
毛尖蘑粗多糖对H22荷瘤小鼠P53表达的影响
2016-02-06 03:36罗文哲吕冬霞金岳雷金绍静王丹丹
中国老年学杂志 2016年24期
罗文哲 吕冬霞 金岳雷 金绍静 张 虎 王丹丹
(佳木斯大学基础医学院,黑龙江 佳木斯 154007)
毛尖蘑粗多糖对H22荷瘤小鼠P53表达的影响
罗文哲 吕冬霞 金岳雷 金绍静 张 虎 王丹丹
(佳木斯大学基础医学院,黑龙江 佳木斯 154007)
目的 研究毛尖蘑子实体粗多糖(MJMP)对H22荷瘤小鼠P53表达的影响。方法 构建H22小鼠模型,随机分组,观察记录小鼠生存状态,采用免疫组化法检测肿瘤组织P53蛋白的表达;RT-PCR法检测肿瘤组织P53 mRNA的表达。结果 免疫组化显示,MJMP可减少突变型P53蛋白阳性细胞百分率,与阴性对照组比较差异显著(P<0.01);RT-PCR法表明,经毛尖蘑粗多糖处理后,P53 mRNA表达与阴性对照组比较明显增强(P<0.01)。结论 MJMP对H22肿瘤的抑制作用可能与P53 mRNA及蛋白高表达有关。
毛尖蘑多糖;P53;凋亡
近些年人们发现中药多糖及其复合物具有抗肿瘤、增加免疫功能等作用,其中尤以真菌多糖的研究最为活跃〔1~6〕。本研究将毛尖蘑子实体粗多糖(MJMP)作用于H22荷瘤小鼠,探究其抑瘤作用机制。
1 材料与方法
1.1 材料 H22肝癌细胞株由中国医学科学院肿瘤研究所提供。毛尖蘑购于大兴安岭呼玛县韩家园林业局松涛鹿苑公司;昆明种近交系雌性小鼠,体重18~22 g,购于佳木斯大学实验动物中心(普通级,普通环境饲养,许可证号:SYXK黑2006-004);环磷酰胺(CTX)为江苏恒瑞医药股份有限公司产品;β-actin多克隆抗体来自北京博奥森生物有限公司。 Trizol试剂购于Gibco公司;总RNA提取试剂盒、反转录 cDNA合成试剂盒购于大连Takara生物公司。
1.2 方法
1.2.1 动物实验分组与给药处理 小鼠常规建模(空白对照组不接种肿瘤),随机分组,每组10只,24 h后以灌胃方式给予不同浓度药物,用药量均为0.2 ml。空白对照组、阴性对照组均予生理盐水0.2 ml,阳性对照组予CTX 25 mg/kg,高剂量组予高浓度MJMP 40 mg/ml,低剂量组(L)予低浓度MJMP 20 mg/ml。连续给药14 d,观察小鼠生存状态。
1.2.2 免疫组化法检测肿瘤组织P53蛋白表达 按说明书要求常规制备小鼠肿瘤组织免疫组化切片。显微镜下随机选择5个视野观察并计数,突变型P53阳性表达细胞为黄色。
1.2.3 RT-PCR方法检测肿瘤组织P53 mRNA的表达 按说明书要求以常规方法进行RT-PCR操作。PCR反应引物P53:上游5'-GAAACTACTTCCTGAAAACAACGA-3',下游5'-GCCTCACAACCTCCGTCAT-3';β-actin:上游5'-AATGGGTCAGAAGGACTCCTATGTGG-3',下游5'-CGCCTAGAAGCACTTGCGGTG-3'。应用凝胶成像系统扫描分析,依据各条带的光密度和面积对扩增产物进行定量分析,以 β-actin密度作为参考定量标准,计算P53/β-actin的值。
1.3 统计学方法 应用SPSS11.5 统计采用方差分析(AVONA),两两比较用Q检验。
2 结 果
2.1 H22荷瘤小鼠生存状态 将H22细胞悬液注射至小鼠右腋下,接种后第6天部分小鼠腋下肿瘤肉眼可见,7~14 d时肿瘤生长速度较快,通过肉眼观察及皮下触摸,阳性对照组肿瘤的生长速度及体积小于其他组。实验过程中,空白对照组小鼠活动自如、体重递增、精神状态佳;阴性对照组小鼠活动减少、蜷缩,体重减轻,精神萎靡,毛色光泽差;阳性对照组于给药7 d后小鼠活动减少,消瘦明显;高、低剂量组小鼠基本状态良好,体重少量增加。
2.2 P53蛋白表达 阳性对照组(22.65±4.53)和MJMP治疗组(H组28.49±5.62;L组32.84±7.65)与阴性对照组(65.23±9.52)的P53有显著差异(P<0.01)。
2.3 P53 mRNA的表达 P53基因在阴性对照组低表达(0.16±0.02),经过CTX和MJMP处理后,P53 mRNA表达明显增强,见图1。阳性对照组、高剂量组、低剂量组(L)P53 mRNA表达量分别为0.30±0.03、0.28±0.02、0.19±0.02。
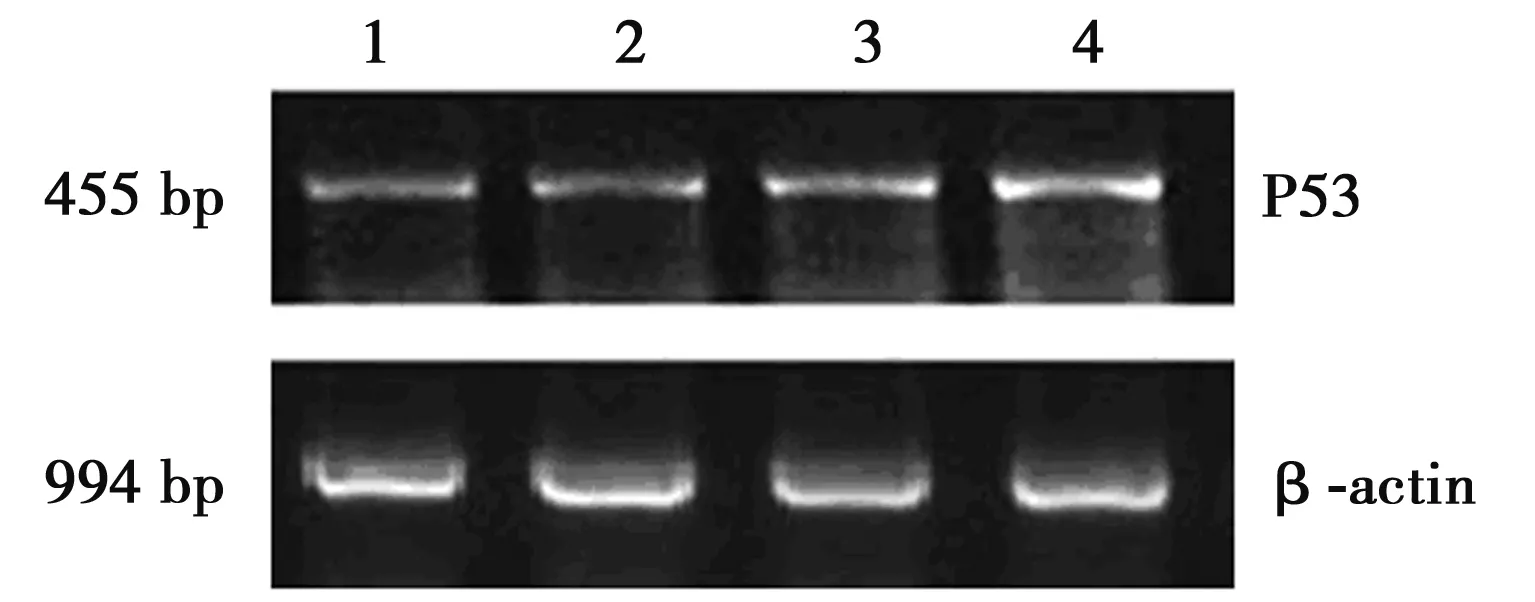
1、阴性对照组;2、低剂量组;3、高剂量组;4、阳性对照组图1 RT-PCR法检测H22小鼠肿瘤组织P53 mRNA 的表达
3 讨 论
肝癌是由肝细胞过度增殖及细胞凋亡异常而导致的疾病,肝癌的发生、发展是多基因共同作用的结果,是由细胞凋亡基因调控的。因此,综合多基因的评价对药物治疗肝癌的疗效具有重要意义。抑癌基因是抑制细胞生长的重要因素之一,是细胞生长的制动器。P53是与肿瘤相关性最高的基因之一,被称为“基因组卫士”。
P53基因能够抑制肿瘤细胞的生长和分裂,在人类肿瘤中常发生变化,可分为野生型和突变型两种。在正常情况下,野生型P53为其存在形式,但表达水平较低,很难在细胞中发挥作用。当细胞受到刺激后,P53表达才得到增高,诱导细胞进入G1静止期,形成细胞周期阻滞,抑制细胞生长,修复损伤细胞〔7〕,野生型P53也可诱发变异细胞凋亡,控制癌前细胞的形成。野生型P53的半衰期仅几分钟,用免疫组化方法难以测出。当受到药物刺激时,野生型P53可被激活,提高其稳定性。在肿瘤发生过程中,野生型P53可转变为突变型P53蛋白。突变型P53失去对肿瘤细胞的抑制功能,减少肿瘤细胞的凋亡发生,从而导致细胞癌变。突变型P53稳定性明显增强,在细胞内聚集比正常细胞高上百倍,可用免疫组化方法检测到。因此,采用免疫组化方法检测到的P53基因均为突变型P53基因。
P53主要通过以下途径对细胞凋亡起到调控作用:①P53对外源性凋亡途径起到调节作用,P53能够激活死亡受体促进细胞凋亡,如Fas、DR4、DR5等〔8〕。②P53对内源性凋亡途径起到调节作用。线粒体是调控细胞内源性凋亡途径的主要部位,P53可对线粒体膜的Bcl-2家族蛋白起到调控作用,如对Bax/Bak的上游调控,从而诱导细胞凋亡〔9〕。
本试验结果提示MJMP能够减少H22肝癌细胞中突变型P53蛋白的表达,增加野生型P53的抑癌功能,减少H22肝癌细胞的增殖,可能诱导肿瘤细胞凋亡,提示MJMP抗肿瘤可能与增强小鼠的免疫功能相关。
1 刘玉红,王凤山.真菌多糖结构研究进展〔J〕.中国药学杂志,2007;42(8):561-4.
2 芦殿荣,祝彼得,芦殿香,等.香菇多糖对正常小鼠以及免疫抑制小鼠免疫功能的影响〔J〕.甘肃中医学院学报,2004;4(21):20-2.
3 周淑英.黄芪多糖对小鼠免疫功能调节作用的观察〔J〕.江苏中医,1994;15(12):41-2.
4 陈国荣,杨开颜,邵黎明,等.香菇多糖对大鼠腹腔巨噬细胞活性的影响〔J〕.温州医学院学报,1996;26(4):225-6.
5 杜泽英.香菇多糖对人淋巴细胞IL-2分泌及IL-2R受体表达双向调节作用〔J〕.中国实验临床免疫学杂志,1994;6(1):40.
6 常花蕾.金针菇多糖的免疫调节作用、抗肿瘤作用及其机制研究〔D〕.广州:南方医科大学,2009.
7 孙淑娟,张清媛.乳腺癌C-erbB-2 Ki-67 p53表达及意义〔J〕.当代医学(学术版),2008;(3):100-1.
8 Longley DB,Allen WL,McDermott U,etal.The roles of thymidylate synthase and p53 in regulating Fas-mediated apoptosis in response to antimetabolites〔J〕.Clin Cancer Res,2004;10(10):3562-71.
9 Alvarez S,Drane P,Meiller A.A comprehensive study of p53 transcriptional activity in thymus and spleen of gamma irradiated mouse:High sensitivity of genes involved in the two main apoptotic pathways〔J〕.Int J Radiat Biol,2006;82(11):761-70.
〔2016-05-09修回〕
(编辑 郭 菁)
佳木斯大学重点研究项目(No:Sz2013-002)
吕冬霞(1965-),女,教授,硕士生导师,主要从事肿瘤发生机制与生物学治疗研究。
罗文哲(1976-),男,副教授,硕士,主要从事肿瘤发生机制与生物学治疗研究。
R730.52
A
1005-9202(2016)24-6083-02;
10.3969/j.issn.1005-9202.2016.24.017
猜你喜欢
故事会(蓝版)(2020年7期)2020-07-21
浙江医学(2020年9期)2020-07-01
浙江中西医结合杂志(2019年4期)2019-05-05
浙江医学(2019年2期)2019-01-23
大众科学(2018年10期)2018-12-11
中国病理生理杂志(2018年9期)2018-09-27
贵茶(2018年6期)2018-05-30
农产品市场周刊(2017年18期)2017-06-07
中国继续医学教育(2015年1期)2016-01-06
癌症进展(2015年5期)2015-07-26
