
家蚕LPH基因的生物信息学分析及表达模式研究
2016-03-16 06:43:00黄飞飞常平安
安徽农业科学 2016年3期
黄飞飞, 常平安
(重庆邮电大学,重庆 400065)
家蚕LPH基因的生物信息学分析及表达模式研究
黄飞飞, 常平安
(重庆邮电大学,重庆 400065)
摘要[目的]对家蚕LPH基因进行系统的分析。 [方法]以家蚕基因组数据和EST数据为基础,用比较基因组学方法从基因组层次系统地鉴定家蚕LPH基因,对基因的结构、染色体分布、基因进化以及与其他模式昆虫的LPH基因进行比较分析,并利用基因芯片数据对LPH基因在家蚕5龄3 d组织中的表达情况进行研究。 [结果]家蚕LPH是一个基因家族,包含19个直向同源基因,分布于6条染色体上,系统发生树分析表明它们主要分成二大类群;基因芯片数据分析表明家蚕LPH基因具有不同的表达模式,推测可能具有不同的功能,其中绝大部分基因在中肠组织中有表达,推测其可能参与黄酮类物质在家蚕体内的转运途径。[结论]该研究为家蚕LPH自身功能的研究提供分子基础,更重要的是为研究其他昆虫的LPH基因提供参考。
关键词乳糖酶-根皮苷水解酶;生物信息学;表达模式;功能分析
Bioinformatics Analysis and Expression Study of SilkwormLPHsGene
HUANG Fei-fei,CHANG Ping-an (Chongqing University of Posts and Telecommunications,Chongqing 400065)
Abstract[Objective] To systematically analyze the silkwormLPHgene.[Method] Based on silkworm genome sequences and EST data,silkwormLPHgene was identified at genome layer by comparative genomics method.The genetic structure,chromosome distribution,genetic evolution andLPHgene of other model insects were comparatively analyzed.SilkwormLPHgene expression in 3-day-old fifth-larvae tissues was studied by gene chip data.[Result] SilkwormLPHwas a gene family,which contained 19 orthologous genes and was distributed on six chromosomes.Phylogenetic analysis showed that they were mainly divided into two groups.Analysis of gene microarray data indicated that silkwormLPHgene had different expression patterns that might have different functions.But the vast majority of genes were expressed in intestinal tissues,which might participate in the transport pathway of flavonoids in the silkworm.[Conclusion] This research provides molecular basis for the research on silkwormLPHfunction,and offers important references for theLPHof other insects.
Ker wordsLactase-phlorizin hydrolase; Bioinformatics; Expression pattern; Function analysis
乳糖酶-根皮苷水解酶(lactase-phlorizin hydrolase,LPH)是一种存在于肠上皮细胞微绒膜上的糖蛋白,具有乳糖酶和根皮苷水解酶活性,在碳水化合物的消化中起重要作用。关于LPH的研究,在哺乳动物中研究较深入。编码人类LPH基因位于第2号常染色体上[1],由1 927个氨基酸组成[2-3]。人类中LPH主要表现为2种类型,一种类型为持续高活性,另一种类型则为非持续高活性。后一种表型的人由于其LPH表达水平低,不能将乳糖分解为葡萄糖和半乳糖,未被消化的乳糖在小肠内堆积,并被肠道细菌发酵而引起恶心、呕吐、腹胀、腹痛和腹泻等一系列症状,称为乳糖不耐受症,这是临床上最常见的胃肠病症之一[4-6]。这种LPH的非持续高活性表型自从被发现以来一直受到研究者的普遍关注,并在其他哺乳动物如鼠、兔、猪等中也开展了相关研究[7-8]。此外有研究指出哺乳动物LPH可参与水解黄酮苷类化合物[9]。与哺乳动物相比,关于昆虫LPH的研究很少。笔者前期从家蚕(Bombyxmori)基因组中成功鉴定了一个LPH基因,命名为BmLPH014192[10]。笔者以家蚕基因组序列和大规模EST数据为基础,用比较基因组学方法从基因组层次系统地鉴定了家蚕LPH基因,并对基因的结构、染色体分布、进化以及与其他模式昆虫的LPH基因进行了比较。该研究不仅为家蚕LPH自身功能的研究提供分子基础,更重要的是为研究其他昆虫的LPH基因提供参考。
1材料与方法
1.1数据库和基因组序列采用的家蚕基因组数据库(http://silkworm.swu.edu.cn/silkdb/)是由中日双方合作测序、组装和注释的9倍覆盖度的全基因组数据。家蚕全基因组oligo芯片数据来源于http://silkworm.swu.edu.cn/。果蝇(Drosophilamelanogaster)、按蚊(Anophelesgambiae)和蜜蜂(Apismellifera)的编码基因序列下载自flybase homopage,Ensemble (AgamP 3.45) and BeeBase (release 2)。
1.2工具软件信号肽预测 (http://www.cbs.dtu.dk/services/SignalP/);跨膜区预测 (http://www.cbs.dtu.dk/services/TMHMM-2.0/);预测基因的外显子、内含子结构采用Sim 4(http://pbil.univ-lyon1.fr/sim4.php);Blast、Clustal X 1.83、Bioedit、MEGA 4.0、PAUP 4.0和PHYLIP。
1.3家蚕LPHs基因的鉴定从GenBank数据库中下载人类、老鼠、兔子LPH基因的蛋白质序列,以这些已知的LPH序列为检索序列(query)与预测的家蚕蛋白质数据库进行BLASTP比对,同时与家蚕9倍基因组数据库作TBLASTN检索[11],根据比对结果收集E值较高的候选基因。采用SMART软件预测这些候选基因编码蛋白质序列的功能域。果蝇、按蚊和蜜蜂的LPH基因也是按照相同的方法进行鉴定。
1.4系统发生树的构建进化树采用家蚕、果蝇、按蚊和蜜蜂4个物种LPH基因的功能域序列构建,将LPH的功能域蛋白质序列用ClustalX[12]进行多序列联配。进化树主要采用MEGA4.0[13]软件中的邻近方法[14]构建,1 000次抽样分析。通过构建进化树推测家蚕LPH基因可能的进化。
1.5EST与芯片数据分析以预测的LPH基因蛋白质序列与家蚕EST数据库进行BLASTN比对,比对结果中序列一致性大于95%且E值小于1e-20的EST作为该基因的表达序列标签。
利用家蚕基因芯片数据分析LPH基因在家蚕5龄3d组织(精巢、卵巢、脂肪体、中肠、头、血液、马氏管、体壁、丝腺)中的转录情况。
2结果与分析
2.1家蚕LPH基因的鉴定从美国国立生物技术信息中心NCBI网站 (http://www.ncbi.nlm.nih.gov)下载人类、老鼠和兔子的LPH基因序列,与家蚕基因组数据库进行同源性比对。经过比对分析,最终从家蚕基因组中鉴别出19个LPH基因,基因的性质及其探针号见表1。功能域分析表明这些基因都具有名称为Glyco-hydro-1的结构域,属于水解酶1超家族。分别从果蝇、按蚊和蜜蜂基因组中鉴别出1、6和1个LPH基因。与果蝇、按蚊、蜜蜂基因组相比,家蚕基因组具有更多的LPH基因。对家蚕所有鉴定的LPH基因进行跨膜区分析,结果表明家蚕所有LPH基因均无跨膜区域。
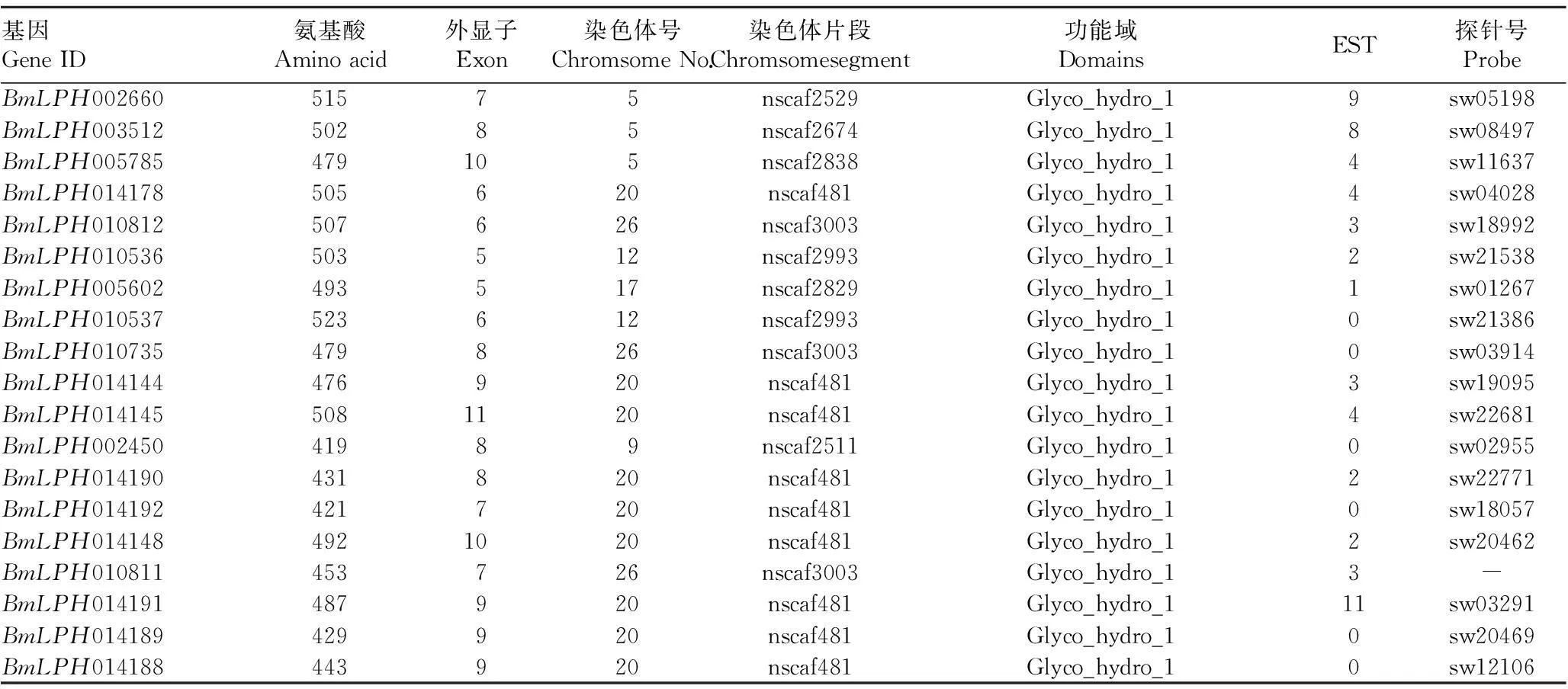
表1 家蚕LPH基因
2.2家蚕LPH基因家族的系统发生分析图1表明,4个物种27个LPH基因主要分成三大类群,将其命名为I~Ⅲ群。I群只包含家蚕的8个LPH基因,这些基因均位于家蚕20号染色体上,并在染色体上呈串联重复排列。II群只包含家蚕的7个LPH基因,这些基因分别位于4条不同的染色体上。其中BmLPH010536和BmLPH010537基因位于12号染色体;BmLPH010812、BmLPH010811和BmLPH010735基因位于26号染色体;BmLPH014178和BmLPH005602基因分别位于20号和17号染色体。Ⅲ群是同属双翅目昆虫的按蚊和果蝇的LPH基因聚在一起,包含果蝇的1个LPH基因和按蚊的6个LPH基因。家蚕的BmLPH003512、BmLPH002660和BmLPH002540基因,蜜蜂的Am18896-PA基因以及按蚊的Ag000481-PA基因均单独成一分支,均没有与上述三大类群聚在一起。

图1 家蚕、果蝇、按蚊、蜜蜂LPH构建的进化树Fig.1 Phylogentic tree of LPH in B.mori,D.Melanogaster,A.Gambiae and A.mellifera
与果蝇1个LPH基因、按蚊6个LPH基因和蜜蜂1个LPH基因相比,家蚕19个LPH基因数目明显增多。I群和II群是家蚕LPH特有的类群,共有15个LPH基因;Ⅲ群是双翅目昆虫所特有的类群,只包含果蝇和按蚊的LPH基因。在Ⅲ群里,果蝇的CG9701-PA和按蚊的Ag006425-PA可能是直系同源基因,节点处具有较高的自展值为100%。在II群中,BmLPH010536 和BmLPH010537在12号染色体上串联重复,BmLPH010812和BmLPH010811在26号染色体上串联重复,因此它们可能是通过位点复制而产生的。这些相似的现象也存在于I群中。家蚕的LPH基因主要分成两大类群I群和II群,没有与其他昆虫的LPH基因聚在一起,是家蚕所特有的类群,每个群上的LPH基因都表现出较高的序列相似性,表明该家族在基因进化过程中发生了基因重复事件。家蚕基因组中的大部分LPH基因可能经历了种族特异性的膨胀。
2.3家蚕LPH基因家族在基因组上的分布LPH基因常呈簇分布于基因组中。已鉴别的家蚕19个LPH基因位于6条染色体上。不同染色体上LPH基因数目从1~9个不等,位于相同染色体上的LPH基因表现出较高的序列相似性。20号染色体上共具有9个LPH基因,是主要的一大类群;此外5号和26号染色体都包含3个LPH基因,12号染色体包含2个基因;而9号和17号染色体均只包含1个基因。家蚕LPH基因家族存在基因转座现象,如有些位于相同染色体上的基因却分布在不同的分支上,这可能是基因转座产生的。基因BmLPH014178虽然位于20号染色体上,但其并没有与位于该染色体的其他LPH基因聚在一起。BmLPH005602与BmLPH010811和BmLPH005602聚在一起,但它们却位于不同的染色体上。BmLPH014178、BmLPH010536和BmLPH010537基因亦是如此。
2.4家蚕LPH基因在家蚕5龄3 d幼虫组织中的表达情况
2.4.1基于EST数据的表达分析。EST数据分析表明,家蚕LPH基因中有13个基因具有EST证据。家蚕LPH基因主要在某些组织中具有转录活性,包括中肠、丝腺、马氏管等。不同群的LPH基因,组织分布存在差异;而且位于相同群的LPH基因,组织表达情况也不同。
2.4.2基于芯片数据的表达分析。由家蚕5龄3 d的各个组织(包括精巢、头、卵巢、脂肪体、体壁、马氏管、血液、丝腺和中肠)组成家蚕全基因组的组织表达谱芯片。芯片数据分析表明,19个家蚕LPH基因中有18个基因有探针序列。将这18个LPH基因的不同组织芯片数据构建了聚类图。由图2可知,绝大多数基因具有不同的表达模式,其中16个基因在中肠组织中具有转录活性;BmLPH014178、BmLPH014190、BmLP002660等基因在家蚕组织中广泛表达;BmLPH010735、BmLPH010536、BmLPH010537、BmLPH002450、BmLPH014192、BmLPH014188呈组织特异性表达模式,只在中肠组织中有表达;BmLPH014148只在马氏管中有表达,而其他基因在某几个特定的组织中有表达。

注:红色代表阳性;黑色代表0;绿色代表阴性Note: Red color represented positive; black color represented zero; and green color represented negative.图2 芯片数据分析家蚕LPH基因在5龄3 d幼虫组织中的表达水平Fig.2 Expression levels of silkworm LPH genes in different tissues of three-day-old fifth-larvae tissues by microarray analysis
3结论与讨论
3.1家蚕LPH基因的数目 利用已组装的家蚕全基因组数据,通过生物学信息分析,在家蚕基因组中共鉴定出19个乳糖酶-根皮苷水解酶基因(LPH)。家蚕与果蝇(1个LPH)、按蚊(6个LPH)、蜜蜂(1个LPH)等其他昆虫相比,具有更多的LPH基因。
研究表明,哺乳动物LPH除参与乳糖水解作用外,还可参与水解黄酮苷类化合物。黄酮苷类的分子质量较大,其水溶性和脂溶性较差,很难靠被动扩散透过小肠上皮细胞。因此,黄酮苷类需要先被肠细胞存在的酶代谢成为苷元才能被吸收,而LPH就参与此种代谢[9]。与果蝇、按蚊、蜜蜂等昆虫相比,家蚕的唯一取食植物——桑叶含有较多的黄酮类化合物,家蚕LPH基因数目发生了膨胀且产生了家蚕特有的LPH基因,可能与桑叶中的黄酮类物质在家蚕体内的转运有关。推测,为了更好地吸收桑叶中的黄酮苷类物质,极有可能家蚕LPH基因数目发生了膨胀。
3.2家蚕LPH基因的结构迄今为止,研究最多的是哺乳动物LPH基因,研究表明哺乳动物LPH仅在小肠细胞中表达,在小肠的起始——空肠中段的含量最丰富,向两端降低,使得回肠末段和十二指肠近段的表达活性最低[15]。人LPH基因由1 927个氨基酸组成,成熟型LPH嵌入小肠微绒毛中,酶蛋白的羧基端有一个跨膜结构域使之可在小肠刷状缘形成二聚体,成为有活性的LPH[16]。通过对下载的兔、鼠LPH的结构分析表明,其蛋白羧基端都存在跨膜区域,而该研究对鉴别的家蚕LPH基因分析表明其不存在跨膜区域,推测其活性作用的方式可能与哺乳动物有所不同。
所鉴别的家蚕19个LPH基因分布于6条染色体上,系统发生树分析表明这些基因主要分为二大类群。每个群上的基因都表现出较高的序列相似性,表明在这个家族进化过程中发生了几个基因的重复事件。这些重复事件包括串联事件,重复的拷贝相互之间临近;此外还包括转座事件,一个拷贝通过转座位于其他不同的染色体上。2种类型的重复事件在基因BmLPH010536、BmLPH005602中可以看到。
3.3家蚕LPH基因的功能家蚕幼虫5龄3 d的组织芯片数据分析表明,家蚕LPH基因表现出不同的表达模式,表明这些基因可能具有不同的功能。绝大部分家蚕LPH基因在中肠组织有表达,这与LPH的基本功能一致,表明LPH极有可能参与黄酮类化合物在家蚕体内的代谢。
家蚕幼虫从它们唯一的寄主植物——桑叶中摄取黄酮类化合物,部分黄酮类化合物转运到生成的茧中,但从桑叶中分离到的黄酮类物质与从家蚕茧层中分离到的黄酮类物质在结构上不同。桑叶中的黄酮苷类物质主要是3-O-糖苷,蚕茧中的黄酮苷类物质主要是5-O-糖苷[17]。鉴于LPH参与哺乳动物体内黄酮类物质的转运,推测在家蚕中,首先是位于中肠上皮的LPH将黄酮苷类物质水解成分子量较小的苷元便于中肠的吸收,后被位于中肠、血液或丝腺中的尿苷二磷酸-糖基转移酶(UGT)所催化[18],从而生成家蚕体内特有的黄酮苷类物质。因此在中肠中高表达或特异表达的LPH基因如BmLPH010735、BmLPH010536、BmLPH010537等可能对家蚕从桑叶中更好地吸收黄酮类化合物起重要作用。此外,BmLPH014192基因已证明在绿色茧品种和白色茧品种的幼虫中肠组织中出现序列选择性剪接[10],也间接表明LPH很有可能参与蚕体内黄酮类化合物的转运机制。
家蚕LPH基因除在中肠表达外,在其他组织也有表达。BmLPH014178 基因在家蚕5龄3 d的各个组织中都有表达,表明该基因可能在家蚕发育过程中起到看家基因的作用。此外个别基因在精巢、卵巢、头、脂肪体、体壁、马氏管和丝腺中有表达,而在这些组织中表达的基因所起的作用尚不清楚,需进一步研究。
总之,家蚕19个LPH基因表达模式多样,表明可能担负的功能不同。对家蚕LPH基因的生物信息学分析及其表达模式的研究可为以后LPH基因的研究提供重要参考。
参考文献
[1] POTTER J,HO M W,BOLTEN H.Human lactase and the molecular basis of lactase persistence[J].Genet,1985,23(5/6):423-439.
[2] KRUSE T A,BOLUND L,GRZESCHIK K H.The human lactase-phlorizin hydrolase gene is located on chromosome 2[J].FEBS Lett,1988,240(1/2):123-126.
[3] HARVEY C B,FOX M F,JEGGO P A.Regional localization of the lactase-phlorizin hydrolase gene,LCT,to chromosome 2q21[J].Ann Hum Genet,1993,57(3):179-185.
[4] SWAGERTY D L JR,WALLING A D,KLEIN R M.Lactose intolerance[J].Am Fam Physician,2002,65(9):1845-1850.
[5] DAHLQVIST A,HAMMOND J D,GRANE R K.Intestine lactase deficiency and lactose intolerance in adults:Preliminary report[J].Gastroenterol ,1963,45(4):488-491.
[6] SAHI T,ISOKOSKI M,JUSSILA J.Recessive inheritance of adult-type lactose malabsorption[J].Lancet,1973,2(7833):823-826.
[7] BURRIN D G,STOLL B,FAN M Z.Oral IGF-I alters the posttranslational processing but not the activity of lactase-phlorizin hydrolase in formula-fed neonatal pigs[J].J Nutr,2001,131(9):2235-2241.
[8] HE W,JI Y,HUANG Y.Molecular cloning of a novel cDNA from Mus muscular BALB/c mice encoding glycosyl hydrolase family 1:A homolog of human lactase-phlorizin hydrolase[J].Biomed Envir Sci,2006,19(5):340-345.
[9] SESINK A L,ARTS I C,FAASSEN-PETERS M,et al.Intestinal up take of quercetin-3-glucoside in rats involves hydrolysis by lactase phlorizin hydrolase[J].J Nutr,2003,133(3):773.
[10] 黄飞飞,赵小靓,鲁成.家蚕乳糖酶-根皮苷水解酶基因BmLPH014192的表达和序列选择性剪接分析[J].蚕业科学,2013,39(1):21-27.
[11] ALTSCHUL S F,MADDEN T L,SCHAFFER A A,et al.Gapped BLAST and PSI-BLAST:A new generation of protein database search programs[J].Nucleic Acids Res,1997,25(17):3389-3402.
[12] THOMPSON J D,GIBSON T J,PLEWNIAK F,et al.The ClustalX windows interface:Flexible strategies for multiple sequence alignment aided by quality analysis tools[J].Nucleic Acids Res,1997,25(24):4876-4882.
[13] TAMURA K,DUDLEY J,NEI M,et al.MEGA4:Molecular evolutionary genetics analysis (MEGA) software version 4.0[J].Mol Biol Evol,2007,24(8):1596-1599.
[14] HOWE K,BATEMAN A,DURBIN R.QuickTree:Building huge Neighbour-Joining trees of protein sequences[J].Bioinformatics 2002,18(11):1546-1547.
[15] MESONERO J E,GLOOR S M,SEMENZA G.Processing of human intestinal prolactase to an intermediate form bu furin or by a furin like proprotein convertase[J].J Biol Chem,1998,273(45):29430-29436.
[16] BOLL W,WAGNER P,MANTEI N.Structure of the chromosomal gene and cDNAs coding for lactase phlorizin hydrolase in humans with adult-type hypolactasia or persistence of lactase[J].Am J Hum Genel,1991,48(5):889-902.
[17] YASUMORI T,NAKAJIMAB K,NAGAYASUA K,et al.Flavonoid 5-glucosides from the cocoon shell of the silkworm,Bombyxmori[J].Phytochemistry,2002,59:275-278.
[18] RAUSELL C,LLORCA J,REAL M D.Separation by FPLC chromatofocusing of UDP-glucosyltransferases from three developmental stages ofDrosophilamelanogaster[J].Arch Insect Biochem Physiol,1997,34(3):347-358.
收稿日期2015-12-30
作者简介黄飞飞(1980- ),女,山东即墨人,讲师,博士,从事生物化学与分子生物学方面的研究。
基金项目重庆市自然科学基金项目(cstc2013jcyjA10005;KJ1400424)。
中图分类号S 188+.1
文献标识码A
文章编号0517-6611(2016)03-111-04
猜你喜欢
食品与机械(2017年4期)2017-07-05 14:46:17
食品与生物技术学报(2017年5期)2017-04-08 10:07:27
中国教育信息化·基础教育(2016年10期)2016-12-20 20:09:27
今传媒(2016年11期)2016-12-19 11:35:50
电脑知识与技术(2016年27期)2016-12-15 19:19:25
中国科技纵横(2016年17期)2016-11-30 21:40:27
科技资讯(2016年19期)2016-11-15 10:39:12
电子技术与软件工程(2016年18期)2016-11-14 00:49:04
戏剧之家(2016年20期)2016-11-09 23:08:07
科技视界(2016年4期)2016-02-22 20:53:20
