
水分胁迫和斜发沸石应用对花生叶片光合特性及水分利用的影响
2016-03-23 03:53姚珍珠夏桂敏王淑君胡家齐李永发迟道才沈阳农业大学水利学院沈阳110161
中国农村水利水电 2016年9期
姚珍珠,夏桂敏,王淑君,胡家齐,李永发,迟道才(沈阳农业大学 水利学院,沈阳 110161)
花生(Arachis hypogaea Linn) 是我国主要的油料作物和重要的经济作物,其生产发展对于增加我国蛋白质及食用植物油的供给具有重要的作用[1-5]。干旱是限制和危害花生生产种植的最大因素之一[6]。据统计,常年干旱造成我国70%的花生受到不同程度干旱胁迫的影响,花生减产占全国总产量的20%以上,且使花生品质降低,黄曲霉毒素污染严重[7,8]。光合作用是影响作物生产最重要的过程[9],其所包含的光合速率、蒸腾速率和气孔导度等指标日变化,直接反应植物叶片对水分胁迫的响应机制[10]。叶片水分利用效率反应植物生产过程中单位水分的转化效率,是植物光合速率和蒸腾速率共同作用的结果[11]。不同土壤水分胁迫下,作物光合特性和水分利用效率不同,水分胁迫会导致植物光合作用下降,作物增产失去原动力最终导致产量大幅降低。很多研究结果表明,提高水分胁迫阶段土壤含水量能够增加叶片光合特性参数,从而提高作物产量[12]。
斜发沸石作为一种良好的非金属节水材料,将其运用在农业生产中除了能够提高土壤物理特性外,还能够暂时缓解土壤水分胁迫,降低生长时期植物萎蔫的风险[13]。已有研究结果表明,沸石是一种具有三维晶体结构的水合硅铝酸盐,能够非破坏性改变其构成元素来可逆性吸收和释放水分。魏江生对人工沸石在干旱地区农业开发中的作用研究时,发现土壤中加入沸石,可使土壤有效水含量和有效雨量增大,从而减少灌溉水量[14]。曹晓燕总结天然沸石的应用得出,砂质土壤施入沸石可增加其持水能力,减缓土壤水分的无效损失,利于作物对水分吸收[15]。Reza对大豆进行水分胁迫条件下沸石和牛粪施用的试验得到,沸石和牛粪的施用提高土壤相对含水量,缓解叶片细胞相应酶活性,进而改善大豆水分胁迫条件下生长环境[13]。但大多数针对斜发沸石运用在农业生产中的研究,并没有涉及水分胁迫下斜发沸石如何影响作物光合特性,进而调节作物自身抗旱机制实现节水增产的目的影响规律。如何科学地将其结合土壤水分胁迫,施用在花生生产实践并发挥其节水保肥的功效,缓解水分胁迫对花生造成的减产损失,对我国花生节水生产具有重要的现实意义。为此,本文以花生为研究对象,进行了盆栽试验,以便进一步深入探讨水分胁迫和斜发沸石施用花生需水关键期净光合速率、蒸腾速率、气孔导度及叶片水分利用效率日变化规律,有助于从理论上分析斜发沸石对水分胁迫下花生光合作用及水分利用的影响规律,以期为斜发沸石运用在花生的生产实践提供理论依据和技术支撑。
1 材料与方法
试验于2015年5月-2015年10月在辽宁省农科院作物抗旱栽培模拟试验场内进行,试验场设有自动遮雨棚。试验采用盆栽覆膜种植方式,盆高30 cm,上、下内径分别为26和18 cm。试验采用耕层0~30 cm的表层土,土壤类型为褐土,土壤质地为砂壤土,0~30 cm土壤田间持水量为32%,土壤容重为1.4 g/cm3。试验前将土壤进行风干,2 mm过筛后装盆,每盆装风干土20 kg,其风干土含水量2.66%。种植前,对盆进行灌水,使土壤含水量达到田间持水量的70%。将36 g(折合大田为1.5 t/hm2)斜发沸石同表土进行混合后,将9.54 g N-P-K复合肥(折合大田600 kg/hm2)穴施于盆内,距离种穴水平7 cm。选取饱满一致的种子3粒穴播,每盆一穴。出苗后间苗,每盆剩余1株。
1.1 试验处理
试验采用裂区试验设计,共设两个变量因子:土壤水分胁迫W和斜发沸石Z,花生需水关键期土壤水分胁迫W设为主区,设3个水平:土壤相对含水量为田间持水量的 55%±5%(重度胁迫W1)、65%±5%(中度胁迫W2)和75%±5%(轻度胁迫W3),其他生育期均保持田间持水量75%±5%;斜发沸石(Z)设为副区,设2个水平:Z1(无斜发沸石)和Z2(斜发沸石施用量36 g/盆)。试验期间通过确定土壤含水率达田间持水率时盆的总重,用称重法控制土壤水分上下限,灌水量根据盆内土壤含水率分别达到相应田间持水率的上下限百分数时的总重量之差确定。试验共设6个处理,每个处理3次重复,具体试验处理布置如表1。使用美国LI-COR公司生产的LI-6400便携式光合仪,选择在晴朗无云的天气测定不同处理花生顶部完全展开叶的光合速率Pn、蒸腾速率Tr、气孔导度gs的日变化,每2h测定一次,各次测定在30 min内完成。同时计算单叶水分利用效率,单叶水分利用效率(WUE)用叶片通过蒸腾消耗一定量的水(mmol)所同化的CO2量(μmol)来表示,即WUE=Pn/Tr。

表1 试验处理布置表格Tab.1 Treatments arranged table
1.2 数据分析
所有数据为3次重复的平均值,用SAS 9.4进行数据的统计分析,并用sigmaplot软件进行作图。
2 结果与分析
2.1 重度水分胁迫下斜发沸石应用花生叶片光合、蒸腾、气孔导度及水分利用效率日动态变化
图1为重度水分胁迫花生叶片光合速率、蒸腾速率、气孔导度及水分利用效率的日动态。由图1可知,Z1和Z2的净光合速率日变化有明显的差异。Z1呈现早晚和中午低的双峰曲线,上午10∶00左右(一天中最高值)出现第一个峰值,其值为15.36 μmol/(m2·s),下午14∶00左右出现第二个峰值,其值为13.32 μmol/(m2·s),中午12∶00左右由于气孔关闭导致光合作用减弱,净光合速率下降了4.34 μmol/(m2·s)。Z2叶片净光合速率日变化呈现不明显的单峰曲线,早晚时段净光合速率较低,上午10∶00达到最大值16.39 μmol/(m2·s),中午12∶00净光合速率降低,降低幅度为2.12 μmol/(m2·s)。Z2比Z1净光合速率提高幅度最大值为3.24 μmol/(m2·s),出现在午间12∶00左右;Z1在下午14∶00之前叶片净光合速率均低于Z2,于下午14∶00之后逐渐高于Z2。重度水分胁迫斜发沸石应用提高花生上午和中午时段叶片净光合速率,减少午间净光合速率降低幅度,极大缓解重度水分胁迫下花生为适应水分胁迫刺激自身调整系统对其叶片净光合速率的影响。
Z1和Z2的蒸腾速率日变化有明显的差异。Z1呈现早晚和中午低经典双峰曲线,两个峰值分别出现在上午10∶00左右(一天中最高值)和下午14∶00左右,其值分别为7.65和6.98 mmol/(m2·s),中午12∶00左右因气孔关闭导致蒸腾速率瞬间下降了1.73 mmol/(m2·s);Z2呈现不明显的单峰曲线,最大值出现在上午10:00左右,其值为7.74 mmol/(m2·s),中午12∶00左右很可能由于非气孔因素和气孔因素导致蒸腾速率降低了0.68 mmol/(m2·s),之后继续下降。Z2比Z1蒸腾速率提高幅度最大值为1.14 mmol/(m2·s),出现在午间12∶00左右;Z2在最大值10∶00至13∶30左右蒸腾速率高于Z1,下午14:00左右后渐渐低于Z1。重度水分胁迫斜发沸石减小花生午间蒸腾速率下降幅度,降低“午休”对花生蒸腾速率的影响,同时降低花生叶片奢侈蒸腾,从而缓解花生水分亏缺状况。

图1 重度水分胁迫叶片光合速率、蒸腾速率、气孔导度和水分利用效率日动态Fig.1 Daily variations of photosynthesis rate,transpiration rate,stomatal conductance and water use efficiency of peanut leaves in severe water deficit stress
Z1和Z2气孔导度日变化有明显的差异。Z1呈现明显的双峰曲线,上午10∶00左右(一天中最高值)和下午14:00左右达到气孔导度达到峰值,其值分别为0.25和0.20 mol/(m2·s),12∶00左右气孔导度明显降低了0.09 mol/(m2·s);Z2呈现不规则的单峰曲线,10∶00左右达到峰值,其值为0.24 mol/(m2·s),随后一直下降,但12:00左右高于Z1,并在14∶00左右微小回升后继续下降。Z1除11∶00-13∶00左右气孔导度值低于Z2之外,剩余时段气孔导度值均高于Z2。重度水分胁迫斜发沸石降低花生叶片午间气孔导度下降幅度,减小水分胁迫下气孔开度,缓解午间气孔导度骤然降低对花生水分利用效率的影响。
Z1和Z2的花生叶片水分利用效率WUE日变化差异显著。Z1和Z2的叶片水分利用效率WUE日变化均呈现明显的双峰曲线,Z1和Z2均在上午10:00左右,达到第一个峰值(一天中最高值),其值分别为2.01和2.12 μmol /mol,Z1在下午16∶00左右达到第二个峰值,其值为1.92 μmol/mol,Z2较Z1提前一个小时左右达到第二个峰值即下午14∶00左右,原因可能由于Z2降低气孔开度导致峰值提前。Z2比Z1叶片水分利用效率提高幅度最大值0.16 μmol/mol出现12∶00左右。重度水分胁迫斜发沸石提高了叶片水分利用效率,同时“午休”后的峰值出现提前。
2.2 中度水分胁迫下斜发沸石应用花生叶片光合、蒸腾、气孔导度及水分利用效率日动态变化
图2为中度水分胁迫花生叶片光合速率、蒸腾速率、气孔导度及水分利用效率的日动态。由图2可知,Z1和Z2的净光合速率日变化有差异。Z1和Z2净光合速率日变化均呈现早晚低中午高明显的单峰曲线,且Z2高于Z1。Z1和Z2上午10∶00左右均出现峰值,分别为19.83和20.54 μmol/(m2·s),随后降低。Z2比Z1净光合速率日变化提高了0.11~0.85 μmol/(m2·s),且提升幅度最大值出现在早上6∶00左右和下午14∶00左右。中度水分胁迫斜发沸石提高花生叶片的净光合速率。

图2 中度水分胁迫叶片光合速率、蒸腾速率、气孔导度和水分利用效率日动态Fig.2 Daily variations of photosynthesis rate,transpiration rate,stomatal conductance and water use efficiency of peanut leaves in moderate water deficit stress
Z1和Z2的蒸腾速率和气孔导度日变化规律相似,即Z1和Z2日变化均呈现明显的双峰曲线,且Z1高于Z2。Z1和Z2均在上午10∶00左右出现第一个峰值,且两者均在第一个峰值处达到一天中最大值8.73 mmol/(m2·s)、0.27 mol/(m2·s)和8.56 mmol/(m2·s)、0.24 mol/(m2·s);Z1和Z2均在上午12∶00左右出现突降;中午14∶00左右出现第二个峰值,7.62 mmol/(m2·s)、0.25 mol/(m2·s)和7.50 mmol/(m2·s)、0.22 mol/(m2·s)。Z2比Z1的蒸腾速率降低幅度最大值0.66 mmol/(m2·s)出现在8∶00左右。中度水分胁迫斜发沸石降低花生蒸腾速率和气孔导度。
Z1和Z2叶片水分利用效率(WUE)日变化有差异。Z1和Z2叶片水分利用效率均呈现早晚低中间高的单峰曲线。Z1和Z2均在12:00左右达到峰值,且达到一天中最大值2.41和2.59 μmol/mol。Z2比Z1叶片水分利用效率提高了0.13~0.73 μmol/mol,提高幅度最大值出现在18∶00左右。中度水分胁迫斜发沸石明显提高花生叶片午间水分利用效率,可能由于叶片为适应水分胁迫控制午间气孔开度,提高叶片对水分的转化能力。
2.3 轻度水分胁迫下斜发沸石应用花生叶片光合、蒸腾、气孔导度及水分利用效率日动态变化
图3为轻度水分胁迫花生叶片光合速率、蒸腾速率、气孔导度及水分利用效率的日动态。由图3可知,Z1和Z2的净光合速率日变化有差异。Z1和Z2均单峰曲线,且Z2高于Z1。Z1和Z2净光合速率日变化上午10∶00左右出现峰值,其值分别为17.78和20.12 μmol/(m2·s),随后降低。Z2比Z1净光合速率日变化提高了0.39~2.94 μmol/(m2·s)。轻度水分胁迫斜发沸石提高了花生叶片的净光合速率。
Z1和Z2的蒸腾速率、气孔导度日变化趋势相同,Z1和Z2均呈现早晚中间低的双峰曲线,且Z2高于Z1。Z1和Z2均在上午10:00左右出现第一个峰值,其值分别为8.17 mmol/(m2·s)、0.28 mol/(m2·s)和8.92 mmol/(m2·s)、0.31 mol/(m2·s),随后因气孔关闭于12∶00左右降低,14∶00左右出现第二个峰值,其值分别为7.75 mmol/(m2·s)、0.27 mol/(m2·s)和8.26 mmol/(m2·s)、0.28 mol/(m2·s)。轻度水分胁迫斜发沸石提高了花生叶片蒸腾速率和气孔导度。
Z1和Z2叶片水分利用效率(WUE)日变化有差异。Z1和Z2均呈现明显的双峰曲线。Z1和Z2均在10∶00左右达到第一个峰值,其值分别为2.18 和2.26 μmol/mol,16∶00左右达到第二个峰值,其值分别为1.79和2.14 μmol/mol;14∶00左右,两者的叶片水分利用效率均降低。Z2比Z1叶片水分利用效率提高幅度最大值为0.35 μmol/mol。轻度土壤水分胁迫斜发沸石提高了花生叶片水分利用效率,且下午提升幅度高于上午。
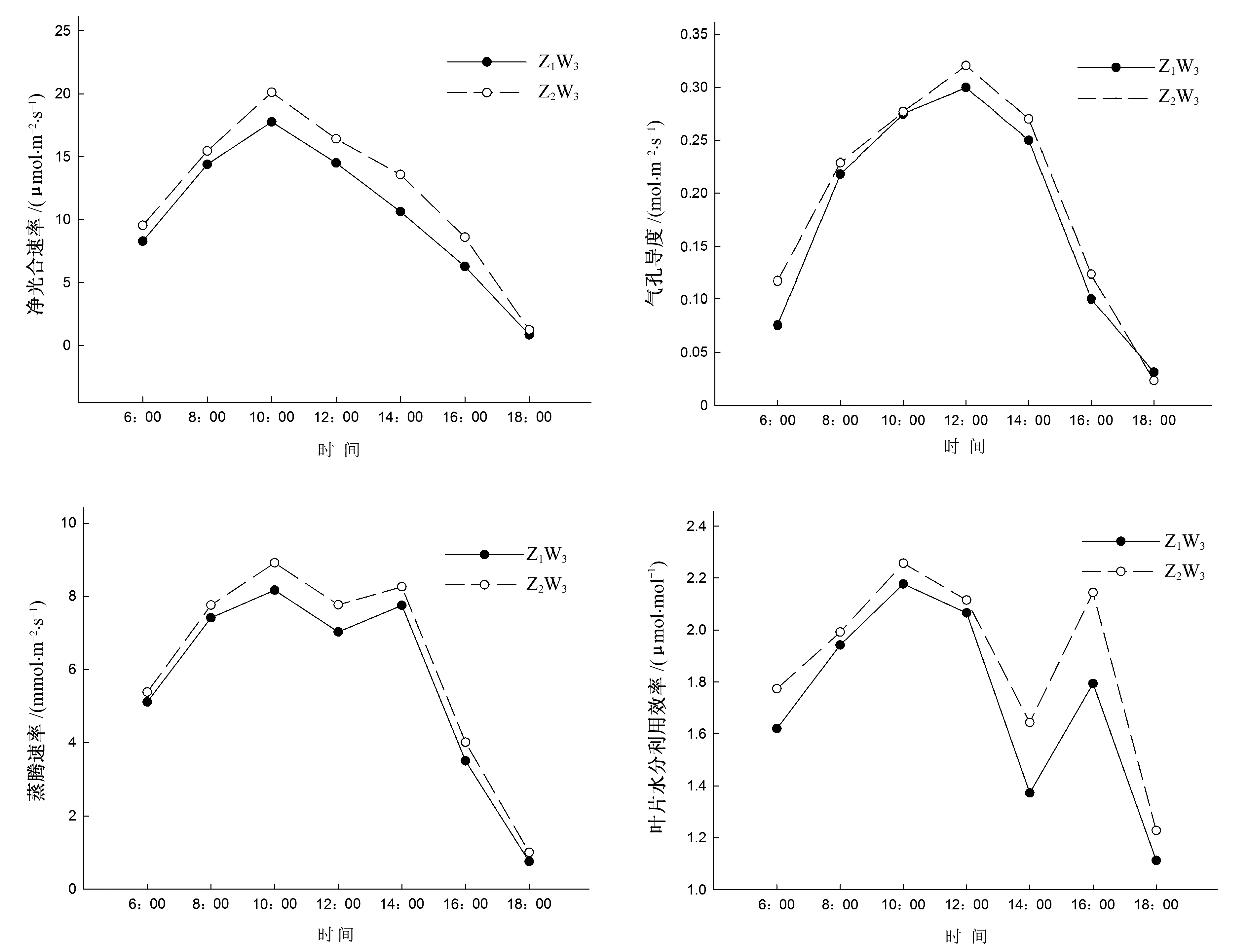
图3 轻度水分胁迫叶片光合速率、蒸腾速率、气孔导度和水分利用效率日动态Fig.3 Daily variations of photosynthesis rate,transpiration rate,stomatal conductance and water use efficiency of peanut leaves in mild water deficit stress
2.4 不同水分胁迫下斜发沸石应用对花生叶片光合、蒸腾、气孔导度及水分利用效率日动态变化
图4为施用斜发沸石不同水分胁迫下花生叶片净光合速率、蒸腾速率、气孔导度及水分利用效率的日动态。由图4可知,不同水分胁迫下花生叶片净光合速率日变化均呈现单峰曲线。10∶00-16∶00左右,W2比W3净光合速率提高了0.31~1.99 μmol/(m2·s),且于14∶00左右提高幅度最大;18∶00左右,W1处理净光合速率最大,其值为1.58 μmol/(m2·s),相比W2和W3增加0.36和0.35 μmol/(m2·s),说明午间中度水分胁迫调节斜发沸石缓释水分能力最佳,缓解水分胁迫,弥补水分胁迫对叶片净光合速率的降低,晚间重度水分胁迫最佳。不同水分胁迫下花生叶片蒸腾速率和气孔导度日变化差异相似, W2和W3呈现双峰曲线,W1呈现单峰曲线。14:00之前,叶片蒸腾速率W3>W2>W1,之后W2>W3;午间12∶00左右,W2比W1蒸腾速率提高了0.04 mol/(m2·s),两者相差甚微;8∶00之前叶片气孔导度W1>W2>W3;8∶00-12∶00时段叶片气孔导度W1>W2;12∶00之后W3>W2>W1,说明水分胁迫不仅控制气孔导度调节叶片抗旱机制,还激发细胞内非气孔因素应对水分胁迫,同时斜发沸石提高土壤含水量,调节花生根系水分胁迫程度进而控制叶片气孔开度,尤其重度水分胁迫效果最明显。不同水分胁迫下花生叶片水分利用效率日变化差异显著。W2和W3一天中变化剧烈,W1变化较平稳;一天中叶片水分利用效率W2>W3,且提高幅度最大值0.48 μmol/mol出现在午间12∶00左右;早间和晚间叶片水分利用效率W3最小,可能由于奢侈蒸腾较多引起。
3 结论与讨论
3.1 不同水分胁迫条件下,斜发沸石对花生叶片光合特性的影响不同
重度水分胁迫条件下,花生根系对土层含水量需求得不到满足,导致叶片净光合速率Pn、气孔导度gs和蒸腾速率Tr对水分亏缺变得很敏感,尤其在午间叶片气孔关闭,3个参数均降低,但土壤中施加斜发沸石后,斜发沸石因午间土壤含水量骤降刺激其体内自由形态水分子释放,提高相同时段土壤含水量,减缓花生叶片净光合速率、气孔导度和蒸腾速率降低幅度,从而缓解花生“午休”对其造成的水分胁迫损伤;中度水分胁迫条件下,土壤含水量相对重度水分胁迫较高,基于斜发沸石具有比表面积大,吸附量大及选择吸收性离子交换性的特点,如同“海绵”般蓄存一定的水量及营养离子,配合水分胁迫调节叶片气孔开度,降低花生叶片的蒸腾作用,减少叶片奢侈蒸腾;轻度水分胁迫下,斜发沸石的应用提高花生生长环境,促进花生生长和生理代代谢,直接表现出叶片光合特性三大指标均增加。很多研究者研究得到斜发沸石的施用能够暂时缓解土壤水分胁迫,Xiubin研究斜发沸石应用对黄土渗透及截留影响得到,斜发沸石能够提高耕土层保水能力和阳离子交换能力[16];He研究砂质土壤中斜发沸石对氨挥发影响得出,斜发沸石能够通过增加土壤表面积和阳离子交换能力,提高土壤含水量和土壤肥力,减少氮的渗漏损失[17];Zahedi研究水分胁迫下施用斜发沸石和硒对菜籽抗氧化酶活性的影响,发现土壤施入斜发沸石能够提高土壤水分蓄存能力,减缓土壤水分胁迫,进而降低抗氧化酶活性[18]。研究表明,水分胁迫条件下,斜发沸石施用通过缓解土壤水分胁迫对花生的损伤,以激发花生叶片抗旱机制,调节光合特性和蒸腾作用使得叶片适应水分胁迫,以此提高叶片抗旱能力。

图4 施用斜发沸石不同水分胁迫下花生叶片光合速率、蒸腾速率、气孔导度和水分利用效率日动态Fig.4 Daily variations of photosynthesis rate,transpiration rate,stomatal conductance and water use efficiency of peanut leaves with clinoptilolite application in different water deficit stress
3.2 应用斜发沸石不同水分胁迫对花生叶片光合特性及水分利用的影响
相比重度和轻度水分胁迫,中度水分胁迫下,花生叶片午间水分利用效率最大,可能由于中度水分胁迫能够调整花生叶片抗旱机制处于最佳状态,叶片蒸腾速率(Tr)较低,净光合速率(Pn)因滞后效应受水分胁迫影响较小,由此叶片对水分的利用生产效率最大;重度水分胁迫,花生叶片水分利用效率日变化最稳定,原因可能由于水分胁迫不仅控制气孔导度调节叶片抗旱机制,还激发细胞内非气孔因素应对水分胁迫,同时斜发沸石提高土壤含水量,调节花生根系水分胁迫程度,进而控制叶片净光合速率和蒸腾速率呈现相同的变化规律,最终展现叶片水分利用效率日变化稳定;轻度水分胁迫,叶片生理代谢进程较为旺盛,光合作用和蒸腾作用均处于较高水平,叶片水分利用效率却不是最佳水平,说明叶片对水分利用效率是植物细胞光合作用和蒸腾作用综合调整的过程,并不是单一因素决定其大小。综上所述,中度水分胁迫花生叶片水分利用效果最佳,这同于文颖等研究水分胁迫下玉米叶片光合特性及水分利用效率所得结论一致,中度水分胁迫提高玉米叶片水分利用效率,增强叶片对水分利用能力和抗旱能力[19]。
□
[1] 苗锦山,王铭伦.水分胁迫对花生生长发育影响的研究进展[J].花生学报,2003,32(增刊):368-371.
[2] 张 俊,刘 娟,臧秀旺,等.不同生育时期干旱胁迫对花生产量及代谢调节的影响[J]. 核农学报,2015,29(6):1 190-1 197.
[3] Bhavanath Jha,Avinash Mishra,Amit Kumar Chaturvedi. Enginner Stress Tolerance in Peanut (ArachishypogaeaL)[J]. Genetically Modified Organisms in Food,2016,27:305-311.
[4] Savage, G.P. and Keenan, J.I. The composition and nutritive value of groundnut kernels[M]∥ In: J. Smart (Editor), The Groundnut Crop: A Scientific Basis for Improvement. Chapman and Hall, London, 1994:173-213.
[5] 张 俊,刘 娟,臧秀旺,等.不同生育时期水分胁迫对花生生长发育和产量的影响[J].中国农学通报,2015,31(24):93-98.
(4)在相同的灌水技术参数组合下,不同灌水时期的灌水质量评价指标差异性均达到了中等变异程度,优化灌水技术参数时应该充分考虑这种时间变异性因子的作用。
□
参考文献:
[1] 李益农,许 迪,李福祥. 影响水平畦田灌溉质量的灌水技术要素分析[J]. 灌溉排水,2001,20(4):10-14.
[2] Smith R J,Raine S R,Minkevich J.Irrigation application irrigated cotton[J]. Agricultural Water Management, 2005,71(2):117-130
[3] 朱 艳,缴锡云,王维汉,等.畦灌土壤入渗参数的空间变异性及其对灌水质量的影响[J].灌溉排水学报,2009,28( 3) :46-49.
[4] 白美健,许 迪,李益农. 不同微地形条件下入渗空间变异对畦灌性能影响分析[J].水利学报,2010,41(6):732-738.
[5] 王维汉,缴锡云,朱 艳,等. 畦灌糙率系数的变异规律及其对灌水质量的影响[J]. 中国农学通报, 2009,25(16):288-293.
[6] 李益农,许 迪,李福祥.田面平整精度对畦灌系统性能影响的模拟分析[J].农业工程学报,200I,17(4):43-48.
[7] Oyonarte N A, Mateos L. Accounting for soil variability in the evaluation of furrow irrigation[J]. Transactions of the ASAE, 2003,46(1):85-94.
[8] 聂卫波,费良军,马孝义.基于土壤入渗参数空间变异性的畦灌灌水质量评价[J].农业工程学报,2012,28(1):100-105.
[9] 聂卫波,费良军,马孝义. 土壤入渗特性和田面糙率的变异性对沟灌性能的影响[J]. 农业机械学报,2014,45(1):108-114.
[10] 蔡焕杰,徐家屯,王 健,等. 基于WinSRFR 模拟灌溉农田土壤入渗参数年变化规律[J].农业工程学报,2016,32(2):92- 98.
[11] 李世瑶,蔡焕杰,陈新明. 基于主成分分析的畦灌质量评价[J]. 农业工程学报,2013,29(24):86-93.
[12] Bautista E, Clemmens A J, Strelkoff T S, et al. Modern analysis of surface irrigation systems with WinSRFR [J]. Agricultural Water Management, 2009,96(7):1 146-1 154.
[13] USDA-ARS (US Department of Agriculture, Agricultural Research Service).WinSRFR4.1 User Manual[M]. Arid land agricultural research center, maricopa, AZ, USA, 2012.
[14] 聂卫波,费良军,马孝义. 畦灌灌水技术要素组合优化[J].农业机械学报,2012,43(1):83-88.
猜你喜欢
农业工程学报(2022年3期)2022-04-16
西南农业学报(2021年7期)2021-08-23
节水灌溉(2021年5期)2021-05-26
生态学报(2021年6期)2021-04-25
农业工程学报(2021年3期)2021-04-15
节水灌溉(2020年9期)2020-09-16
中国农业科技导报(2020年12期)2020-03-15
新课程·下旬(2017年7期)2017-08-14
西北林学院学报(2015年5期)2015-01-03
植物营养与肥料学报(2011年2期)2011-10-26
