
用药动作线索诱发海洛因戒断者的镜像神经活动:一项fMRI研究*
2016-02-01 22:11苏得权叶浩生
心理学报 2016年12期
苏得权 曾 红 陈 骐 叶浩生
(1广州大学心理系; 广州大学心理与脑科学研究中心, 广州 510003) (2华南师范大学心理学院, 广州 510631)
1 问题的提出
精神活性物质能够直接或间接地引起纹状体(包括伏隔核)的多巴胺(Dopamine, DA)浓度增加,产生用药后的急性反应, 给用药者带来欣快感(Koob, 1992), 使用药者对精神活性物质逐渐产生依赖。精神活性物质对于依赖者来说是非条件刺激,药物依赖者重复用药, 在条件(conditioning)作用下,经常在用药情境中出现的中性刺激就成为相关线索, 如药物、用药动作、用药场所, 以及当事人的心境等。研究表明, 相关线索同样可以引起纹状体DA浓度的增加(Owesson-White et al., 2009)。
相关线索和吸食毒品都能引起纹状体DA释放的增加, 这两种情况下多巴胺系统的活动有何差异,为何吸食毒品后的DA释放能够带来足够的欣快感,相关线索引发的多巴胺系统的活动非但不能满足药物依赖者的期望, 还引发了药物寻求行为。这种差异并非是因为急性用药引起的DA释放量多, 相关线索引起DA的释放量少导致的(Saunders, Yager,& Robinson, 2013)。可见, DA奖赏系统还不能充分解释相关线索诱发作用下的药物寻求行为。
药物依赖者在相关线索诱发作用下产生心理渴求, 其负责动机、记忆(Grant et al., 1996)和执行控制功能的脑区会产生显著激活, 如中脑奖赏系统(Saunders et al., 2013)和前额叶皮层(Wagner, Cin,Sargent, Kelley, & Heatherton, 2011)。除此之外, 前运动皮层和顶下小叶等脑区与物质依赖程度表现出显著的正相关(Smolka et al., 2006)。顶下小叶, 以及负责运动准备和动作模拟的前运动皮层和辅助运动皮层是镜像神经系统(mirror neuron system,MNS)的主要组成部分, 同时也是动作观察时相对活跃的脑区。
镜像神经系统, 也称为动作观察脑区(action observe neuron, AON), 具有动作表征作用。研究者使用手部动作图片和静止物品图片作为实验材料,寻找人类的镜像神经系统。研究发现, 观看手部抓取物体动作和观看静止物体两种条件下激活脑区相比较, 顶下小叶、颞上回、额下回和背外侧前运动皮层和中央前/后回会出现相应激活(Decety et al.,1997; Rizzolatti, Fadiga, Matelli, et al., 1996), 或者观看手部动作或可用手操作的工具图片时, 镜像神经系统也会出现激活(Grafton, Fadiga, Arbib, &Rizzolatti, 1997)。
镜像神经系统的活动受到观察者动作熟练程度的影响。研究发现, 羽毛球运动员(Wright, Bishop,Jackson, & Abernethy, 2011)、篮球运动员(Abreu et al., 2012)和剑术运动员(Kim et al., 2011)与非专业运动员相比, 在观看专业运动项目动作时, 专业运动员的AON出现了更显著的激活。专业舞蹈演员和不会跳舞的普通人在观看舞蹈动作的时候, 舞蹈演员AON出现了更为显著的活动(Calvo-Merino,Grèzes, Glaser, Passingham, & Haggard, 2006; Cross et al., 2012)。研究者认为, AON的活动是因为专业人员对熟悉动作的心理模拟引起的。
尼古丁成瘾者与不抽烟的人观看相同的抽烟动作, 前者的顶内沟前部、左侧额下回, 以及前运动皮层出现了更为显著的激活(Wagner et al., 2011)。这些脑区分别与熟练动作加工和运动计划等功能有关(Yalachkov & Naumer, 2011)。随着时间的延续,相关线索诱发的脑激活水平会逐渐提高。长期的海洛因依赖者在看到用药动作时, 他们的颞叶、枕叶、前/后扣带回、丘脑、小脑和左侧海马都出现了显著激活; 其中, 后扣带回和海马的激活水平与药物使用时间显著相关(Lou, Wang, Shen, & Wang, 2012)。可见, 成瘾者的镜像神经活动与成瘾行为之间也存在一定的关系。
药物依赖者在摆脱对毒品的生理依赖之后, 仍然存在心理渴求。相关线索能够重新点燃药物依赖者的心理渴求。目前, 尚未有针对海洛因戒断者镜像神经系统的研究。作者假设, 戒断后的海洛因依赖者对相关线索比较敏感, 与对照线索相比, 相关线索能够诱发戒断后的海洛因依赖者镜像神经系统更加强烈的活动, 对于健康人来说可能不存在这种情况。
2 方法
2.1 研究对象
选用戒断后的海洛因依赖者(abstinent heroin dependent individuals, AHD)和没有药物依赖史的健康人(no-drug use health participants, NDP)为研究对象。AHD共15人, 年龄30~50周岁, 初中或高中文化程度。药物依赖时间和戒断时间均为12个月以上, 无赌博和酗酒等不良嗜好。NDP共12人,来自社会不同领域的工作人员, 主要包括后勤服务人员, 医院护工等。两组被试在年龄、教育程度等人口学变量无显著差异(见表1)。
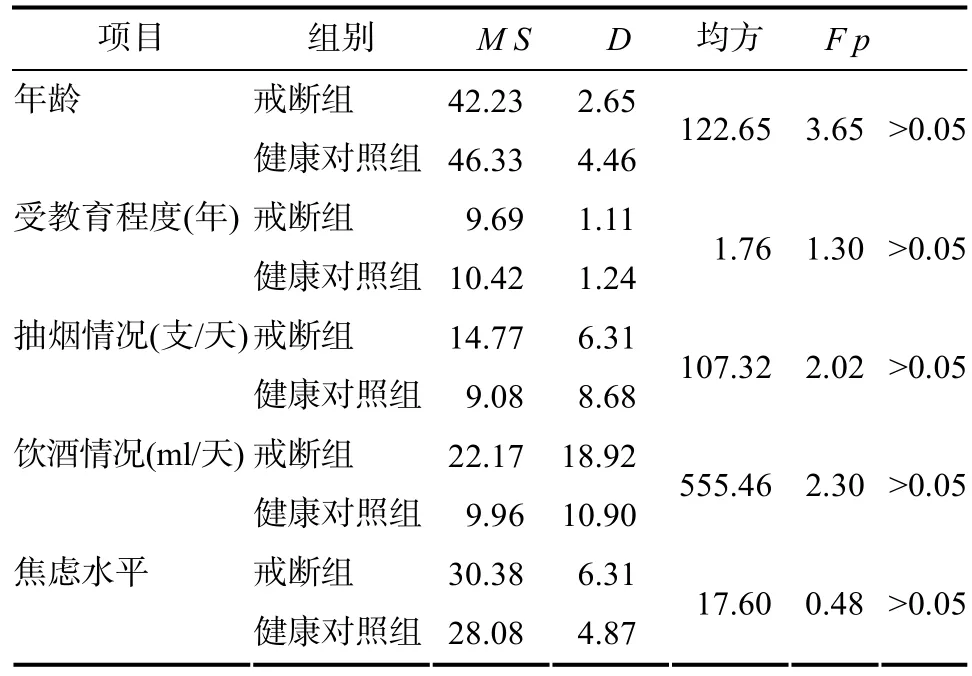
表1 被试基本信息表
所有被试在实验前填写“核磁共振(MRI)扫描被试检查单”, 符合磁共振检查要求的被试在正式实验前将被详细告知实验的过程, 并签署知情同意书。实验程序经伦理委员会审核通过, 在研究过程中, 严格按照预定的实验程序实施。
2.2 实验材料
相关线索包括静止的海洛因药物图片(药物线索)和吸食海洛因的动作图片(用药动作线索); 选用日常生活中不可食用的粉末状(对照药物线索)或颗粒状物体(如沙子)图片和日常行为动作图片(对照动作线索)作为对照线索。要求药物线索图片只包含药物本身(如海洛因粉末), 或者带包装的药物(小包装的海洛因), 画面中不能显示人物和人物动作; 用药动作线索图片显示的是吸食海洛因的情景。对照药物和对照动作图片的选取也按照同样的标准。所有图片统一转换为jpg格式, 分辨率72像素, 尺寸大小为1024×768的图片。图片筛选完毕后, 请戒毒康复机构的戒毒人员评价相关线索图片诱发心理渴求的程度, 评价者共计67人, 年龄39.27 ± 5.37岁, 教育程度10 ± 2.42年, 药物依赖时间11 ± 5.68年, 戒断时间19.3 ± 15个月, 主要吸食海洛因。要求他们对每一张图片诱发的心理渴求程度进行0~7级评分(0=这幅图片不能让我想起海洛因, 7=这幅图片能让我极其渴望使用海洛因)。删除评价得分为0的图片之后, 获得药物线索图片和用药动作线索图片各45张; 独立样本t检验结果显示, 用药动作图片评价得分显著高于药物图片(M
=2.36,M
=1.86;t
=4.14,df
=88,p
<0.001)。两类相关线索诱发心理渴求程度的能力均偏低, 不排除正在康复的戒断者有意掩饰线索诱发的心理反应。2.3 实验设计
采用2×2×2混合实验设计。因素一为被试间变量(group), 包括药物戒断和健康对照; 因素二为线索相关性(factor1), 根据线索刺激是否与药物依赖行为有关, 分为相关线索和对照线索; 因素三为线索类型(factor2), 包括实物线索和动作线索; 因素二与因素三均为被试内变量, 两个因素共产生4种组合, 分别为药物线索(drug)、用药动作线索(action)、对照药物线索(comparison drug, C-drug)和对照动作线索(comparison action, C-action), 如表2所示。

表2 线索刺激内容明细表
2.4 实验程序
采用block模式呈现刺激, 每个block中包含5个trail。每个trail中, 先呈现注视点“+”, 时间为500 ms, 接着出现线索刺激, 呈现时间为2000 ms,图片消失后, 屏幕中央呈现注视点, 时间为500 ms(为了减少被试在观看图片时的眼动, 图片中心也设置了注视点), 每个trail呈现时间为3000 ms, 整个block的呈现时间为15000 ms。刺激呈现过程中,对照线索block与相关线索block交替呈现。整个实验过程中, 被试头部不动, 眼睛注视屏幕中央的注视点。要求被试在看清楚图片上的内容后, 右手食指按键反应。
使用E-prime (2.0.8.22 professional, Copyright©1996-2005 psychology software Tools, http://www.pstnet.com)呈现线索刺激。为了保证程序调用图片时间的精确性, 图片大小不超过240 K, 呈现时间模式(timing mode)设定为“cumulative”, 并提前300 ms预加载下一张图片, 保证刺激呈现与磁共振扫描的同步。
2.5 数据的收集与处理
使用华南师范大学磁共振成像中心的西门子(SIEMENS MAGNETOM TrioTim syngo MR B17,3.0T)磁共振成像系统采集数据。扫描基线与前−后联合连线(AC-PC)平行, 采用梯度回波平面成像序列(echo planar Imaging, EPI)采集功能像。重复时间TR=2200 ms, 回波时间TE=30 ms, 翻转角FA=90°, 视野FOV=1320×1320 mm, 矩阵matrix=64 ×64, 隔层扫描, 覆盖全脑36层, 层厚slice thickness=3 mm。每个被试采集445~450个TR不等, 记录被试观看不同画面时的脑活动。
在Matlab 2010平台上, 采用SPM8工具箱对fMRI数据进行预处理。首先使用SPM8把采集到的DICOM文件转换为SPM可处理的NIFTI格式文件, 抽取被试的前442个TR, 进行头动校正, 删除头动大于1个voxel尺寸(3 mm × 3 mm × 3 mm)的被试数据; 然后进行配准, 再将配准后的图像分割为灰质、白质和脑脊液; 把分割得到的灰质归一化到MNI (montreal neurological institute, MNI)系统, 并以2 mm × 2 mm × 2 mm大小的体素重采样;最后使用全宽半高值为8 mm的高斯核函数进行平滑, 以降低空间噪声。通过以上步骤完成数据的预处理。
first-level参数模型包括4种条件, 分别为药物对照线索、动作对照线索、成瘾药物线索、用药动作对照线索, 把每个被试的错误反应和头动校正产生的6个头动函数作为一种条件放入模型中, 但不对它进行分析。采用简单线性模型构建每个被试4种实验条件的多重回归设计矩阵。运用Flexible factorial 设计进行second-level的模型建构, 模型中包括线索相关性和线索类型两个因素, 提取first-level中产生的4种条件下的contrast文件, 建立多重回归设计模型。模型估计后, 使用SPM软件作ANOVA分析, 得到4种线索条件下的全脑激活情况; 然后把两组被试4种条件下的脑激活数据建立在同一个模型中, 以健康组被试药物线索和用药动作线索条件下的激活脑区作为mask, 采用exclusive运算法则, 获得两组被试相关线索条件下脑激活情况的差异, 报告经FWE校正后p
<0.05的激活脑区。3 结果分析
3.1 药物线索激活脑区的组间比较
与健康组被试(NDP)相比较, 药物线索导致戒断后的海洛因依赖者(AHD)左侧枕叶、左侧扣带回和左侧楔前叶出现了更高水平的激活(见表3)。相同显著性水平下, NDP只有右侧枕叶出现了更为显著的激活(见图1)。
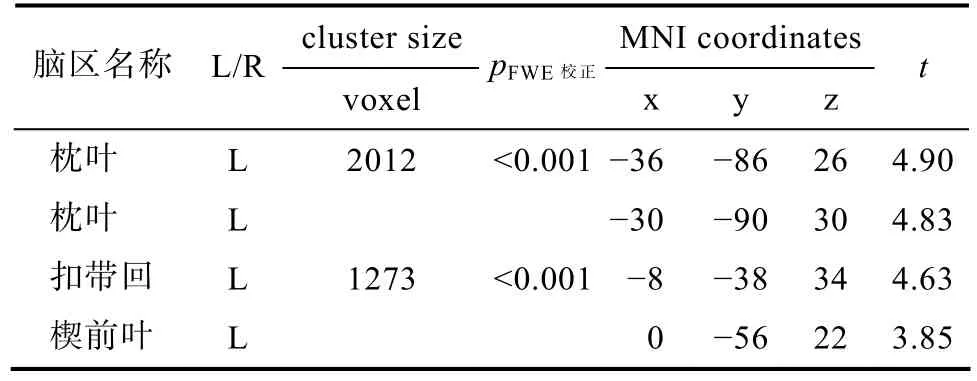
表3 药物线索条件下激活脑区的组间比较(AHD vs NDP)
3.2 戒断组和健康对照组镜像神经脑区分析
观看手部抓取物体动作和观看静止物体两种条件下激活脑区相比较, 可以获得一般动作的镜像神经脑区(van Elk, 2014)。两组被试在观看用药动作与静止药物两种条件下的脑区活动比较后, 得到用药动作线索任务引发的镜像神经活动, 如表4所示。
戒断后的海洛因依赖者用药动作与静止药物线索激活脑区比较[AHD (action vs drug)]显示, 枕叶和颞叶出现了极其显著的活动, 颞叶激活峰值点位于大脑左半球, 颞中回和颞上沟结合部位。双侧的顶下小叶, 以及右侧缘上回也出现了显著激活。丘脑和中脑系统在用药动作的镜像神经活动中也有激活。在p
< 0.05的情况下, 左侧海马, 左侧BA6区的额上回和额中回与中央前回结合部出现了显著激活的体素(−26/8/68, −38/−2/38), 颞下回和右侧额下回也出现了较多的激活体素, 双侧的中央前回也出现了激活(−36/−2/54, 34/−4/48)。用药动作与静止药物线索激活脑区比较后, 在p
<0.05水平下, 健康人只有双侧枕叶和颞下回出现了显著激活(如表4所示)。戒断后的海洛因依赖者观看对照动作与药物对照图片激活脑区比较[AHD (C-action vs C-drug)],出现了3个显著激活的cluster, 包括双侧的枕−颞联合皮层、顶叶和右侧的额下回脑区, 左侧脑激活区域较多(如表5所示)。左侧的cluster包括枕叶的枕中回、颞叶的颞中回, 以及顶下小叶, BA2/3区的中央后回, 顶上小叶、缘上回; 还包括BA4区的中央前回。右侧的cluster包括BA37区的枕叶−颞叶联合皮层, 颞中回、颞下回和颞上回也有激活; 右侧顶上小叶和顶下小叶也出现了显著激活。右侧BA45区的额下回三角部出现了显著激活。在p
< 0.05的前提下, 降低cluster size的大小, 在左侧BA45区额下回三角部(−56/20/16, cluster size=19)和左侧的中央前回(−34/−2/48, cluster size=19)也发现有显著激活的体素。健康人对照动作与对照药物图片激活脑区比较[NDP (C-action vs C-drug)], 出现了4个显著激活的cluster, 枕叶−颞叶联合皮层、右侧的顶上小叶和顶下小叶, 以及BA2区的中央后回, 左侧顶下小叶都出现了显著活动。NDP没有发现右侧额下回的显著激活。
3.3 AHD和NDP对照动作和用药动作激活镜像神经脑区的联合分析
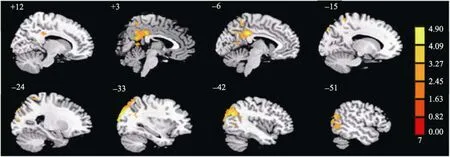
图1 药物线索条件下激活的脑区的组间比较(AHD vs NDP)
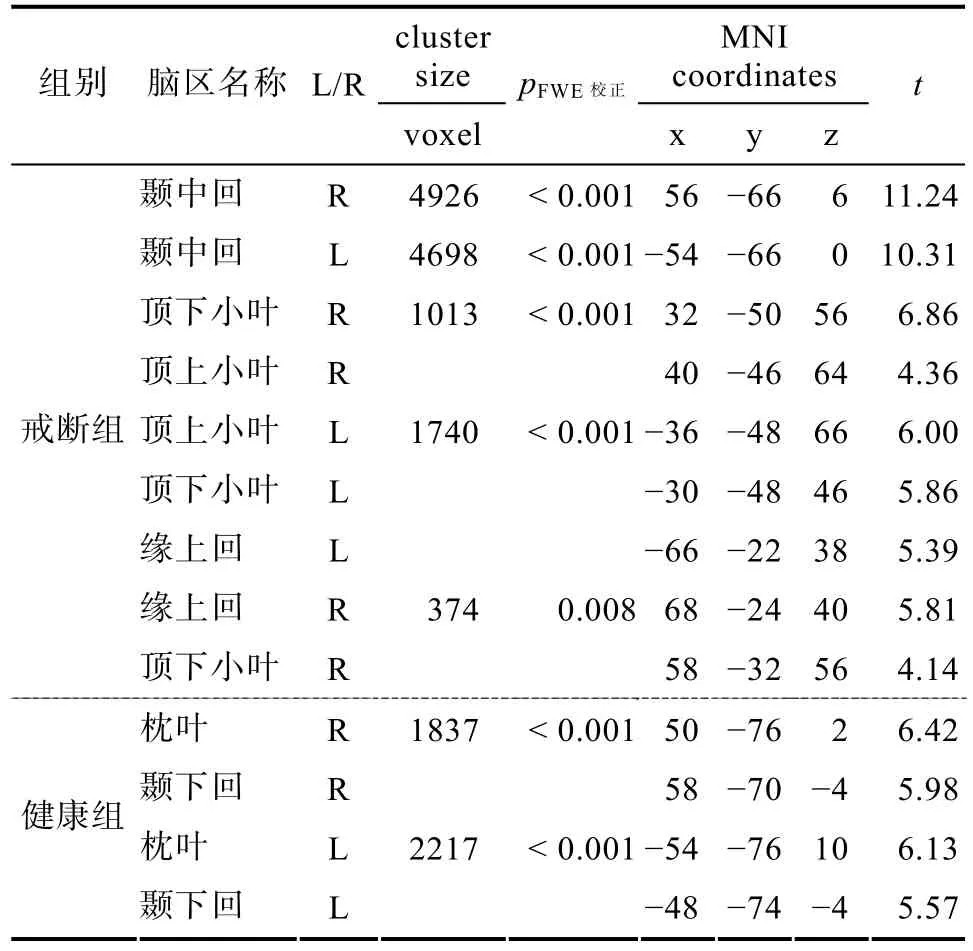
表4 戒断组和健康组用药动作引发的镜像神经脑区
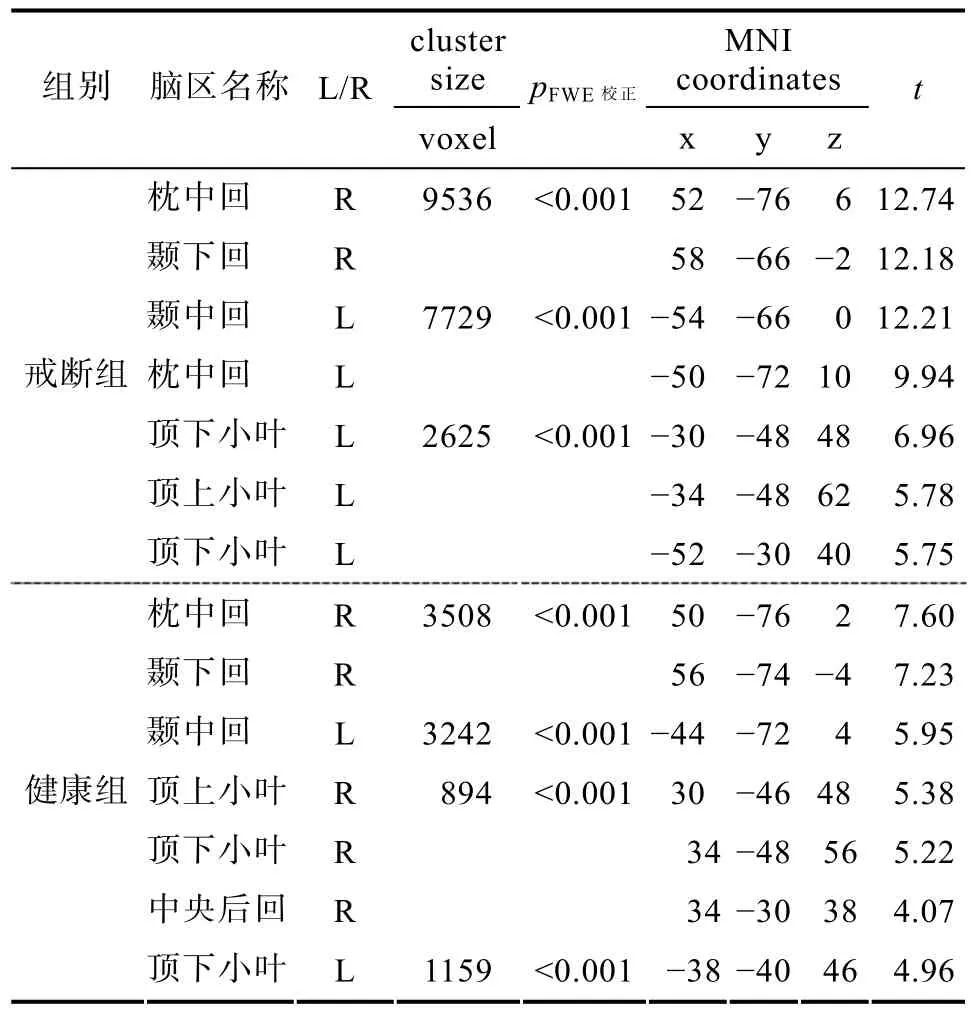
表5 戒断组和健康组对照动作的镜像神经脑区
结合两组被试在对照动作和用药动作线索引发的镜像神经活动情况, 可以发现, AHD和NDP对照动作引发的镜像神经活动没有显示出太大差异;但是他们在用药动作引发的镜像神经活动差异比较大。为了进一步分析药物依赖者在相关线索诱发作用下的镜像神经活动情况, 把AHD在用药动作和对照动作条件下的镜像神经脑区做联合分析。通过联合分析获得AHD用药动作和对照动作共同诱发的镜像神经系统(如表6所示)。
联合分析结果显示, AHD在观看对照动作和用药动作均引发了枕叶−颞叶联合皮层的颞中回, 双侧的顶上小叶和顶下小叶, 以及双侧的中央后回的显著激活; 在p
< 0.05水平下, 减少cluster size的大小, 也会发现右侧额下回的激活。对于NDP来说, 由于用药动作线索引发的脑区较少, 联合分析只有枕叶和颞下皮层有显著激活, 顶叶、颞中回和额下回没有显著激活, 如图2所示。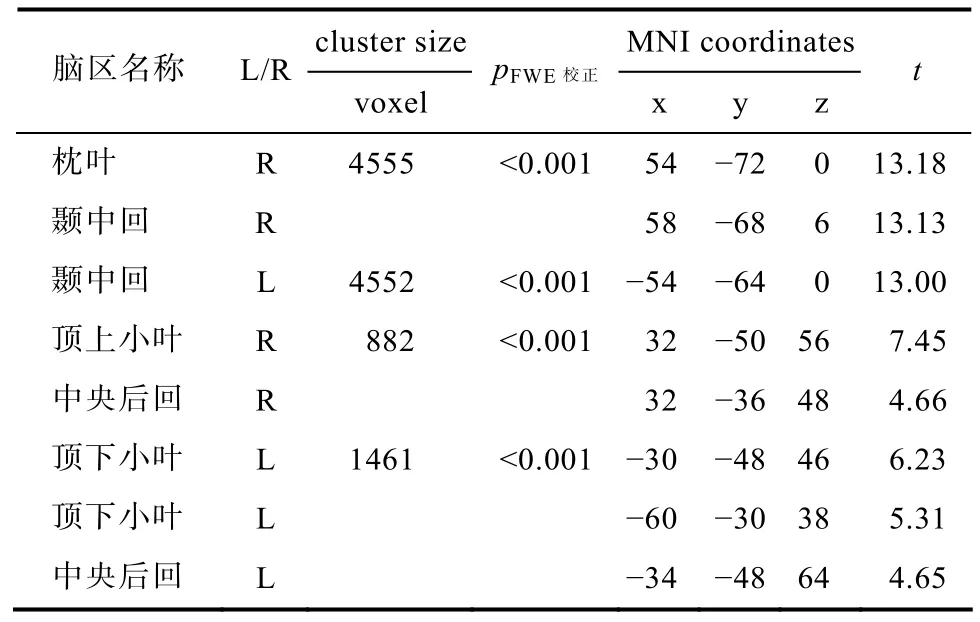
表6 海洛因戒断者用药动作与对照动作镜像神经脑区联合分析
通过联合分析, 比较了戒断后的海洛因依赖者(AHD)和健康人(NDP)用药动作镜像神经活动与对照动作镜像神经活动共同激活的脑区。为了进一步分析两组被试在两种动作条件下镜像神经活动的区别, 选择用药动作与静止的药物两种线索激活脑区比较结果, 对照动作与静止的非药物物品两种线索激活脑区的比较结果, 把这两个比较结果进行T
检验, 即(action vs drug)vs (C-action vs C-drug), 获得AHD和NDP用药动作和对照动作两种镜像神经活动的差异。结果发现, 在p
< 0.001未校正的情况下, AHD用药动作引发的神经活动比一般动作更多激活了枕叶, 以及左侧海马(−22/−28/−6)和中脑(0/−22/−18)和前扣带回的少量体素, 而NDP只有枕叶和右侧额上回出现少量激活。4 讨论
前人在研究相关线索诱发药物依赖者心理渴求的认知神经机制时, 大多选用用药动作的图片(Yalachkov & Naumer, 2011)或者视频(Wagner et al.,2011), 没有对实验材料进行分类, 这在一定程度上阻碍了认识戒断者产生心理渴求的真正原因。本研究选用静止的海洛因图片和吸食海洛因的动作图片作为相关线索, 比较戒断后的海洛因依赖者(AHD)和健康人(NDP)在观看相关线索图片和对照图片时大脑活动的差异, 重点关注AHD和NDP镜像神经系统的活动, 把与药物成瘾主要脑区从中脑奖赏环路发展到全脑, 为更全面地认识心理渴求与复吸提供证据。
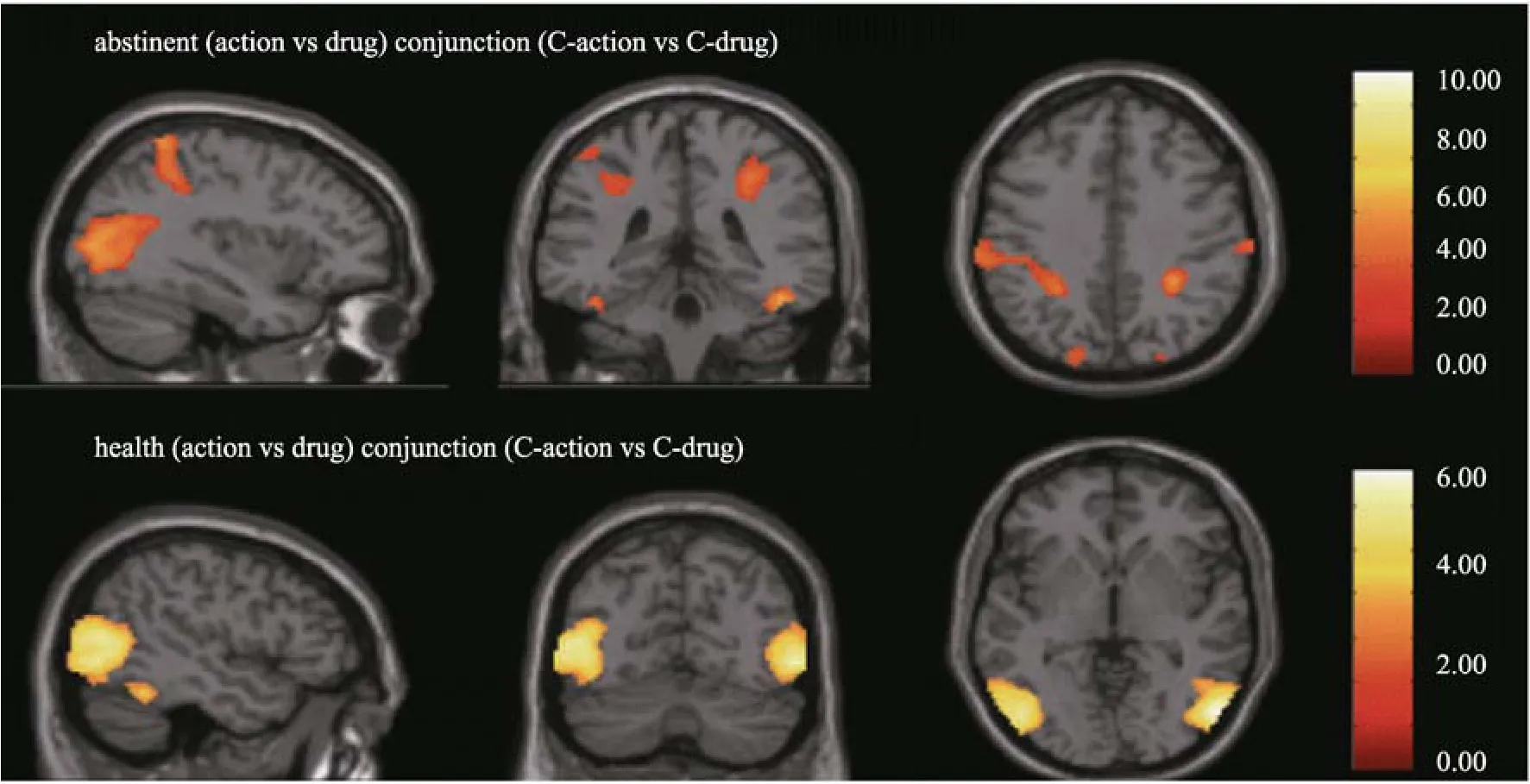
图2 戒断组与健康组用药动作与对照动作镜像神经的联合分析
戒断后的海洛因依赖者(AHD)和健康人(NDP)观看对照动作引发了较为一致的脑区活动, 颞中回、顶上小叶和顶下小叶均出现了显著激活。这一研究发现与van Elk的研究结果是一致的(van Elk,2014)。他选用NDP为研究对象, 发现被试在观看日常生活中的动作线索图片与静止物体的图片相比, 更多激活了双侧颞中回、双侧顶下小叶和双侧的背外侧前运动皮层。这些脑区正是人类的镜像神经脑区。神经科学研究表明, 认知加工可以在环境线索与行为反应之间建立联系。例如, 可操作的物体可以引起动作相关脑区的自动激活, 如前运动皮层和顶叶皮层, 它们反映的是自动化的动作图式和动作经验(Yalachkov & Naumer, 2011)。并且, 观察他人的动作会激活顶叶和额叶的侧部, 这些脑区负责动作的计划、模拟和执行。这表明, 日常生活中常见的动作线索可能引起了AHD和NDP不自觉的动作模拟加工。
在用药动作线索诱发作用下, 戒断后的海洛因依赖者(AHD)双侧颞中回、双侧顶下小叶、左侧顶上小叶和右侧额下回显示出显著活动, 并且与日常生活动作线索激活脑区一致; 健康人(NDP)对应脑区在用药动作线索条件下没有出现显著激活。这一发现与Wagner等人的研究是一致的, 他们发现,抽烟者看到抽烟动作的相关视频材料时, 顶上小叶、顶内沟前端、额下回和前运动皮层等动作表征脑区自动激活。与不抽烟的对照被试相比, 尼古丁依赖者的上述脑区出现了更显著的活动(Wagner et al., 2011)。由此可见, AHD看到用药动作线索, 可能会不自觉地模拟这一熟悉的动作, NDP不会产生类似的动作加工。
AHD虽然摆脱了对药物的生理依赖, 但是他们会对药物线索产生自动化模拟加工。用药动作的自动化加工可能导致心理渴求感, 进而引发药物寻求行为。这一机制可能是精神活性物质和相关线索引起DA释放增加背后不同的地方。本文没能分析出海洛因戒断者在相关线索诱发作用下各个激活脑区之间的动态关系, 未能揭示药物渴求感与用药动作模拟, 以及用药后的欣快感的记忆等神经过程究竟孰先孰后, 这还有待于进一步的探索。
用手拿东西的动作和嘴部动作能引起猴子的镜像神经元活动, 但猴子在观看使用复杂工具的动作时, 镜像神经元没有生物电位变化。研究者认为,因为猴子没有使用过复杂工具的动作经验, 这类动作不能在它们的感觉运动区获得表征(Rizzolatti,Fadiga, Gallese, & Fogassi, 1996)。脑成像研究发现,芭蕾舞蹈演员在看到芭蕾舞蹈动作时, 她们的镜像神经脑区出现了显著激活; 不会跳舞的普通人的上述脑区激活不明显(Calvo-Merino, Glaser, Grèzes,Passingham, & Haggard, 2005)。并且, 不熟悉的舞蹈动作也不会引起舞蹈演员镜像神经脑区的显著活动。例如, 芭蕾舞演员在看爵士时的镜像神经激活水平不及芭蕾舞; 对于不会任何舞蹈的普通人来说, 他们在观看两种舞蹈动作时的镜像神经的激活水平较低, 并没有表现出明显的差异(Cross, Kraemer,Hamilton, Kelley, & Grafton, 2009)。可见, 镜像神经元的活动与个体的动作经验有关。AHD和NDP均熟悉日常生活动作, 因而两组被试在日常生活动作的诱发作用下, 镜像神经系统激活情况是一致的。NDP不熟悉用药动作, 更没有用药经验, 所以他们的镜像神经系统对相关线索不敏感。
相关线索能够诱发戒断后海洛因依赖者(AHD)的心理渴求。研究表明, 中脑奖赏回路和背外侧基底核等是心理渴求的关键脑区(Childress et al., 1999;Volkow, Wang, Fowler, Tomasi, & Telang, 2011)。AHD用药动作与日常生活动作引发的脑区比较结果显示, 他们的左侧海马、中脑和前扣带回也有活动,而健康人(NDP)被试没有发现类似的结果。这表明,相关线索在一定程度上诱发了AHD的药物渴求感。研究结果中, AHD主观报告心理渴求水平较低,且与NDP没有显著差异。一方面, AHD可能在有意隐藏或者压抑渴求感; 另一方面, 相关线索引发的脑激活水平与戒断时间存在显著相关(Lou et al.,2012)。研究中的海洛因依赖者戒断已有十多年(平均戒断时间M
=206.15个月), 这可能是AHD报告低水平的渴求感和心理渴求相关脑区激活不明显的主要原因之一。研究中没有发现心理渴求程度与相关线索引发脑区激活水平之间的显著相关。这可能是由于相关线索图片的诱发心理渴求的能力过低造成的, 以后的研究可以选用在渴求程度上更具有区分度的实验材料。5 结论
相关线索能够引发戒断后的海洛因依赖者和健康人不同的脑神经活动。戒断者对相关线索更加敏感, 相关线索中的动作成分能够引起戒断者镜像神经系统的自动化活动。镜像神经系统对用药动作的心理模拟可能是促发心理渴求的主要原因。进一步的研究需要探索镜像神经系统与纹状体、杏仁核等边缘系统的活动情况, 即明确用药动作模拟与心理渴求这两种心理活动的关系。
致谢
: 华南师范大学博士生王鹏飞对本研究的支持和帮助。Abreu, A. M., Macaluso, E., Azevedo, R. T., Cesari, P., Urgesi,C., & Aglioti, S. M. (2012). Action anticipation beyond the action observation network: A functional magnetic resonance imaging study in expert basketball players.European Journal of Neuroscience, 35
(10), 1646−1654.Calvo-Merino, B., Glaser, D. E., Grèzes, J., Passingham, R. E.,& Haggard, P. (2005). Action observation and acquired motor skills: An FMRI study with expert dancers.Cerebral Cortex, 15
(8), 1243−1249.Calvo-Merino, B., Grèzes, J., Glaser, D. E., Passingham, R. E.,& Haggard, P. (2006). Seeing or doing? Influence of visual and motor familiarity in action observation.Current Biology, 16
(19), 1905−1910.Childress, A. R., Mozley, P. D., McElgin, W., Fitzgerald, J.,Reivich, M., & O’brien, C. P. (1999). Limbic activation during cue-induced cocaine craving.American Journal of Psychiatry, 156
(1), 11−18.Cross, E. S., Kraemer, D. J. M., Hamilton, A. F. d. C., Kelley,W. M., & Grafton, S. T. (2009). Sensitivity of the action observation network to physical and observational learning.Cerebral Cortex, 19
(2), 315−326.Cross, E. S., Liepelt, R., de C. Hamilton, A. F., Parkinson, J.,Ramsey, R., Stadler, W., & Prinz, W. (2012). Robotic movement preferentially engages the action observation network.Human Brain Mapping, 33
(9), 2238−2254.Decety, J., Grèzes, J., Costes, N., Perani, D., Jeannerod, M.,Procyk, E., ... Fazio, F. (1997). Brain activity during observation of actions. Influence of action content and subject's strategy.Brain, 120
(10), 1763−1777.Grafton, S. T., Fadiga, L., Arbib, M. A., & Rizzolatti, G.(1997). Premotor cortex activation during observation and naming of familiar tools.NeuroImage, 6
(4), 231−236.Grant, S., London, E. D., Newlin, D. B., Villemagne, V. L.,Liu, X., Contoreggi, C., … Margolin, A. (1996). Activation of memory circuits during cue-elicited cocaine craving.Proceedings of the Na tional Ac ademy of Sc iences of the United States of America, 93
(21), 12040−12045.Kim, Y.-T., Seo, J.-H., Song, H.-J., Yoo, D.-S., Lee, H. J., Lee,J., ... Chang, Y. M. (2011). Neural correlates related to action observation in expert archers.Behavioural B rain Research, 223
(2), 342−347.Koob, G. F. (1992). Neural mechanisms of drug reinforcement.Annals of the New Y ork Academy of Sciences, 65 4
,171−191.Lou, M. W., Wang, E. L., Shen, Y. X., & Wang, J. P. (2012).Cue-elicited craving in heroin addicts at different abstinent time: An fMRI pilot study.Substance Use & Misuse, 47
(6),631−639.Owesson-White, C. A., Ariansen, J., Stuber, G. D., Cleaveland,N. A., Cheer, J. F., Mark Wightman, R., & Carelli, R. M.(2009). Neural encoding of cocaine-seeking behavior is coincident with phasic dopamine release in the accumbens core and shell.European Journal of Neuroscience, 30
(6),1117−1127.Rizzolatti, G., Fadiga, L., Gallese, V., & Fogassi, L. (1996).Premotor cortex and the recognition of motor actions.Cognitive Brain Research, 3
(2), 131−141.Rizzolatti, G., Fadiga, L., Matelli, M., Bettinardi, V., Paulesu,E., Perani, D., & Fazio, F. (1996). Localization of grasp representations in humans by PET: 1. Observation versus execution.Experimental Brain Research, 111
(2), 246−252.Saunders, B. T., Yager, L. M., & Robinson, T. E. (2013).Cue-evoked cocaine “craving”: Role of dopamine in the accumbens core.The Journal of N euroscience, 3 3
(35),13989−14000.Smolka, M. N., Bühler, M., Klein, S., Zimmermann, U., Mann,K., Heinz, A., & Braus, D. F. (2006). Severity of nicotine dependence modulates cue-induced brain activity in regions involved in motor preparation and imagery.Psychopharmacology,184
(3-4), 577−588.van Elk, M. (2014). The left inferior parietal lobe represents stored hand-postures for object use and action prediction.Frontiers in Psychology, 5
, 333.Volkow, N. D., Wang, G.-J., Fowler, J. S., Tomasi, D., &Telang, F. (2011). Addiction: Beyond dopamine reward circuitry.Proceedings of the National Academy of Sciences of the United States of America, 108
(37), 15037−15042.Wagner, D. D., Dal Cin, S., Sargent, J. D., Kelley, W. M., &Heatherton, T. F. (2011). Spontaneous action representation in smokers when watching movie characters smoke.Journal of Neuroscience, 31
(3), 894−898.Wright, M. J., Bishop, D. T., Jackson, R. C., & Abernethy, B.(2011). Cortical fMRI activation to opponents’ body kinematics in sport-related anticipation: Expert-novice differences with normal and point-light video.Neuroscience L etters,500
(3), 216−221.Yalachkov, Y., & Naumer, M. J. (2011). Involvement of action-related brain regions in nicotine addiction.Journal of Neurophysiology, 106
(1), 1−3.猜你喜欢
High Technology Letters(2021年4期)2022-01-09
浙江大学学报(理学版)(2021年1期)2021-01-26
河北果树(2020年4期)2020-11-26
护理研究(2020年14期)2020-08-11
海峡姐妹(2019年1期)2019-03-23
中学科技(2018年9期)2018-12-19
中成药(2018年9期)2018-10-09
健康管理(2017年3期)2017-04-20
健康必读(2016年8期)2016-09-26
中国医学科学院学报(2012年3期)2012-03-25
