
利用GS3基因功能性分子标记改良水稻粒型的研究
2016-04-07 08:40徐小艳冯芳君马孝松梅捍卫
上海农业学报 2016年1期
李 扬,徐小艳,严 明,冯芳君,马孝松,梅捍卫*
(1上海海洋大学水产与生命学院,上海200090;2上海市农业生物基因中心,上海201106)
利用GS3基因功能性分子标记改良水稻粒型的研究
李 扬1,2,徐小艳2,严 明2,冯芳君2,马孝松2,梅捍卫1,2*
(1上海海洋大学水产与生命学院,上海200090;2上海市农业生物基因中心,上海201106)
摘 要:以长粒品种‘三粒寸’‘IAC1246’‘JAPPENI TUNGKUNGO’和‘MIGA’作为供体亲本,以优良选系SAGC-4作为受体亲本,进行杂交和连续回交,利用GS3基因的功能性分子标记SF28进行BC1F1和BC2F1的分子标记辅助选择。在不同BC2F1单株派生的BC2F2分离群体中,观察到粒长性状的典型双峰分布或单峰连续分布。结合田间农艺性状考察,选出89个具备细长粒型且综合农艺性状良好的单株,其中4个优良单株的平均粒长从原始受体亲本的8" 32 mm增加至9" 84 mm左右,平均长宽比由原来的2" 61增加至3" 00。结果表明:GS3基因具有控制粒长和长宽比的较大遗传效应,对该基因进行分子标记辅助选择可快速改良亲本的粒型性状。
关键词:水稻;粒型;GS3;分子标记辅助选择
水稻是最重要的粮食作物之一,优质稻的发展已成为消费者和水稻育种工作者共同关注的焦点问题之一。提高产量、改良稻米品质是水稻育种的两个主要目标,水稻粒型既是重要的稻米外观品质性状,也与粒重等产量构成因素密切相关。水稻粒型主要通过粒长、粒宽、长宽比和粒厚等4个指标进行描述,由于世界上大多数地区的消费者更偏爱细长型的稻米[1],因此提高粒长和长宽比是籼稻品种的重要育种目标。
水稻中控制粒长的主效基因GS3已被克隆出来[2],该基因是粒长和粒重的主效QTL,对粒宽和粒厚只有微效作用。该基因编码的有功能蛋白表现为粒长的反向调控因子,在第二个外显子第165个核苷酸上的C-A单碱基替换,造成编码蛋白质提前终止,具有增加水稻粒长的功能,由此开发出GS3基因的CAPS功能性分子标记SF28[3-4]。然而,GS3基因的调控模式决定了细长型谷粒为隐性性状,该基因的杂合基因型和显性纯合基因型都表现出相同的短粒表型,在连续回交后代中粒长性状没有分离,不能根据表型直接进行选择,如果逐代自交分离后选择单株进行回交,粒型性状改良的时间将延长近一倍。利用分子标记辅助选择技术,在水稻幼苗阶段进行紧密连锁或者基因内共显性标记的基因型检测,能准确判断目标基因的杂合及纯合基因型,选择杂合单株在抽穗后与受体亲本回交,保证后代中有1/2单株携带GS3的长粒型等位基因,通过连续回交达到快速改良受体亲本粒长性状的目的。
SAGC-4是上海市农业生物基因中心通过多亲本导入系相互杂交,后代经干旱胁迫和不施氮肥大田等条件的鉴定筛选,培育的常规稻优良选系之一。在比尔及梅琳达盖茨基金会“绿色超级稻”国际合作项目中,该选系在多个亚洲、非洲国家的试种观察和多点试验中,尤其在雨养田试验中表现突出。但由于该选系的籽粒不够细长,在菲律宾等东南亚国家不符合居民的消费习惯,影响了其在这些国家的审定和推广。
本研究以长粒品种‘三粒寸’‘IAC1246’‘JAPPENI TUNGKUNGO’和‘MIGA’等作为供体亲本,采用分子标记辅助回交育种方法,对受体亲本SAGC-4的粒型进行快速的遗传改良,以培育在我国、亚洲及非洲国家具有应用前景的常规水稻新品种。
1 材料与方法
1.1 试验材料与分子标记辅助选择流程
SAGC-4属中等粒型,用作受体亲本;长粒品种‘三粒寸’(中国)、‘IAC1246’(巴西)、‘JAPPENI TUNGKUNGO’(塞内加尔)和‘MIGA’(巴西)等作为供体亲本。
受体亲本SAGC-4分别与上述4个供体亲本杂交,标记检测选择GS3/gs3单株与SAGC-4回交,每个回交群体均进行分子标记检测,选择GS3/gs3杂合基因型,同时农艺性状与SAGC-4较为相似的单株2次回交后进行1次自交,获得BC2F2分离群体。
1.2 粒型考察和单株选种
2014年冬到2015年春在上海市农业科学院海南陵水南繁基地大田种植亲本和BC2F2群体,成熟后从每个单株上收获一个穗子上的种子,利用图像分析系统(SC-G自动考种分析与千粒重仪,杭州万深检测科技有限公司)测量籽粒的平均长度和宽度,计算长宽比。
田间结合粒型、株高、分蘖力、分蘖角度、株叶型、穗型、结实率等性状的目测评判,选择农艺性状优良、粒型细长的改良单株,成熟后考察株高、单株有效穗数、每穗颖花数、每穗实粒数、结实率和千粒重等主要农艺性状,其中4个改良单株在株叶型、丰产性等方面表现突出,本研究以此为例分析粒型改良效果。
1.3 分子标记分析
采集水稻幼嫩叶片并利用CTAB法[5]快速提取DNA,利用Fan等[4]开发的SF28标记在亲本间进行多态性分析,在后代群体间进行基因型检测。
SF28标记上下游引物序列分别为:SF28-F(5’-TGCCCATCTCCCTCGTTTAC-3’);SF28-R(5’-GAAACAGCAGGCTGGCTTAC-3’)。
PCR反应体系为:DNA模板(50 ng/μL)2" 0 μL,10×缓冲液(含Mg2+)2" 0 μL,dNTPs(10 mmol/L)2" 0 μL,引物各0" 5 μL,Taq DNA聚合酶(5 U/μL)0" 5 μL,灭菌双蒸水(ddH2O)12" 5 μL。反应条件为:94℃预变性4 min;94℃变性40 s,55℃退火40 s,72℃延伸40 s,34个循环;72℃延伸10 min。
PCR反应结束后进行PstⅠ酶切反应。PstⅠ酶切体系为:PCR产物8" 0 μL,PstⅠ酶(15 U/μL)0" 1 μL,10×H缓冲液2" 0 μL,灭菌双蒸水(ddH2O)9" 9 μL。37℃水浴2 h,取8" 0 μL的酶切反应终产物在2" 0%琼脂糖凝胶上电泳检测,通过凝胶成像仪观察和照相。
2 结果与分析
2.1 亲本间的分子标记多态性及后代群体中标记检测效果
利用GS3基因功能性CAPS分子标记SF28对供体亲本、受体亲本进行多态性分析,结果表明:SF28在供体与受体亲本间存在明显的多态性,在电泳图谱上酶切条带具有分子量上的明显差异,即受体亲本的带型(C)和供体亲本的带型(A)(图1)。在连续回交后代中,SF28的扩增和酶切产物呈现两种带型,即受体亲本SAGC-4的带型(C)和杂合带型(H)(图2)。

图1 SF28扩增与酶切产物在受体与供体亲本间的多态性Fig.1 Polymorphism of SF28 amplification and digestion products between the recipient and donor parents

图2 部分BC2F1单株的SF28标记检测结果Fig.2 DNA bands detected by SF28 marker of some BC2F1plants
2.2 GS3基因的功能分子标记辅助选择
将‘三粒寸’‘IAC1246’‘JAPPENI TUNGKUNGO’和‘MIGA’与SAGC-4分别杂交,利用GS3基因功能性CAPS分子标记SF28鉴定F1的真伪,GS3杂合型真杂种与SAGC-4回交。在秧苗期进行BC1F1群体每个单株的SF28标记检测,选择具有GS3杂合基因型且田间农艺性状与SAGC-4较相似的单株,在抽穗期与SAGC-4回交。在4个回交组合的BC2F1代群体中筛选GS3基因杂合型且田间农艺性状与SAGC-4较相似的单株,收获自交种子。在BC2F2代群体中观察粒长性状的分离情况,在长粒型单株中根据田间目测的农艺性状表现,选择出具备细长粒型且综合农艺性状优良的单株89株。
2.3 BC2F2群体的粒长分离情况
选择4个不同回交组合在BC2F2代的分离群体进行粒长分析(图3)。来自‘三粒寸’‘IAC1246’与SAGC-4回交后代的14HBL-16和14HBL-27等2个分离小区中,单株平均粒长的频率分布呈现典型的双峰分布。以图3中双峰交叉处为分界,左侧的短粒单株数与右侧的长粒单株数之间的比例分别为2" 88∶1和4" 13∶1,经卡方测验符合一对基因3∶1的理论分离比,表明在回交群体背景下GS3能够表现出单个主效基因的效应。

图3 不同回交组合分离群体的粒长频率分布Fig.3 The grain length frequency distribution of different backcross segregation populations
来自‘JAPPENI TUNGKUNGO’‘MIGA’与SAGC-4的14HBL-42和14HBL-66等2个BC2F2群体中,单株平均粒长的分离情况与前者不同,粒长的频率分布图虽然在长粒侧有较长的尾部,但总体上呈现连续分布(图3)。造成这一现象的原因可能是群体中除GS3位点之外还同时存在其他控制粒型基因的分离,使得双峰分布不明显。
2.4 入选单株的粒型改良效果
从4个不同回交组合中各选择1个优良单株,分别命名为GL1/2、GL1/3、GL1/4和GL1/5(图4)。从表1可以看出,4个供体亲本与轮回亲本比SAGC-4粒长分别长1" 5—2" 2 mm,4个改良单株的粒长变化范围为9" 67—9" 98 mm,均比SAGC-4的粒长极显著增加,平均为9" 84 mm,比改良前的受体亲本SAGC-4提高1" 52 mm。
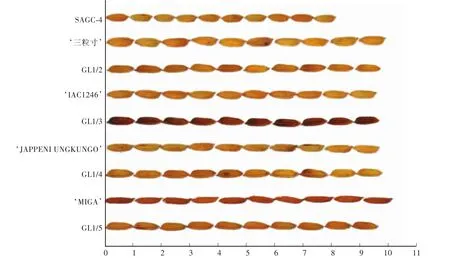
图4 亲本和改良单株的水稻粒型对比Fig.4 Comparison of rice grain shape between the parents and improved individuals

表1 受体亲本、供体亲本与入选单株的粒型比较Table 1 Comparison of rice grain shape among the parents and selected individuals
除GL1/4的粒宽有所增加以外,另3个入选单株的粒宽与受体亲本相近。4个入选单株籽粒长宽比的变化范围为2" 99—3" 04,平均为3" 00,比原始受体亲本的2" 61增加了0" 39。结果表明:利用本试验的4个长粒型供体亲本和针对GS3的功能性分子标记辅助选择方法,都可以对受体亲本SAGC-4的粒型进行有效的改良,保持粒宽基本不变,显著提高粒长和长宽比,培育SAGC-4的细长粒型改良系。
3 结论与讨论
GS3是一个控制粒长和粒重的主效QTL,其长粒等位基因广泛存在于诸多栽培品种中。通过分子育种手段对该基因加以充分利用,不仅可以改善待改良品种的外观品质,而且可以增加粒重、提高产量,具有十分广阔的应用前景。
目前在改良水稻粒型的分子育种中,重要粒型基因的利用还不是非常广泛,多数报道都选择了GS3作为目标基因。杨梯丰等[6]和Wang等[7]分别利用单片段代换系和构建近等基因系进行聚合育种,研究结果均证实GS3对增加粒长和粒重起到非常关键的作用。方珊茹等[8]利用连锁标记MRG5881和GS09进行回交导入法结合MAS和农艺性状选择,将GS3长粒等位基因及其他外观品质性状基因导入‘Ⅱ-32B’中,获得粒长、长宽比和粒重均得到改良的株系。张剑霞[9]利用SF28标记的MAS技术,将GS3的长粒等位基因和Xa23聚合到‘珍汕97B’和‘Ⅱ-32B’中,获得各自粒长得到改良的株系。本研究利用Fan等[4]开发的GS3基因功能性分子标记SF28进行回交导入结合农艺性状选择,从BC2F2代分离群体中直接筛选出长粒表型并且综合农艺性状表现优良的入选单株。从优良入选单株的粒型对比结果来看,不同供体亲本对受体亲本SAGC-4粒长增加的贡献基本相同,都可以达到快速改良SAGC-4粒长和长宽比性状的目的。与此同时,由于GS3基因不是粒宽和粒厚的主效基因,导入该基因不会改变粒宽性状,改良单株的籽粒宽度几乎与受体亲本相近,从籽粒外观品质的更高要求来看,可能需要通过后续改良适当降低粒宽,选育具有更为细长籽粒类型的品种。SAGC-4本身存在株型过于紧束的缺点,在本试验的BC2F2群体中可以见到分蘖角度和株叶型的广泛分离,通过对株叶型的目测选择,大部分入选单株的株叶型比原始受体亲本更为适中,有利于形成合理的群体冠层结构。
目前,优选的单株将在上海进行小区种植,在正季气候环境、高产田间管理条件下进行农艺性状、产量构成等鉴定,结合籽粒外观和食味品质等进行下一轮选择,以获得比原始受体亲本SAGC-4粒长、长宽比显著增加,农艺性状和丰产性相当或者更好的选系,稳定后的高代选系将提交后续品比试验、病虫害抗性等鉴定筛选以及国内外的测试和示范试种。
参 考 文 献
[1]Juliano B O,Villareal C" Grain quality evaluation of world rices[M]" Los Banos:IRRI,1993"
[2]Takano-Kai N,Jiang H,Kubo T,et al" Evolutionary history of GS3,a gene conferring grain length in rice[J]" Genetics,2009,182:1323-1334"
[3]Fan C,Xing Y,Mao H,et al" GS3,a major QTL for grain length and weight and minor QTL for grain width and thickness in rice,encodes a putative transmembrane protein[J]" Theoretical Applied Genetics,2006,112:1164-1171"
[4]Fan C,Yu S,Wang C,et al" A causal C-A mutation in the second exon of GS3 highly associated with rice grain length and validated as a functional marker[J]" Theoretical Applied Genetics,2009,118:465-472"
[5]Murray M G,Thompson W F" Rapid isolation of high molecular weight plant DNA[J]" Nucleic Acids Research,1980,8:4321-4325"
[6]杨梯丰,曾瑞珍,朱海涛,等"水稻粒长基因GS3在聚合育种中的效应[J]"分子植物育种,2010,8(1):59-66"
[7]Wang P,Xing Y,Li Z,et al" Improving rice yield and quality by QTL pyramiding[J]" Molecular Breeding,2011,29(4):903-913"
[8]方珊茹,吴春珠,刘玉芹,等"分子标记辅助选择改良Ⅱ_32B的外观品质[J]"分子植物育种,2013,11(6):673-679"
[9]张剑霞"利用分子标记辅助选择转移野生稻增产QTL和聚合水稻优良基因[D]"武汉:华中农业大学,2009"
(责任编辑:闫其涛)
Improvement of rice grain shape by functional molecular marker of GS3 gene
LI Yang1,2,XU Xiao-yan2,YAN Ming2,FENG Fang-jun2,MA Xiao-song2,MEI Han-wei1,2*
(1College of Fisheries and Life Science,Shanghai Ocean University,Shanghai 200090,China;
2Shanghai Agrobiological Gene Center,Shanghai 201106,China)
Abstract:Four donor parental lines with long grain shape,‘SANLICUN’,‘IAC1246’,‘JAPPENI TUNGKUNGO’and‘MIGA’,were crossed with a promising indica rice line SAGC-4 as the recipient parent,followed by continuous backcrosses" Marker-assisted selection was conducted in both BC1F1and BC2F1generations by using the CAPs marker SF28 of GS3 gene" Segregation of grain length showed typical bimodal or continuous distributions in BC2F2populations" A set of 89 individuals were selected based on both grain shape and visual inspection on other agronomic traits" Among them,4 promising individuals had average grain length of 9" 84 mm and length/width ratio of 3" 00,in comparison to 8" 32 mm and 2" 61 of the original recipient parent,respectively" The results indicated that GS3 gene had large genetic effects on both grain length and length/width ratio" Markerassisted selection of the gene could accelerate the improvement of rice grain shape"
Key words:Rice;Grain shape;GS3;Marker-assisted selection(MAS)
*通信作者,E-mail:hmei@sagc" org" cn
作者简介:李扬(1989—),男,在读硕士,主要从事节水抗旱稻分子育种研究。E-mail:foryli@163" com
基金项目:比尔及梅琳达盖茨基金“绿色超级稻”国际合作项目;上海市科委基础研究重点项目(12JC1408000)
收稿日期:2015-05-26
文章编号:1000-3924(2016)01-001-05
中图分类号:S511" 035
文献标识码:A
猜你喜欢
青少年科技博览(中学版)(2022年6期)2022-12-27
农学学报(2021年6期)2021-12-26
今日农业(2021年21期)2021-11-26
军事文摘(2021年22期)2021-11-26
今日农业(2021年14期)2021-10-14
江苏农业科学(2021年12期)2021-08-02
文苑(2020年6期)2020-06-22
江苏农业学报(2020年2期)2020-05-27
文苑(2019年22期)2019-12-07
山东农业科学(2018年11期)2018-01-17
