoli V,et al. Expression of Penicillium variabile P16 glucose oxidase gene in Pichia pastoris and characterization of the recombinant enzyme[J]. Enzyme Microb Tech,2006,39:1230-1235.
[11] Guo Y,Lu F,Zhao H,et al. Cloning and heterologous expression of glucose oxidase gene from Aspergillus niger Z-25 in Pichia pastoris[J]. Appl Biochem Biotech,2010,162:498-509.
[12] Frederick K,Tung J,Emerick R,et al. Glucose oxidase from Aspergillus niger. Cloning,gene sequence,secretion from Saccharomyces cerevisiae and kinetic analysis of a yeast-derived enzyme[J]. J Biol Chem,1990,265:3793-802.
[13] Sitia R,Braakman I. Quality control in the endoplasmic reticulum protein factory[J]. Nature,2003,426:891-894.
[14] Damasceno LM,Huang C-J,Batt CA. Protein secretion in Pichia pastoris and advances in protein production[J]. Appl Microbiol Biotechnol,2012,93:31-39.
[15] Hohenblum H,Gasser B,Maurer M,et al. Effects of gene dosage,promoters,and substrates on unfolded protein stress of recombinant Pichia pastoris[J]. Biotechnol Bioeng,2004,85:367-375.
[16] Li Z,Moy A,Gomez SR,et al. An improved method for enhanced production and biological activity of human secretory leukocyte protease inhibitor(SLPI)in Pichia pastoris[J]. Biochem Bioph Res Co,2010,402:519- 524.
[17] Shen Q,Wu M,Wang H-B,et al. The effect of gene copy number and co-expression of chaperone on production of albumin fusion proteins in Pichia pastoris[J]. Appl Microbiol Biotechnol,2012,96:763-772.
[18] Valkonen M,Penttilä M,Saloheimo M. Effects of inactivation and constitutive expression of the unfolded-protein response pathway on protein production in the yeast Saccharomyces cerevisiae[J]. Appl Environ Microbiol,2003,69:2065-2072.
[19] Guerfal M,Ryckaert S,Jacobs PP,et al. The HAC1 gene from Pichia pastoris:characterization and effect of its overexpression on the production of secreted,surface displayed and membrane proteins[J]. Microb Cell Fact,2010,9:49-12.
[20] Suye SI,Ogawa A,Yokoyama S,et al. Screening and identification of Candida methanosorbosa as alcohol oxidase producing methanol using yeast[J]. Agric Biol Chem,1990,54:1297-1298.
[21] Sha C,Yu X-W,Li F,et al. Impact of gene dosage on the production of lipase from Rhizopus chinensis CCTCC M201021 in Pichia pastoris[J]. Appl Biochem Biotech,2013,169:1160-1172.
[22] Hohenblum H,Gasser B,Maurer M,et al. Effects of gene dosage,promoters,and substrates on unfolded protein stress of recombinant Pichia pastoris[J]. Biotechnol Bioeng,2004,85:367-375.
[23] Gasser B,Maurer M,Rautio J,et al. Monitoring of transcriptional regulation in Pichia pastoris under protein production conditions[J]. BMC Genomics,2007,8:179.
[24] Wang Y,Wang Z H,Xu Q L,et al. Lowering induction temperature for enhanced production of polygalacturonate lyase inrecombinant Pichia pastoris[J]. Process Biochem,2009,44:949-954.
[25] Jungo C,Rerat C,Marison IM,et al. Quantitative characterization of the regulation of the synthesis of alcohol oxidase and of the expression of recombinant avidin in a Pichia pastoris Mut+strain[J]. Enzyme Microb Technol,2006,39:936-944.
[26] Harding H P,Zhang Y,Zeng,et al. An integrated stress response regulates amino acid metabolism and resistance to oxidative stress[J]. Mol Cell,2003,11:619-633.
[27] Ron D,Walter P. Signal integration in the endoplasmic reticulum unfolded protein response[J]. Nat Rev Mol Cell Bio,2007,8:519-529.
Effects of Co-Expression of HAC1 on Glucose Oxidase Production in Recombinant Pichia pastoris
GU Lei1,3,ZHANG Juan1,3,DU Guocheng1,3,CHEN Jian*2,3
(1. School of Biotechnology,Jiangnan University,Wuxi 214122,China;2. National Engineering Laboratory for Cereal Fermentation Technology,Jiangnan University,Wuxi 214122,China;3. Key Laboratory of Industrial Biotechnology,Ministry of Education,Jiangnan University,Wuxi 214122,China)
Abstract:In this work,the enhancement of unfolded protein response transcription pathway at different cultivation temperatures were integrated as a novel protein secretion strategy to improve extracellular enzyme concentration and specific productivity of heterologous glucose oxidase(GOD)in Pichia pastoris. Efficient production of heterologous proteins in P. pastoris was often limited by metabolic and intrinsic stresses such as folding rate and ER stress by accumulation of unfolded protein. The results showed that enzymatic activity of engineered secretion strain PP-G-HAC1,co-expression of UPR activating transcription factor Hac1p from P. pastoris,was up 161 U/mL in shake flasks,34% higher compared to control strain. Furthermore,the effects of different cultivation
E-mail:jchen@jiangnan.edu.cntemperatures(22、28℃)were also investigated. The highest GOD activity and extracellular concentration at 28℃in PP-G-HAC1 were 1,008 U/mL and 14.43 g/L,respectively,3.12-fold increase than that of control strain.
Keywords:Pichia pastoris,glucose oxidase,unfolded protein response,HAC1
*通信作者:陈坚(1962—),男,江苏无锡人,工学博士,教授,博士研究生导师,主要从事发酵工程和食品生物技术方面的研究。
基金项目:国家自然科学基金重大项目(31470160);国家863计划项目(2011AA100905);国家973计划项目(2013CB733902);博士后科学基金项目(114957,2013M540538);江苏省自然科学基金项目(BK2012553)。
收稿日期:2014-08-29
中图分类号:Q 555
文献标志码:A
文章编号:1673—1689(2016)02—0113—10
猜你喜欢
中国饲料(2021年5期)2021-12-05 15:05:21
河南工业大学学报(自然科学版)(2021年1期)2021-03-18 12:07:18
饲料博览(2019年8期)2019-02-13 16:46:10
饲料博览(2019年7期)2019-02-12 22:28:15
陶瓷学报(2019年5期)2019-01-12 09:17:42
猪业科学(2018年5期)2018-07-17 05:55:54
饲料博览(2017年2期)2017-04-04 10:56:02
大科技(2016年27期)2016-07-13 06:09:30
食品与发酵工业(2015年1期)2015-12-16 08:05:22
动物营养学报(2015年10期)2015-12-01 06:51:18
