
玉米单倍体诱导系诱导性状的杂种优势分析
2016-01-15 05:26张如养段民孝赵久然刘新香邢锦丰
种子 2016年1期
张如养, 段民孝, 赵久然, 刘新香, 邢锦丰
(北京市农林科学院玉米研究中心/玉米DNA指纹及分子育种北京市重点实验室, 北京100097)
杂种优势是指2个遗传性不同的亲本杂交产生的杂种一代在生长势、抗逆性、产量等方面优于其亲本的现象[1-2]。在玉米常规育种中杂种优势得到广泛的应用,通过将2个遗传背景存在一定差异的自交系进行杂交获得具有强优势的组合应用于生产实践[3-5]。国内外研究结果表明,不同玉米单倍体诱导系诱导率、株高等性状之间也存在杂种优势[6-7],因此如何将诱导性状的杂种优势有效的应用于玉米单倍体育种实践中,不断的提高育种效率是玉米育种者需要深入研究的内容。提高单倍体诱导材料的诱导率可在同等诱导规模下获得更多的单倍体;增加单倍体诱导材料的株高和增大诱导材料的雄穗小花数可以满足DH工程化大规模隔离诱导材料的需求。
玉米单倍体诱导系间杂种优势的利用可为诱导率的进一步提高及其农艺性状的改良提供可能,其主要目的:1)筛选诱导率提高,株高和花粉量等性状表现都优良的诱导系F1;2)获得诱导率与高亲接近,而株高、花粉量得到明显改善的诱导系F1;3)利用诱导率性状提高的F1构建基础群体,为后期新型高频诱导系的选育奠定良好基础[8]。本研究利用8个玉米单倍体诱导系和按照NCⅡ方法进行杂交组配的28个诱导系F1,分析株高、雄穗小花数的杂种优势,并利用6个骨干自交系京724、京92、京 X 005、昌7-2、郑58和农系531为测验种进行诱导率测验,分析不同单倍体诱导系的杂交F1诱导率的杂种优势,以期获得诱导率性状和农艺性状显著改善的诱导系F1,满足大规模隔离种植诱导玉米单倍体的需要,同时也为后期新型高频诱导系的选育提供参考,以期提高玉米单倍体育种效率。
1 材料与方法
1.1 试验材料
试验材料为36个玉米诱导系和6个测验种。其中36个诱导系分别是8个玉米单倍体诱导系,以及28个单倍体诱导系F1。实验中用到的8个玉米单倍体诱导系均为北京市农林科学玉米研究中心从国外引进的不同来源的优良单倍体诱导材料,并经多代自交和测验选育而成,分别是:诱导系JKY 005、JKY 041、JKY 042、JKY 043、JKY 044、JKY 046,都具有控制籽粒顶层和形成胚芽色素的R-nj基因,以及控制不定根、叶鞘和茎秆色素形成的ABPI基因,2个基因同时存在,可使诱导产生的后代具有籽粒和植株双显性标记;而JKY 006、JKY 045只具有控制子粒顶层和形成胚芽色素的R-nj基因。另外28个诱导系F1是由上述8个玉米单倍体诱导系,经过不完全双列杂交而来。6个测验种为玉米自交系,分别是京724、京92、京x 005、郑58、昌72和农系531。
1.2 试验设计
1.2.1 调查诱导材料的株高和雄穗小花数
2012年冬季在海南和2013年夏季在北京分别调查8个诱导系和28个诱导系F1的株高和雄穗小花数性状。每个材料调查5株,3次重复,取均值进行整理分析。
1.2.2 诱导材料与测验种杂交
36个单倍体诱导材料与6个自交系测验种于2012年冬季在北京市农林科学院海南玉米南繁育种基地进行杂交。为了保证诱导系材料与测验种的花期能够相遇,将测验种一次播种,36个单倍体诱导系材料分别在第3天和第5天分2期播种。同时,在开花时期,以36个诱导材料为父本分别对6个测验种进行杂交诱导,平均每个组合杂交15穗。
1.2.3 单倍体数的统计
成熟后收获杂交果穗,脱粒后统计顶部紫色子粒总数,即为杂交子粒数。依据Navajo标记将顶部为紫色、胚部无色的子粒视为拟单倍体子粒。当选的拟单倍体子粒于2013年夏季在小汤山育种基地进行单粒点播种植,精细管理,在苗期依据ABPI紫色植株标记和植株生长势进行进一步的鉴定。淘汰幼苗叶鞘为紫色和生长势强的植株,选择幼苗叶鞘为绿色、生长发育慢、植株较矮、叶片上冲和叶色较浅的单倍体植株,即为单倍体数。2013年夏季在北京市农林科学院玉米研究中心小汤山育种基地进行重复诱导试验,并于2013年冬季在海南进行单倍体鉴定。诱导系的诱导率以对6个测验种的2年试验数据的均值进行整理分析。
1.3 统计项目
单倍体诱导率(%)= 单倍体数/杂交子粒数 ×100%;
中亲优势(%)=(F1-亲平均值)/双亲平均值×100%;
超亲优势(%)=(F1-高亲值)/高亲值×100%;
低亲优势(%)=(F1- 低亲值)/低亲值×100%。
2 结果与分析
2.1 8个诱导系诱导性状的方差分析
由于单倍体诱导率的高低与基础材料遗传背景紧密相关,在测定诱导率时利用了遗传背景存在较大差异的6个自交系为测验种,所得诱导率具有一定的代表性。通过方差分析发现:1)不同诱导系间的诱导率均值、株高和雄穗小花数性状存在极显著差异(表1)。2)不同地点间诱导率均值和雄穗小花数差异不显著。3)不同地点间株高性状存在显著差异,可能原因是海南基地日均气温较高,在一定程度上缩短了个体的生长发育进程,使株高降低。
由表2可知,对于诱导率性状,诱导系JKY 043、JKY 044、JKY 045较高,JKY 006、JKY 042、JKY 046居中,而JKY 005、JKY 041的诱导率较低;对于株高性状,由高到低依次为JKY 005、JKY 046、JKY 042、JKY 044、JKY 045、JKY 041、JKY 043、JKY 006,最高相差30cm;对于雄穗小花数性状,由多至少分别为JKY 045(358.3)>JKY 043(294.0)>JKY 006(271.3)>JKY 046(268.0)>JKY 041(237.3)>JKY 044(225.3)>JKY 042(209.0)>JKY 005(143.7)。8个单倍体诱导系的诱导率、株高和雄穗小花数性状都存在较大差异,这为杂种优势的利用奠定了良好的基础。
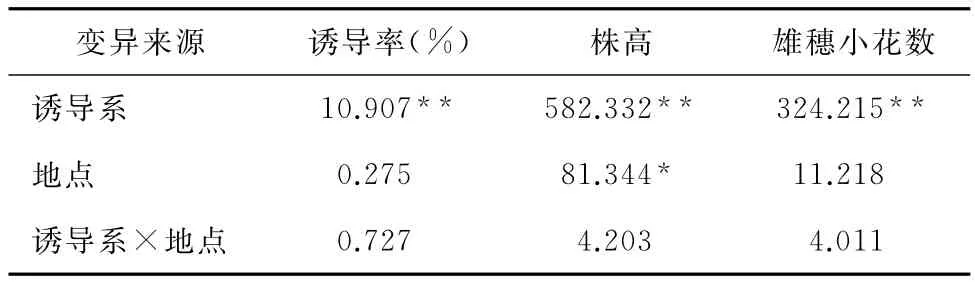
表1 8个诱导系诱导性状均值的方差分析(F值)
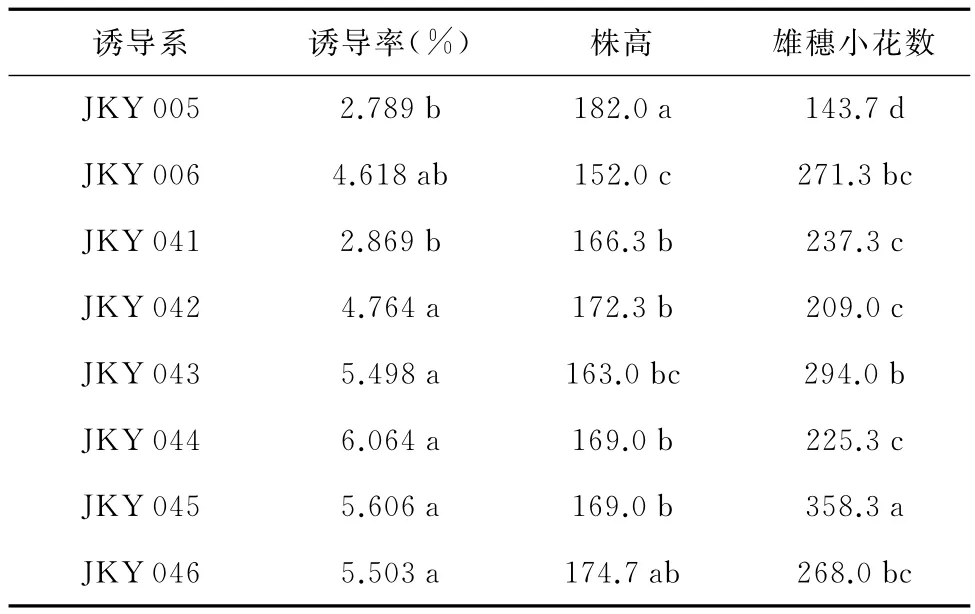
表2 8个诱导系诱导性状均值的多重比较
2.2 株高和雄穗小花数的杂种优势分析
鉴于目前大多数玉米单倍体诱导系都存在株高较低和花粉量偏少的问题,本研究利用不同诱导系相互杂交方法,通过利用株高和雄穗小花数的杂种优势来克服其不足,从而满足单倍体大规模隔离诱导的要求。对8个不同单倍体诱导系相互杂交后的28个杂交种及其父母本的株高和雄穗小花数进行进行杂种优势分析。由图1可知,不同诱导系间杂交F1在株高(左)和雄穗小花数(右)性状上存在着明显的杂种优势。从表3可以看出,对于株高性状,大部分F1在株高性状上都存在一定的杂种优势株,高位于前5位的组合为JKY 005/JKY 042、JKY 044/JKY 046、JKY 005/JKY 044、JKY 044/JKY 045、JKY 005/JKY 043。只有 3个组合(JKY 006/JKY 045、JKY 006/JKY 046、JKY 042/JKY 044)没有表现出超高亲优势。雄穗小花数的多少可以在一定程度上代表雄穗个体花粉量的大小。对于雄穗小花数性状,所有组合的雄穗小花数与其高值亲本相比都有不同程度的提高,位于前5位的组合为JKY 045/JKY 046、JKY 006/JKY 042、JKY 042/JKY 046、JKY 043/JKY 046、JKY 041/JKY 046。因此,利用不同来源的诱导系杂交就能克服单倍体诱导系株高较低和花粉量较少的不足。
2.3 诱导率的杂种优势分析
对玉米单倍体诱导系杂交后所得28个F1诱导率的超高亲优势、中亲优势、低亲优势进行分析结果(表4)表明,15个F1的诱导率与其高亲值相比都有不同程度的提高,最高超高亲值2.296,平均超高亲值0.874 1。另外有8个组合的F1的诱导率没有表现出中亲优势,2个F1组合的诱导率没有表现出低亲优势,整体来看,诱导率主要表现为高亲和中亲优势。同时,表现出超高亲优势组合的2个亲本的诱导率值都普遍较,如JKY 042/JKY 046(6.690),两亲本值分别为4.372和4.678;JKY 042/JKY 044(8.083),两亲本值分别为4.372和5.787;JKY 043/JKY 044(7.473),两亲本值分别为5.842和5.787;因此,通过正确选择双亲,利用其杂种优势来提高杂交组合的诱导率是可能的。诱导率均值前5位的有JKY 042/JKY 044、JKY 043/JKY 044、JKY 044/JKY 045、JKY 042/JKY 046、JKY 042/JKY 043,可在利用杂种优势的基础上,结合自身表现,进一步重点研究利用。
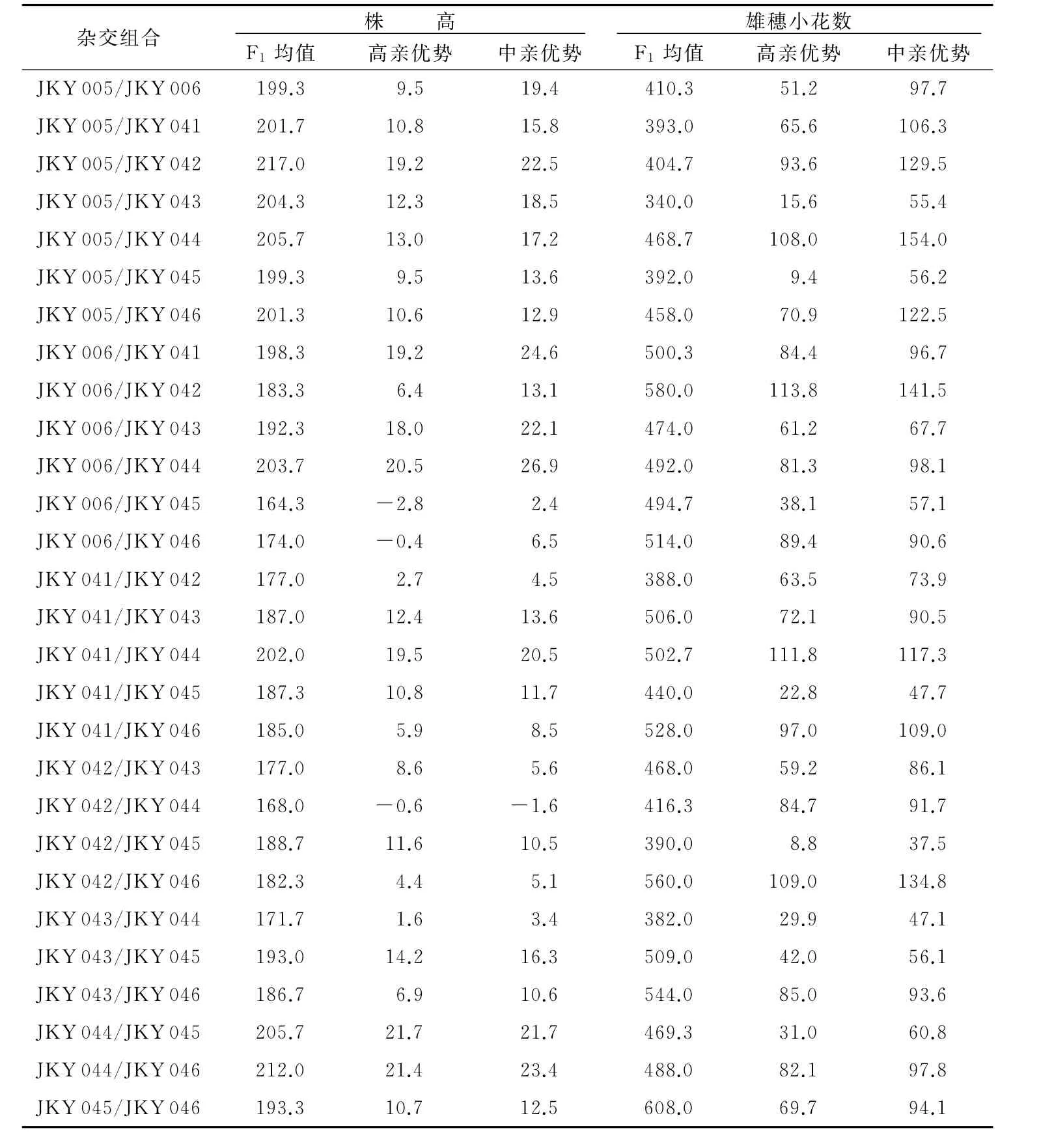
表3 株高和雄穗小花数的杂种优势分析

图1 株高和雄穗小花数的杂种优势

表4 诱导率的杂种优势分析
3 结论与讨论
随着目前玉米DH工程化育种进程的不断推进,对单倍体获得的数量和效率有了更高的要求,而提高玉米单倍体诱导率,是实现大规模获取单倍体的有效途径[9-10]。因此,对于新型玉米单倍体诱导系的选育应以提高诱导率为重点,同时兼备优良的农艺性状。诱导率提高可以大大增加获得玉米单倍体的数量,而农艺性状的改良可以满足规模化隔离诱导单倍体的需求。由本研究结果可知,大部分诱导系F1在株高和雄穗小花数性状上都存在超亲优势,通过利用诱导系间的杂种优势就能较好的克服株高较低和花粉量较少的不足,因此需重点研究诱导率性状的变化。对诱导率的杂种优势分析发现,整体表现以中亲优势为主,同时存在一定的超亲优势。有些表现出超高亲优势组合的两个亲本诱导率均值都较高,如JKY 042/JKY 046、JKY 042/JKY 044、JKY 043/JKY 044。因此,在杂种优势利用时应选择高诱导率的材料进行应用,以期组配和筛选出在诱导率和农艺性状上都存在较大优势的F1应用于DH工程化育种进程。通过杂种优势利用,获得了在诱导率、株高和花粉量性状表现都较为优良的组合JKY 043/JKY 044、JKY 044/JKY 045、JKY 042/JKY 046、JKY 042/JKY 043;而组合JKY 042/JKY 044株高没有超亲优势,但诱导率最高、花粉量较大,具有较高的利用价值,可在后期的育种实践中进一步研究利用。
[1]王懿波,王振华.中国玉米主要种质杂交优势利用模式研究[J].中国农业科学,1997,30(4):16-24.
[2]曾孟潜,吉海莲,李九云,等.玉米杂种优势群及其杂交利用模式概念的形成与发展[J].华北农学报,2007,22(6):30-37.
[3]李竞雄.玉米杂种优势回顾与展望[C]∥刘大钧.植物遗传理论与应用研讨会文集.北京:科学出版社,1990:1-7.
[4]陈彦惠,张世煌,吴连成,等.中国主要玉米改良群体杂种优势组合模式的初步评价[J].华北农学报,2002,17(4):30-36.
[5]魏良明,戴景瑞,张义荣,等.玉米淀粉含量的杂种优势与基因效应分析[J].作物学报,2005,31(7):833-837.
[6]Hartwig H.Geiger.Doubled Haploids[M].IN:Maize Handbook-Volume II:Genetics and Genomics.J.L.Bennetzen and S.Hake(eds),pP 644.Springer Science+Business Media LLC,2009.
[7]徐小炜,董昕,黎亮,等.玉米单倍体诱导系间杂交种评价及其杂种优势分析及其杂交组合选育[J].中国农业大学学报,2014,19(4):1-6.
[8]徐玉波,任保财,张宝石,等.玉米单倍体诱导系的杂种优势利用研究[J].辽宁农业科学,2010(3):40-43.
[9]才卓,徐国良,刘向辉,等.玉米高频率单倍生殖诱导系吉高诱系3号的选育[J].玉米科学,2007,15(1):1-4.
[10]张如养,段民孝,赵久然,等.单倍体技术在玉米种质改良和育种中的应用方向[J].作物杂志,2012(5):4-8.
猜你喜欢
蔬菜(2021年7期)2021-11-27
山西农业科学(2020年8期)2020-08-13
趣味(语文)(2018年7期)2018-06-26
河南农业科学(2018年5期)2018-01-19
考试周刊(2016年88期)2016-11-24
中国学术期刊文摘(2015年8期)2015-10-29
西北农林科技大学学报(自然科学版)(2015年5期)2015-02-21
少年科学(2014年10期)2014-11-14
种子科技(2014年8期)2014-05-18
茶叶通讯(2014年2期)2014-02-27
