
中-晚全新世冷水团对西格陵兰Disko湾的影响*
2016-01-15 11:22李冬玲沙龙滨刘焱光吴燕妮
海洋与湖沼 2016年6期
李冬玲 沙龙滨 刘焱光 蒋 辉 吴燕妮
(1. 宁波大学 地理与空间信息技术系 宁波 315211; 2. 国家海洋局第一海洋研究所 海洋沉积与环境地质国家海洋局重点实验室 青岛 266061; 3. 华东师范大学 地理信息科学教育部重点实验室 上海 200062 )
北极—北大西洋地区由于其海冰、冰川和大陆冰盖覆盖面积广, 反射率高(为开放大洋的 5—10倍),生态系统极其脆弱, 是全球气候变化最为敏感的区域之一。大量研究表明, 大气—海冰—海洋之间的正反馈效应使得该地区在全球气候变化中的作用被放大(Overpeck et al, 1997), 其自身微小的变化就能给全球气候带来较大影响。在全球变暖的背景下, 北极—北大西洋地区的近表层气温变化是全球平均值的两倍(Solomon et al, 2007)。而西格陵兰地区位于格陵兰岛西部, 巴芬湾和戴维斯海峡东侧, 由于特殊的地理位置(北极—北大西洋地区), 其对全球气候变化的响应十分敏感, 因此该区域气候及海洋环境演变逐渐受到研究者的广泛关注。
在西格陵兰北部的巴芬湾地区, Levac等(2001)根据海洋沉积物中孢粉及沟鞭藻孢囊重建了该区域全新世以来的海水表层温度变化, 发现早中全新世时海水表层温度较高, 其中最高温比现代高出约 3°C;晚全新世以来海洋环境转冷, 海冰覆盖面积扩大。格陵兰西南部峡湾也是研究者广泛关注的区域。其中Lassen等(2004)和Roncaglia等(2004)分别对格陵兰西南部 Igaliku峡湾沉积物中的底栖有孔虫和沟鞭藻孢囊做了研究, 其结果记录了明显的中世纪暖期和小冰期。在Igaliku峡湾更北的Ameralik峡湾, 其沉积物粒度、元素地球化学、有孔虫及硅藻分析的结果显示该峡湾在4400—3200a BP期间经历了全新世大暖期的末段, 之后气候逐渐转冷且波动变化, 且Ameralik峡湾的这种变化与 Igaliku峡湾及北大西洋地区的气候变化有很好的一致性(Møller et al, 2006;Seidenkrantz et al, 2007; Ren et al, 2009)。
此外, 位于西格陵兰中部的Disko湾由于湾口较大, 能与湾外水团进行顺利交换, 且有大量格陵兰冰川融水经峡湾流入湾内, 因此, 了解 Disko湾水团性质是十分必要的。通过对Disko湾内Jakobshavn峡湾两个钻孔的有孔虫和粒度进行分析, Lloyd等(2005)发现9200—7800a BP西格陵兰洋流的增强与冰盖的后退同步发生; 7800a BP之后西格陵兰洋流开始影响Disko湾东部。Moros等(2006)和Lloyd等(2007)对该地区的相关研究同样揭示了Disko湾地区暖期与西格陵兰洋流中北大西洋暖流的增强密切相关。Lloyd(2006b)分析了 Disko湾表层沉积物中底栖有孔虫的分布, 认为有孔虫属种的分布特征与西格陵兰洋流携带的冷、暖水团强度有关。Seidenkrantz等(2008)通过Disko湾内沟鞭藻的含量变化重建了3000年以来大西洋暖水团的变化情况。以上研究均表明,西格陵兰洋流中冷、暖水团的此消彼长对Disko湾的气候及海洋环境变化影响显著。
然而, 这些研究都集中在 Disko湾湾口及中部,对 Disko湾北部 Vaigat海峡处的相关研究则较少。Andresen等(2011)通过对Vaigat海峡沉积物的岩性、鞭毛藻及有孔虫的分析, 发现中全新世以来西格陵兰洋流对该区域海洋环境演化具有十分重要的影响。此外, Sha等(2014)重建了该区域中全新世以来的海冰密集度变化, 同样发现西格陵兰洋流冷、暖水团的变化是除太阳辐射强度变化之外另一个重要的海冰密集度变化驱动因素。但以上研究只是通过各指标属种的分布简单描述了西格陵兰洋流的变化, 缺乏对洋流强度的系统研究。
硅藻化石种类多、数量大、分布广, 对温度、盐度及水深等环境变化反应敏感, 且易于保存, 能反映水团特性的变化, 是良好的环境指示器, 因而被广泛应用于古气候、古海洋环境演变研究中(Hendey, 1964;Hasle et al, 1997)。近年来, 研究者通过对北极―北大西洋地区化石硅藻组合的系统研究, 为了解北极高纬地区古气候及古海洋环境演变提供了详细资料(Koç Karpuz et al, 1990, 1993; Williams, 1990; Cremer,1999; Jiang et al, 2001, 2002, 2005, 2015)。本文以海洋沉积硅藻为研究对象, 通过对西格陵兰Disko湾北部Vaigat海峡内沉积钻孔中特征性硅藻属种的百分含量变化进行研究, 并运用主成分分析方法, 试图对研究区中全新世以来西格陵兰洋流中冷、暖水团的强度变化进行系统的研究, 从而为进一步系统论证北极—北大西洋地区全新世海冰变化的影响机制提供佐证。
1 区域概况
Disko 湾位于 68°30′—69°15′N, 50°—54°W, 是西格陵兰地区较大的海湾之一, 面积约为 4×104km2(图1)。其纬度位置决定了该区域温度季节变化较大, 且太阳光照时间也存在着明显的季节变化(11月下旬至次年1 月中旬处于极夜)。Disko湾海底地形复杂, 海槽、海槛较多。西南部湾口处的Egedesminde Dyb海槽水深最大约1000m; 湾中部平均水深在200—400m;湾北部的Vaigat海峡处最大水深约600m (Andresen et al, 2011)。
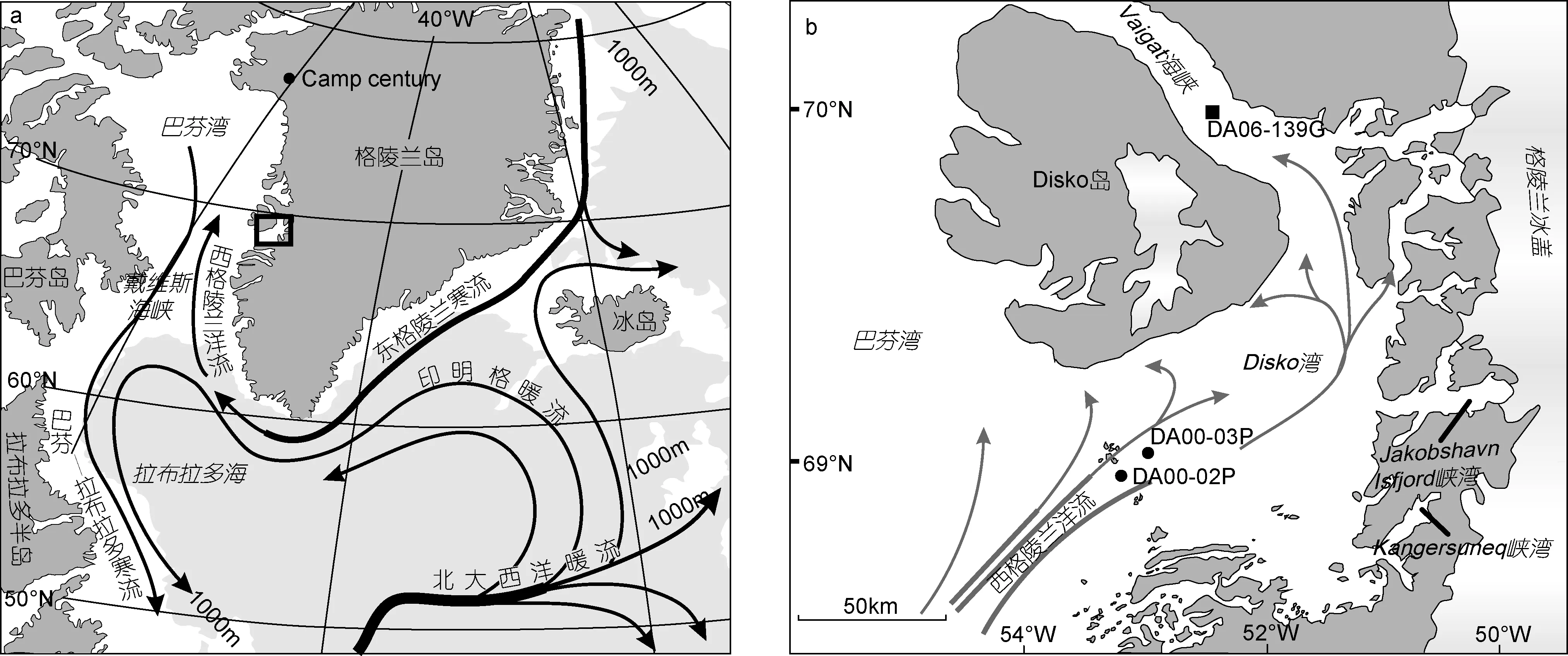
图1 北大西洋表层海洋环流系统(a)及DA06-139G孔的站位图(b)Fig.1 Oceanographic surface current system in the North Atlantic (a) and location of core DA06-139G (b)
Disko湾北部的Vaigat海峡位于Disko岛和格陵兰大陆之间, 长约130km, 宽约20—25km (Andresen et al,2011)。Vaigat海峡不仅是西格陵兰洋流流出的必经之地,也是湾内浮冰流入巴芬湾的主要出口。而 Jakobshavn峡湾是Disko湾最重要的冰川出口。现代Jakobshavn港位于 Jakobshavn峡湾的最前端, 格陵兰冰盖近 7%的冰融水排入港内(Bindschadler, 1984)。西格陵兰中部每年冰川崩解后有约1/3经Jakobshavn峡湾流入Disko湾。Disko湾海底的几个主要海槽很可能是冰期Jakobshavn Isbrae冰川向西前进侵蚀而成(Long et al, 2003)。
Disko湾复杂的地形也使得湾内水文特性与西格陵兰沿岸其他区域明显不同(Jensen, 2003)。Disko湾的现代水文特征主要受西格陵兰流(West Greenland Current, WGC)的影响。西格陵兰洋流的上层水团主要来自东格陵兰寒流(East Greenland Current, EGC,全年水温都低于0°C, 平均为-1.8°C, 盐度为32—33)的极地水和西南格陵兰的冰川融水(Jennings et al,1996, 2002), 下层水团为来自印明格暖流 (Irminger Current, IC, 温度约 4—6°C, 盐度约 35)的大西洋暖水团, 深度约为 200—500m (Eiríksson et al, 2000;Cuny et al, 2002)。此外, 由Jakobshavn峡湾流出的冰融水和冰山也对 Disko湾水文特征产生重要的影响,冰融水及冰山在洋流的作用下继续北上经 Vaigat海峡注入巴芬湾(Andresen et al, 2011)。每年夏季, 格陵兰冰盖及海冰大量融化, 造成湾内水团分层明显; 而9月份开始冰盖海冰融化减弱, 气旋活动增强, 导致水团交换不断加强, 特别是冬季时湾内水团盐度较为均一(Andresen et al, 2011)。大量研究表明, 西格陵兰洋流中冷、暖水团的强度变化决定了Disko湾特别是 Vaigat海峡处的海洋环境演变(Lloyd, 2006b;Andresen et al, 2011)。
Disko湾现代海冰分布具有明显的季节性, 每年1月中旬至4月中旬被海冰覆盖, 平均厚度约为0.7m,其余时间以冰山分布为主(Andresen et al, 2011)。Disko湾是格陵兰西部最为典型的冰山分布区域之一。湾内冰山一部分在西格陵兰洋流的携带下经Vaigat海峡流入巴芬湾, 然后继续北上; 另一部分经戴维斯海峡进入Labrador海, 然后在Baffin-Labrador流携带下南下(Tang et al, 2004)。
2 材料和方法
2.1 硅藻样品处理与属种统计
DA06-139G(70°05.486′N, 52°53.585′W)为 2006年丹麦Dana航次于Vaigat海峡所钻取的柱状岩心(图1)(Dalhoff et al, 2007)。该孔位于南Vaigat海峡的中部, 水深384m, 柱长4.46m。
本文对DA06-139G孔共采集了90个样品, 采样间隔为 5cm。硅藻样品按 Håkansson(1990)的方法处理, 先加入 10% HCl去除钙质, 然后用蒸馏水洗净,再加入30% H2O2后, 在恒温 60°C下水浴加热1—2h直至沉积物呈灰白色, 以去除沉积物中的有机质, 洗净后用 Naphrax胶制片, 然后在 1000倍徕卡油镜下进行属种鉴定, 每个样品至少计 300个硅藻壳面(Chaetoceros休眠孢子除外)。
2.2 年代框架
DA06-139G孔测年由丹麦Aarhus大学加速器质谱(AMS)14C测试中心完成, 共测得10个AMS14C数据, 测年材料分别为海洋软体动物壳体、海洋植物碎屑以及底栖有孔虫(表1, 图2)。所有的14C年龄均利用OxCal 4.1软件(Ramsey, 2008)的Marine09数据库(Reimer et al, 2009)校正为日历年龄(a BP), 其中碳储库年龄R为540±30年(McNeely et al, 2006; Lloyd et al, 2011)。各测年点之间采用线性插值和外推以计算各个深度的年龄。
2.3 主成分分析
主成分分析(principal component analysis, PCA)是一种常见的统计分析方法, 其原理主要是通过特定的降维手段将众多变量转化为几个主成分(综合变量), 这些主成分能够准确反映出原始变量的绝大部分信息。
本文采用CANOCO软件对DA06-139G孔硅藻属种结果进行主成分分析, 研究 5000年以来钻孔所在海域不同硅藻属种间的相互关系, 并结合硅藻属种的生态环境意义, 确定主成分轴所指示的环境指标, 进而对环境指标进行定量/半定量探讨。主成分属种载荷图(图 4)展示了不同属种间的相互关系,其中各矢量轴表示主要的硅藻属种, 横、纵坐标轴(轴一和轴二)表示最主要的主成分。矢量轴的长短与其代表的硅藻属种的丰富程度呈正比。各矢量间及其与坐标轴之间的夹角代表了属种之间以及属种与各主成分之间的相关性, 夹角为 90°时, 表明两者间无明显相关性, 夹角小于或大于 90°则分别表示两者呈正相关或负相关关系, 而硅藻属种与环境变量之间的定量关系则由其在主成分轴上的投影得到。

表1 DA06-139G孔AMS14C测年资料 (Andresen etal, 2011; Sha etal, 2014)Tab.1 AMS 14C age determinations on core DA06-139G

图2 DA06-139G孔深度-年代模式图Fig.2 Age-depth model of gravity Core DA06-139G
3 主要硅藻属种生态及古环境意义
Disko湾 DA06-139G孔共鉴定硅藻样品 90个,主要硅藻属种有Thalassiosiraantarcticavar.borealisresting spores,T.antarcticavegetative cells,Fragilariopsiscylindrus,Fossulaarctica,Thalassiosira bulbosa,Fragilariopsisoceanica,Thalassiosira nordenskioeldii,Bacterosirabathyomphala,Detonula confervaceaeresting spores,Thalassiosirahyalina,Thalassionemanitzschioides及Thalassiosiraoestrupii。其中, 广布种T.antarcticavar.borealisresting spores以及T.antarcticavegetative cells占绝对优势, 其含量多在40%以上 (Shaetal, 2014)。
DA06-139G孔所发现的硅藻海冰种主要包括F.cylindrus,F.arctica,T.bulbosa和D.confervaceaeresting spores。
F.cylindrus是典型的极地海冰硅藻种(Medlinet al, 1990; Hasleetal, 1997)。该种广泛分布于南北两极,既可生活在冰下甚至冰内, 同时也可以存在于冰缘区域(Hasleetal, 1997)。在Nordic海海域该种是海冰型硅藻组合的主要组成成分, 主要分布于格陵兰海西部(Koç Karpuzetal, 1990); 在冰岛附近海域该种是海冰硅藻群落的最主要组成成分, 分布在受东格陵兰寒流影响强烈的海域(Jiangetal, 2001)。Ren等(2014)通过对北太平洋表层沉积物硅藻的现代分布特征进行研究, 发现F.cylindrus分布在冬季海冰覆盖区域内, 且该种在海水表层温度为 5—8°C的海域分布最广, 而在海水表层温度超过 12.5°C的海域没有发现该种。
D.confervaceaeresting spores在北极、北冰洋Laptev海等是常见的海冰种(Bauchetal, 2000;Polyakova, 2001)。Syvertsen等(1984)研究发现T.bulbosa能在海冰或海冰边缘生存, 因此也被认为是海冰硅藻的重要组成。F.arctica是常见的极地半咸水种, 主要分布在海冰周围(von Quillfeldt, 1996), 在Disko湾(Jensen, 2003; Krawczyketal, 2010)和 Nares海峡(Knudsenetal, 2008)沉积物中均有发现。因此,以上四种硅藻被认为是北大西洋西北部海域最典型的海冰硅藻种属, 其含量越高, 表明海冰覆盖范围越广(Syvertsenetal, 1984; Jensen, 2003; Krawczyketal,2010; Shaetal, 2014)。
DA06-139G孔所发现的极地冷水种主要包括F.oceanica,T.nordenskioeldii,B.bathyomphala以及T.hyalina。F.oceanica是一种常见的极地浮游硅藻, 与海冰关系密切(Hasleetal, 1997), 因此, 与F.cylindrus一样, 该种之前常被作为海冰指示种( Koç Karpuz et al, 1990; Jiang et al, 2001)。然而, 最近一些研究发现两者的生活环境存在较大差异(Jensen et al,2004; Ren et al, 2009; Sha et al, 2014)。Heimdal (1989)用 F. oceanica指示西格陵兰洋流冷暖混合水团。Jensen等( 2004)认为 F. oceanica的生活环境较 F.cylindrus偏暖。Disko湾表层硅藻属种与环境变量的CCA分析(Canonical correspondence analysis)也显示F. oceanica与海冰密集度无明显相关性(Sha et al,2014), 因此本文认为该种可能反映了极地冷水团对研究区的影响。
B. bathyomphala主要分布在北半球冷水海域(Hasle et al, 1997)。已有研究发现在挪威海的西南海域, 该种与海冰有着密切的联系, 它也是Nordic海海冰硅藻组合的重要组成部分(Koç Karpuz et al, 1990)。但冰岛附近海域沉积硅藻的研究结果(Jiang et al,2001)表明该种是极地冷水硅藻组合的重要组成, 其适宜的水温条件比海冰种 F. cylindrus略高, 而与极地冷水硅藻的生存环境更为接近, 表明该种的分布主要受东格陵兰寒流携带的极地冷水团影响。
T. hyalina常出现于北大西洋极地海域(Hendey,1964), 前人研究显示T. hyalina为极地冷水种, 与春季海冰有密切关系(Kanaya et al, 1966)。Hasle等(1997)认为该种主要分布于北半球冷水到温水海域。Jiang等(2001)通过对冰岛附近海域表层沉积硅藻的研究,发现该种主要分布于冰岛北部和西部受冷水团影响的陆架海域。
T. nordenskioeldii是一种分布于北半球极地地区的冷水种, 广泛分布于北大西洋、北海、及北冰洋等沿岸海域(Hendey, 1964; Hasle et al, 1997)。该种春季时大量繁殖, 含量迅速增加, 有时也出现在浮冰边缘(De Sève and Dunbar, 1990; Cremer, 1999)。Karentz 等(1984)通过研究美国东北部 Narragansett湾浮游硅藻1959—1980年的温度及季节分布特征, 发现 T.nordenskioeldii是冬季浮游种, 其生长的最适宜水温在(2.8±1.8)°C。T. antarctica var. borealis resting spores作为主要的北方硅藻种, 广泛分布于北半球冷水到温水海域(Hasle et al, 1997; von Quillfeldt, 2000)。通过对西格陵兰海域浮游植物群落的研究发现, T.antarctica var. borealis休眠孢子在春、夏季无海冰覆盖的海域大量繁殖生长, 且主要分布在西格陵兰外海海域(Krawczyk et al, 2014)。
DA06-139G孔所发现的暖水种主要包括 T.nitzschioides和T. oestrupii。T. nitzschioides作为广布种在北海和英吉利海峡中都有被发现(Hendey, 1964),但从未在北极地区发现过该种(Hasle et al, 1997)。在Nordic海(Koç Karpuz et al, 1990)、Skagerrak 海(Jiang,1996)以及冰岛南部和西部海域(Jiang et al, 2001), T.nitzschioides大量出现, 反映出该种与大西洋暖水团(平均盐度>34.9, 平均海水表层温度>3°C)联系紧密,其百分含量随着印明格暖流的减弱而逐渐降低。对冰岛附近海域表层沉积物硅藻分布的研究发现, T.nitzschioides含量在冰岛南部最高, 在冰岛西部和北部逐渐减少(Jiang et al, 2001)。此外, 在Disko湾和格陵兰西部海域, 其百分含量仅占 1%左右(Sha et al,2014)。因此, T. nitzschioides含量的波动可用来反映大西洋暖水团的强弱变化。
T. oestrupii是一种暖水至温水硅藻种(Hasle et al,1997), 广泛分布于低纬度暖水海域(Jiang et al,2006)。在 Nordic海海域, 该种是大西洋硅藻组合的主要组成属种(Koç Karpuz et al, 1990), 主要分布在受北大西洋暖流影响的海域。Jiang等(2001)通过研究冰岛周围海域表层沉积硅藻的分布特征, 发现 T.oestrupii在冰岛南部和西南部海域含量最为丰富, 是该区域暖水硅藻群落的主要组成成分, 主要反映印明格暖流的强弱变化。
4 结果
4.1 特征性硅藻属种分布
根据硅藻属种的生态环境意义, 本文将DA06-139G孔除广布种之外的海冰种硅藻、极地冷水种硅藻和暖水种硅藻的百分含量分别进行求和,如图3所示。其中, 海冰种硅藻(F. cylindrus, F. arctica,T. bulbosa和D. confervaceae resting spores)、极地冷水种硅藻(F. oceanica, T. nordenskioeldii, B.bathyomphala以及T. hyaline)百分含量的变化主要反映海冰及东格陵兰寒流携带的极地冷水团对研究区海洋环境的影响; 而暖水种硅藻(T. nitzschioides和T.oestrupii)的百分含量变化主要反映印明格暖流携带的北大西洋暖水团对研究区的影响。
5000—3800a BP期间, DA06-139G孔中海冰种硅藻百分含量多低于平均值 14.0%(图 3a), 极地冷水种硅藻含量也低于平均值25.4%(图3b), 而暖水硅藻的百分含量在该时期多高于平均值(图 3c), 表明 Disko湾海域受极地冷水团的影响较弱, 而受北大西洋暖水团的影响相对较强, 海洋环境较温暖。

图3 Disko湾DA06-139G孔5000年以来海冰种(a)、极地冷水种(b)和暖水种硅藻(c)的含量(%)变化(虚线为平均值)Fig.3 Changes in abundance of sea-ice species, Arctic species and warm water species over the past 5000 years in Disko Bugt(the dashed lines are the average values of sea-ice, Arctic and warm-water diatom species, respectively)
DA06-139G孔中海冰种的百分含量在 3800—3400a BP期间显著增加, 随后呈现平稳的波动变化(图3a)。在2700—2300a BP期间, 其含量高于平均值14.0%, 而极地冷水种的含量在此期间也呈逐渐上升的趋势, 并达到峰值(图 3b), 表明该时期 Disko湾受极地冷水团的影响增强, 海洋环境较为寒冷。相比较而言, 暖水种T. nitzschioides和T. oestrupii的含量在3050—2850a BP期间存在一个显著的低值(图3c), 也证明Disko湾该时期海水温度较低。
DA06-139G孔海冰种和极地冷水种含量在2000—1500a BP期间显著减少(图3a, b), 而暖水硅藻的含量在此期间却出现增加的趋势(图3c), 均表明该时期东格陵兰寒流携带的极地冷水团势力减弱,Disko湾受北大西洋暖水团的影响增强, 海洋环境较为温暖。
1500a BP以后, 海冰种和极地冷水种的含量开始波动上升, 特别是在 650—0a BP期间, 其含量均达到峰值(图3a, b)。暖水硅藻含量自1500a BP以后波动较大, 且从 650 a BP开始显著下降, 特别是在650—250a BP期间暖水硅藻几乎没有出现(图3c), 以上都反映了1500a BP以来, Disko湾主要受东格陵兰寒流携带的极地冷水团的影响, 海洋环境呈不断变冷的趋势。
4.2 主成分分析结果
为进一步确定不同硅藻属种的环境指示意义,本文对DA06-139G孔的硅藻属种进行了主成分分析,各属种在主成分轴上的得分如图4所示。主成分分析结果显示, 第一和第二主成分对所有属种变量总方差的累计贡献率达到了 89.2%(主成分一: 79.7%; 主成分二: 9.5%), 而其余主成分贡献率均较小。这表明第一和第二主成分可以解释包含在所有属种数据中的大部分信息, 因而可以通过研究第一和第二主成分所代表的环境意义, 来获取研究区域主要的环境变化信息。
通过主成分属种载荷图可以看出, 硅藻种 F.cylindrus、D. confervaceae resting spores、T. bulbosa、T. hyalina、B. bathyomphala、T. nordenskioeldii和 F.oceanica等分布于载荷图的右侧, 且与主成分轴一正方向的夹角小于90°(图4), 这表明以上硅藻属种与主成分轴一呈明显的正相关关系。另一方面, 硅藻种T.antarctica var. borealis resting spores、T. nitzschioides、T. oestrupii和Rhizosolenia borealis等分布于载荷图的左侧, 与主成分轴一负方向的夹角小于 90°(图 4),表明上述硅藻属种与主成分轴一呈负相关关系。结合主要硅藻属种的生态环境意义不难发现, 分布于主成分轴一正方向(即在轴一得分为正)的硅藻属种主要以海冰种硅藻和极地冷水种硅藻为主, 而轴一负方向上(即在轴一得分为负)的硅藻属种以暖水种硅藻和广布种为主。Disko湾Vaigat海峡处海洋环境的演变主要受携带极地冷水团的东格陵兰寒流和北大西洋暖流的分支印明格暖流共同影响 (Lloyd, 2006b;Andresen et al, 2011)。当海冰种硅藻和极地冷水种硅藻得分较高时说明此时海洋环境受极地冷水团影响较强; 反之当极地冷水团势力减弱, 北大西洋暖水团增强时, 暖水硅藻得分较高。综上所述, 主成分轴一可以用来指示西格陵兰地区极地冷水团的强度变化。
此外, 由于主成分轴二对所有硅藻属种变量总方差的贡献较小(9.5%), 且主要硅藻属种与主成分轴二的相互关系不明确, 无法确定主成分轴二所指示的环境意义, 因此本文将不对主成分轴二展开讨论。
根据硅藻各属种在主成分一上的得分, 计算DA06-139G孔沉积物样品中各属种含量的加权平均值, 得到各样品在主成分一上的得分(图 5a), 可用于评估5000年以来西格陵兰地区极地冷水团的强度变化, 样品得分高指示了西格陵兰地区极地冷水团势力较强, Disko湾海水温度相对较低, 海冰覆盖度高;相反, 样品得分低则表明极地冷水团强度减弱, 北大西洋暖水团的影响可能增强, 湾内海冰融化。

图4 DA06-139G孔主成分分析属种载荷图Fig.4 Principal component analysis (PCA) of taxa scatter diagram of axis 1 against axis 2
由图5a可以看出, 过去5000年以来DA06-139G孔各样品在主成分一的得分呈现阶梯式增加的趋势,表明中晚全新世以来Disko湾海域极地冷水团的强度阶段性增强。5000—3800a BP期间, 样品在主成分一的得分较低; 之后样品得分缓慢增加, 并在3800—2750a BP期间保持较为平稳的变化; 2750a BP之后样品得分再次增加, 该高值持续到 2000a BP左右; 样品在主成分轴一的得分值在 1600a BP之后开始显著升高, 特别是距今1500—1200a BP和650a BP之后, 得分更是达到峰值。
5 讨论
5.1 5000—3800a BP气候温暖期
5000—3800a BP期间, DA06-139G孔各样品在主成分一的得分较低(图5a), 反映该时期东格陵兰寒流所携带的极地冷水团的强度较弱, 受此影响Vaigat海峡海冰种和极地冷水种硅藻含量较低而暖水种硅藻含量相对较高(图3a, b)。该钻孔的有孔虫记录也表明在5000—4500a BP期间进入Vaigat海峡的极地冷水团强度较弱, 而大西洋水团在此期间对Vaigat海峡的影响较强(图5b)(Andresen et al, 2011)。
全新世大暖期(Holocene thermal maximum, HTM)在格陵兰地区持续的时间因区域而异。其中在格陵兰南部地区HTM持续时间为7000—3000a BP (Kaplan et al, 2002); 在东格陵兰陆架 HTM 结束于 4700a BP(Jennings et al, 2002; Kaufman et al, 2004); 而在西格陵兰地区HTM结束于3500—3100a BP(Møller et al,2006; Moros et al, 2006)。本文认为5000—3800a BP期间Disko湾相对温暖的海洋环境对应于格陵兰地区的HTM末期(Dahl-Jensen et al, 1998; Kaplan et al,2002; Kaufman et al, 2004)。
Jiang等( 2002)通过对冰岛北部的沉积硅藻研究,发现在4600—3600a BP期间印明格暖流向北的分支增强, 同时也伴随着其向南分支(即影响西格陵兰沿岸的暖水团)的逐渐增强(Moros et al, 2006; Lloyd et al, 2007)。来自格陵兰东部、冰岛南部及西南部的研究也表明, 印明格暖流的加强, 会导致西格陵兰洋流加强且变暖(Jennings et al, 2002; Knudsen et al, 2008;Ólafsdóttir et al, 2010; Jennings et al, 2011)。
来自西南格陵兰峡湾的相关研究表明, 在 HTM末期, 西南格陵兰地区受北大西洋暖水团的影响, 海水温度较高, 而此时东格陵兰寒流携带的极地冷水团则较弱(Seidenkrantz et al, 2007; Ren et al, 2009)。北大西洋暖水团的增强, 抑制了极地冷水团的北上,进而影响Disko湾的水文和海洋环境变化。Disko湾南部两个沉积钻孔的硅藻结果显示, 4800—3500a BP期间海冰种硅藻含量明显减少(Moros et al, 2006), 且重建的 Disko湾海冰密集度变化在 5000—3800a BP期间值较低(Sha et al, 2014, 图5c), 以上均表明HTM末期Disko湾地区极地冷水团强度较弱, 海冰覆盖面积小, 海洋环境偏暖。
此外, 在西格陵兰北部的巴芬湾, 海水表层温度的重建结果显示7300—4000a BP期间海水表层温度比现在高出约 3°C且一年中有 4—5个月的无冰期(Levac et al, 2001); 有孔虫记录也表明该时期西格陵兰洋流对巴芬湾的影响有所增强(Knudsen et al,2008)。
Camp Century冰芯与格陵兰西部沿岸毗邻, 因此Camp Century冰芯δ18O数据记录的表层空气温度变化与西格陵兰的海水温度变化密切相关。Camp Century冰芯记录显示5500—3500a BP期间, 空气温度相对较高(图5d)。Jakobshavn Isbræ附近的湖泊记录也反映了当时较为温暖的陆地条件(Axford et al,2013)。Disko湾南部 Kangerlussuaq地区的高湖平面记录也与当时较为温暖的的条件有关(Aebly et al,2009)。Briner等(2013)运用贝壳年代学重建了冰雪覆盖范围, 认为 5000—3000a BP期间冰雪覆盖范围最小。根据转换函数定量重建的海冰密集度变化, 同样显示该时期 Disko湾海冰覆盖面积较小(Sha et al,2014, 图 5c), 该结果与加拿大极地群岛的生物标志物IP25记录(Vare et al, 2009; Belt et al, 2010)以及冰岛北部硅藻重建结果(Justwan et al, 2008)均相近。这表明极地冷水团的减弱(或印明格暖流的增强)是影响HTM末期海冰减少的重要因素。
5.2 3800—2000 a BP气候较冷期
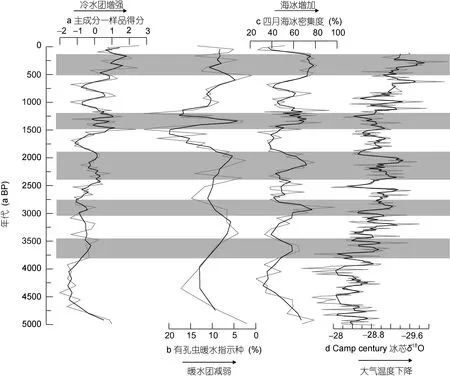
图5 DA06-139G孔各样品在主成分一上的得分(a)、该孔有孔虫北大西洋暖水指示种的百分含量(Andresen et al, 2011)(b)、以及重建的四月海冰密集度(Sha et al, 2014)(c)与Camp Century冰芯的δ18O数据(Vinther et al, 2009)(d)的比较Fig.5 Comparison of the present diatom record (sample scores on axis 1) with the Atlantic foraminiferal assemblage and reconstructed April sea-ice concentration of core DA06-139G (Andresen et al, 2011; Sha et al, 2014), and the δ18O data from the Camp Century ice core (Vinther et al, 2009)
3800a BP之后, 主成分轴一的得分开始逐渐升高, 表明西格陵兰洋流中极地冷水团对Disko湾的影响缓慢增强, 而北大西洋暖水团的影响减弱, 对应研究区海水温度下降, 尤其是在 3700—3500a BP,3000—2800a BP和2500—2100a BP期间(图5a)。根据 DA06-139G孔的有孔虫研究结果, 大西洋暖水指示种在 3800—2000a BP期间含量减少(图 5b), 反映当时大西洋暖水团对研究区的影响减弱(Andresen et al, 2011), 这与本文硅藻结果有较好的一致性, 对应于欧洲典型的气候寒冷期“新冰期”(Williams et al,1985; Dyke et al, 1996; Kaplan et al, 2002)。Camp Century冰芯也记录了 3500—2600a BP这一冷事件(Vinther et al, 2009; 图 5d)。
新冰期是北半球全新世大暖期之后比较明显的寒冷期, 一般开始于3900a BP左右(Matthews, 2007)。该冷事件在格陵兰岛(冰盖扩张)(Kelly, 1980)、Labrador海东部、巴芬湾、格陵兰东西沿岸(气候和海洋环境变冷)等地区均有记录(Dyke et al, 1996;Dahl-Jensen et al, 1998; Kaplan et al, 2002; Kaufman et al, 2004; Seidenkrantz et al, 2007, 2008; Knudsen et al,2008; Andresen et al, 2013; Erbs-Hansen et al, 2013),但出现时间略有不同。如Kaplan等(2002)认为格陵兰南部新冰期开始于 3000a BP, Andresen等(2011)认为格陵兰西部新冰期开始于 3800a BP, 而格陵兰东南部则开始于3600a BP左右 (Andresen et al, 2013)。
新冰期时西格陵兰洋流中极地冷水团增强、北大西洋暖水团减弱的特征在 Disko湾南部的沉积物岩性、有孔虫及硅藻分析结果中也同样有所反映(Moros et al, 2006; Lloyd et al, 2007)。如 Moros等(2006)研究发现 Disko湾地区相对温暖的气候结束于 3500a BP左右, 3500—2000a BP期间Disko湾存在明显的变冷事件(Lloyd et al, 2007), 考古学证据也表明以水上猎取食物为生的 Saqqaq部落搬离Disko湾的原因很可能与当时气候变冷导致的食物缺乏及生存环境恶劣有关(Moros et al, 2006; Lloyd et al, 2007)。此外, 位于西格陵兰南部 Ameralik峡湾处的多项指标记录显示西格陵兰洋流中北大西洋暖水团的强度在 3200—2700a BP期间减弱(Seidenkrantz et al, 2007), 与主成分分析重建的极地冷水团的增强相对应。格陵兰东部和东南部的海洋沉积也同样记录了3500a BP以后新冰期气候变冷, 大西洋暖水团减弱, 北冰洋冷水团增强, 格陵兰冰盖扩张, 以及极锋南移(Jennings et al,2011; Andresen et al, 2013)。与此同时, Disko湾海冰覆盖面积波动增加(图5c), 表明新冰期时东格陵兰寒流携带的极地冷水团强度逐渐增强, 而印明格暖流强度减弱, 从而影响了研究区域的海冰覆盖变化(Jiang et al, 2002; Sha et al, 2014)。受此影响, 西格陵兰北部的巴芬湾 3600a BP以来海冰覆盖时间增加(Levac et al, 2001), 而有孔虫大西洋暖水指示种含量3000a BP左右不断减少(Knudsen et al, 2008), 加拿大极地群岛 3500—3000a BP期间海冰覆盖明显增强(Vare et al, 2009; Belt et al, 2010)。以上研究均表明,新冰期时西格陵兰洋流中极地冷水团的增强与海洋环境转冷、海冰覆盖增加和冰盖消融的减缓密切相关(Fisher et al, 1995)。
5.3 2000 a BP以来气候波动期
2000a BP以来DA06-139G孔样品在主成分一上的得分出现波动增加的趋势。其中, 1900—1500a BP期间, 主成分得分较低(图 5a), 海冰种和极地冷水种硅藻含量低于平均值(图3a, b), 而暖水种硅藻含量在平均值之上(图3c), 这表明该时期极地冷水团的强度减弱, 而大西洋暖水团对Disko湾的影响增强。有孔虫大西洋暖水指示种含量的显著增加(图 5b), Disko湾海冰覆盖度的明显减小(Sha et al, 2014; 图5c), 也表明该时期大西洋暖水团对Disko湾的影响增强, 使得海冰覆盖面积明显减少。
Disko湾南部DA00-02P孔和DA00-03P孔的硅藻、底栖有孔虫和颗石藻等含量变化同样显示在2200/2000—1500/1400a BP期间, 西格陵兰洋流携带的暖水团增强, 导致 Disko湾海水表层温度增加(Moros et al, 2006; Lloyd et al, 2007; Seidenkrantz et al, 2008)。此外, 东格陵兰沿岸的有孔虫记录(Jennings et al, 2002), 以及格陵兰南部Igaliku峡湾(Lassen et al, 2004)和Holsteinsborg海槽(Erbs-Hansen et al, 2013)的有孔虫记录也都显示 2000—1500a BP期间东格陵兰寒流强度较弱, 而西格陵兰洋流中印明格暖流势力增强, 海洋环境变暖。
1500—1200a BP期间, DA06-139G孔各样品在主成分一上的得分显著升高(图5a), 海冰种和极地冷水种硅藻含量高于平均值(图3a, b), 而指示北大西洋暖水团的暖水种硅藻(图3c)及有孔虫(图5b)含量显著下降, 且海冰密集度显著增加(图 5c), 表明该时期极地冷水团对Disko湾的影响增强, 从而导致海水表层温度降低, 与当时格陵兰冰芯 DYE-3记录的冷事件以及北大西洋海水表层温度的下降有很好的一致性(Dahl-Jensen et al, 1998; Jiang et al, 2002)。
Disko湾地区, 基于沟鞭藻-转换函数重建的全新世以来的海水表层温度变化也记录了1500a BP以来的降温事件(Ouellet-Bernier et al, 2014); 孢粉记录指示了 1450—1250a BP期间的冷事件(Ribeiro et al,2012); 颗石藻暖水组合含量减少反映了 1500—1300a BP期间北大西洋暖水团的影响减弱(Seidenkrantz et al, 2008); 而底栖有孔虫记录也反映1600—1200a BP期间较强的极地冷水团(Seidenkrantz et al, 2007)。此外, 格陵兰南部的硅藻和颗石藻记录同样表明该时期受东格陵兰寒流的影响较强, 海洋环境变冷(Jensen et al, 2004; Roncaglia, 2004)。
1200—650a BP期间, 除 1050—850a BP以外,DA06-139G孔各样品在主成分一上的得分下降(图5a), 有孔虫大西洋暖水指示种含量的增加(图5b), 海冰密集度也显著减少(图5c), 反映该时期西格陵兰洋流中极地冷水团强度减弱, 而北大西洋暖水团势力增强, Disko湾海洋环境转暖, 海冰覆盖减少。此外,Camp century冰芯 δ18O记录也显示 1200—800a BP期间, 格陵兰地区气温明显上升(Vinther et al, 2009,图5d), 对应于中世纪暖期。
Disko湾MSM343300孔1000—800a BP期间夏季海水表层温度比现在高约(4.4± 1.24)°C, 达到 10°C左右, 而该孔沟鞭藻重建的夏季海水表层温度也记录了这一暖期(Ouellet-Bernier et al, 2014)。Hass(1996)重建了欧洲北部海域晚全新世以来的古气候变化,认为中世纪暖期并不是一个持续的暖期, 而是被1050—800a BP这一冷期一分为二, 本文硅藻结果与前人研究结果一致 (图5a)。
有关中世纪暖期的记载, 在格陵兰西部、北大西洋北部和西北部海域等均有发现。其中, 在北大西洋西北部Holsteinsborg海槽处, 硅藻记录所反映的海洋环境变化显示在1200—650a BP期间, 该海域表层海水温度较高(Sha et al, 2012)。在格陵兰西南部Igaliku峡湾, Jensen等(2004)研究发现1180—600a BP期间东格陵兰寒流强度减弱, 使得该峡湾海冰覆盖面积减小, 而 Roncaglia等(2004)通过颗石藻记录研究发现990—665a BP期间季节性海冰覆盖明显减弱。
另外, 在东格陵兰沿岸, 1200—850a BP期间极地冷水团影响逐渐减弱, 使得大西洋中层水对东格陵兰沿岸的影响增强(Jennings et al, 1996)。在1200—650a BP期间, 冰岛北部沿岸也主要受大西洋暖水团的影响(Eiríksson et al, 2000; Jiang et al, 2002;Knudsen et al, 2004, 2012; Ran et al, 2011)。同样, 在加拿大极地群岛的硅藻记录也指示了1200—600a BP期间较为温暖的气候和海洋环境(LeBlanc et al,2004)。Kinnard等(2011)根据高分辨率的陆生指标重建了北冰洋夏季海冰覆盖, 指出750 a BP前北冰洋夏季海冰值极低, 与中世纪暖期相对应。
DA06-139G孔各样品在主成分一上的得分在650a BP之后显著升高(图 5a), 暖水种硅藻在 650 a BP之后含量显著下降甚至消失, 海冰种和极地冷水种硅藻大量出现(图3a, b), 表明该时期西格陵兰洋流中极地冷水团势力显著增强, 暖水团减弱, Disko湾地区气候变冷, 对应于小冰期。
Camp century冰芯δ18O记录反映500—100a BP期间西格陵兰地区气温下降(Vinther et al, 2009; 图5d)。同时, Disko湾海冰密集度的重建结果也显示,500a BP以来海冰显著增加(Sha et al, 2014, 图5c),与本文各样品在主成分一上的得分反映的极地冷水团显著增强具有很好的一致性, 说明小冰期时东格陵兰寒流携带的极地冷水团携带了大量的极地海冰随着西格陵兰海流北上, 间接影响Disko湾地区。
Disko湾的有孔虫记录也表明500a BP之后极地冷水团对Disko湾水文影响增强, 使得海洋环境变冷(Lloyd, 2006a)。已有研究表明, 小冰期期间格陵兰西部、格陵兰南部海域等也均存在较高的海冰覆盖率以及较寒冷的海洋环境(Jensen et al, 2004; Roncaglia et al, 2004; Sha et al, 2012)。
此外, 格陵兰东南部沉积硅藻记录反映750a BP之后东格陵兰寒流增强(Jensen, 2003), 这一现象在该区域其他沉积记录中也有被发现(Wagner et al,2000)。同时, 根据硅藻重建的冰岛北部海水表层温度显示, 650a BP之后东格陵兰寒流增强, 使得该海域海水表层温度显著降低(Jiang et al, 2002)。生物标志物IP25的研究结果也表明在750a BP之后冰岛北部海域海冰覆盖明显增强(Massé et al, 2008)。格陵兰冰芯重建的温度结果也显示在400a BP和100a BP, 温度分别比现在低0.5°C和0.7°C(Dahl-Jensen et al, 1998)。
6 结论
通过研究西格陵兰地区Disko湾DA06-139G孔沉积物中主要硅藻属种的百分含量变化及主成分分析发现, 过去5000年以来特征性硅藻属种(包括海冰种、极地冷水种和暖水种)百分含量的变化及主成分轴一可以指示研究区西格陵兰洋流中冷暖水团强度的变化。
5000—3800a BP期间, 海冰种和极地冷水种的含量较低, 暖水种含量较高, 样品在主成分一上的得分较低, 表明当时西格陵兰洋流中极地冷水团势力较弱, 而北大西洋暖水团则较强; 海冰种和极地冷水种含量在 3800—2000a BP开始增加, 样品在主成分一上的得分较高, 对应暖水种含量的减少, 表明该时期冷水团的势力增强, 而印明格暖流携带的北大西洋暖水团减弱; 2000a BP以后, 海冰种和极地冷水种含量呈波动上升的趋势, 但在 1900—1500a BP以及1200—600a BP期间存在两个低值, 样品在主成分一上的得分较高, 表明 2000a BP以来西格陵兰洋流中极地冷水团的势力不断增强, 期间间隔了北大西洋暖水团的两次增强事件。
Disko湾DA06-139G孔过去5000年以来特征性硅藻属种的百分含量变化及主成分分析结果反映的西格陵兰洋流中冷暖水团的强度变化与同一钻孔有孔虫北大西洋暖水指示种百分含量、海冰密集度的重建结果、以及Camp century冰芯δ18O记录的大气温度反映的西格陵兰地区气候变化有很好的一致性。
Aebly F A, Fritz S C, 2009. Palaeohydrology of Kangerlussuaq(Søndre Strømfjord), West Greenland During the Last—8000 Years. The Holocene, 19(1): 91—104
Andresen C S, Hansen M J, Seidenkrantz M S et al, 2013. Midto late-Holocene oceanographic variability on the Southeast Greenland shelf. The Holocene, 23(2): 167—178
Andresen C S, McCarthy D J, Dylmer C V et al, 2011.Interaction between subsurface ocean waters and calving of the Jakobshavn Isbræ during the late Holocene. The Holocene, 21(2): 211—224
Axford Y, Losee S, Briner J P et al, 2013. Holocene temperature history at the western Greenland Ice Sheet margin reconstructed from lake sediments. Quaternary Science Reviews, 59: 87—100
Bauch H A, Polyakova Y I, 2000. Late Holocene variations in Arctic shelf hydrology and sea-ice regime: evidence from north of the Lena Delta. International Journal of Earth Sciences, 89(3): 569—577
Belt S T, Vare L L, Massé G et al, 2010. Striking similarities in temporal changes to spring sea ice occurrence across the central Canadian Arctic Archipelago over the last 7000 years.Quaternary Science Reviews, 29(25—26): 3489—3504
Bindschadler R A, 1984. Jakobshavns Glacier drainage basin: a balance assessment. Journal of Geophysical Research:Oceans, 89(C2): 2066—2072
Briner J P, Kaufman D S, Bennike O et al, 2013. Amino acid ratios in reworked marine bivalve shells constrain Greenland Ice Sheet history during the Holocene. Geology, 42(1):75—78
Cremer H, 1999. Distribution patterns of diatom surface sediment assemblages in the Laptev Sea (Arctic Ocean). Marine Micropaleontology, 38(1): 39—67
Cuny J, Rhines P B, Niiler P P et al, 2002. Labrador Sea boundary currents and the fate of the Irminger Sea water.Journal of Physical Oceanography, 32(2): 627—647
Dahl-Jensen D, Mosegaard K, Gundestrup N et al, 1998. Past temperatures directly from the Greenland ice sheet. Science,282(5387): 268—271
Dalhoff F, Kuijpers A, 2007. Havbunds prøveindsamling ud for Vest Grønland 2006. RV Dana Cruise Report. Danmarks og Grønlands Geologiske Undersøgelse Rapport. 4: 1—51
De Sève M A, Dunbar M J, 1990. Structure and composition of ice algal assemblages from the Gulf of St. Lawrence,Magdalen Islands area. Canadian Journal of Fisheries and Aquatic Sciences, 47(4): 780—788
Dyke A S, Hooper J, Savelle J M, 1996. A history of sea ice in the Canadian Arctic Archipelago based on postglacial remains of the bowhead whale (Balaena mysticetus). Arctic, 49(3):235—255
Eiríksson J, Knudsen K L, Haflidason H et al, 2000. Late-glacial and Holocene palaeoceanography of the North Icelandic shelf. Journal of Quaternary Science, 15(1): 23—42
Erbs-Hansen D R, Knudsen K L, Olsen J et al, 2013.Paleoceanographical development off Sisimiut, west Greenland, during the mid- and late Holocene: a multiproxy study. Marine Micropaleontology, 102: 79—97
Fisher D A, Koerner R M, Reeh N, 1995. Holocene climatic records from Agassiz ice cap, Ellesmere Island, NWT,Canada. The Holocene, 5(1): 19—24
Håkansson B, 1990. On the stability properties of an oceanic jet stream. Journal of Physical Oceanography, 20(10):1638—1643
Hasle G R, Syvertsen E E, 1997. Marine diatoms. In: Tomas C R ed. Identifying Marine Phytoplankton. San Diego: Academic Press, 5—386
Hass H C, 1996. Northern Europe climate variations during late Holocene: evidence from marine Skagerrak.Palaeogeography, Palaeoclimatology, Palaeoecology,123(1—4): 121—145
Heimdal B R, 1989. Arctic Ocean phytoplankton. In: Herman Y ed. The Arctic Seas. New York: Van Nostrand Reinhold Company, 193—222
Hendey N I, 1964. An introductory account of the smaller algae of British coastal waters. Part V: bacillariophyceae (diatoms).Fishery Investigations Series IV of U.K. Ministry of Agriculture, Fisheries and Food. London: Her Majesty's Stationary Office
Jennings A E, Andrews J, Wilson L, 2011. Holocene environmental evolution of the SE Greenland Shelf North and South of the Denmark Strait: irminger and East Greenland current interactions. Quaternary Science Reviews,30(7—8): 980—998
Jennings A E, Knudsen K L, Hald M et al, 2002. A mid-Holocene shift in Arctic sea-ice variability on the East Greenland Shelf.The Holocene, 12(1): 49—58
Jennings A E, Weiner N J, 1996. Environmental change in eastern Greenland during the last 1300 years: evidence from foraminifera and lithofacies in Nansen Fjord, 68°N. The Holocene, 6(2): 179—191
Jensen K G, 2003. Holocene hydrographic changes in Greenland coastal waters: reconstructing environmental change from sub-fossil and contemporary diatoms. Copenhagen: Doctoral Dissertation of University of Copenhagen, 1—89
Jensen K G, Kuijpers A, Koç N et al, 2004. Diatom evidence of hydrographic changes and ice conditions in Igaliku Fjord,South Greenland, during the past 1500 years. The Holocene,14(2): 152—164
Jiang H, 1996. Diatoms from the surface sediments of the Skagerrak and the Kattegat and their relationship to the spatial changes of environmental variables. Journal of Biogeography, 23(2): 129—137
Jiang H, Björck S, Ran L H et al, 2006. Impact of the Kuroshio Current on the South China Sea based on a 115 000 year diatom record. Journal of Quaternary Science, 21(4):377—385
Jiang H, Eiríksson J, Schulz M et al, 2005. Evidence for solar forcing of sea-surface temperature on the North Icelandic Shelf during the late Holocene. Geology, 33(1): 73—76
Jiang H, Muscheler R, Björck S et al, 2015. Solar forcing of Holocene summer sea-surface temperatures in the northern North Atlantic. Geology, 43(3): 203—206
Jiang H, Seidenkrantz M S, Knudsen K L et al, 2001. Diatom surface sediment assemblages around Iceland and their relationships to oceanic environmental variables. Marine Micropaleontology, 41(1—2): 73—96
Jiang H, Seidenkrantz M S, Knudsen K L et al, 2002.Late-Holocene summer sea-surface temperatures based on a diatom record from the north Icelandic shelf. The Holocene,12(2): 137—147
Justwan A, Koç N, Jennings A E, 2008. Evolution of the Irminger and East Icelandic Current systems through the Holocene,revealed by diatom-based sea surface temperature reconstructions. Quaternary Science Reviews, 27(15—16):1571—1582
Kanaya T, Koizumi I, 1966. Interpretation of diatom thanatocoenoses from the North Pacific applied to a study of core V20-130 (Studies of a deep-sea core V20-130. Part IV).Science Reports of the Tohoku University, Series 2(Geology), 37(2): 89—130
Kaplan M R, Wolfe A P, Miller G H, 2002. Holocene environmental variability in southern Greenland inferred from lake sediments. Quaternary Research, 58(2): 149—159 Karentz D, Smayda T J, 1984. Temperature and seasonal occurrence patterns of 30 dominant phytoplankton species in Narragansett Bay over a 22-year period (1959-1980). Marine Ecology-Progress Series, 18: 277—293
Kaufman D S, Ager T A, Anderson N J et al, 2004. Holocene thermal maximum in the western Arctic (0-180°W).Quaternary Science Reviews, 23(5—6): 529—560
Kelly M, 1980. The status of the Neoglacial in western Greenland.Rapport Grønlands Geologiske Undersøgelse, 96: 1—24
Kinnard C, Zdanowicz C M, Fisher D A et al, 2011.Reconstructed changes in Arctic sea ice over the past 1, 450 years. Nature, 479(7374): 509—512
Knudsen K L, Eiríksson J, Bartels-Jónsdóttir H B, 2012.Oceanographic changes through the last millennium off North Iceland: temperature and salinity reconstructions based on foraminifera and stable isotopes. Marine Micropaleontology, 84—85: 54—73
Knudsen K L, Eiríksson J, Jansen E et al, 2004.Palaeoceanographic changes off North Iceland through the last 1200 years: foraminifera, stable isotopes, diatoms and ice rafted debris. Quaternary Science Reviews, 23(20—22):2231—2246
Knudsen K L, Stabell B, Seidenkrantz M S et al, 2008. Deglacial and Holocene conditions in northernmost Baffin Bay:sediments, foraminifera, diatoms and stable isotopes. Boreas,37(3): 346—376
Koç Karpuz N, Schrader H, 1990. Surface sediment diatom distribution and Holocene paleotemperature variations in the Greenland, Iceland and Norwegian Sea. Paleoceanography,5(4): 557—580
Koç Karpuz N, Jansen E, Haflidason H, 1993. Paleoceanographic reconstructions of surface ocean conditions in the Greenland,Iceland and Norwegian seas through the last 14 ka based on diatoms. Quaternary Science Reviews, 12(2): 115—140
Krawczyk D, Witkowski A, Moros M et al, 2010. Late-Holocene diatom-inferred reconstruction of temperature variations of the West Greenland Current from Disko Bugt, central West Greenland. The Holocene, 20(5): 659—666
Krawczyk D W, Witkowski A, Waniek J J et al, 2014. Description of diatoms from the Southwest to West Greenland coastal and open marine waters. Polar Biology, 37(11): 1589—1606 Lassen S J, Kuijpers A, Kunzendorf H et al, 2004. Late-Holocene Atlantic bottom-water variability in Igaliku Fjord, South Greenland, reconstructed from foraminifera faunas. The Holocene, 14(2): 165—171
LeBlanc M, Gajewski K, Hamilton P B, 2004. A diatom-based Holocene palaeoenvironmental record from a mid-arctic lake on Boothia Peninsula, Nunavut, Canada. The Holocene,14(3): 417—425
Levac E, De Vernal A, Blake Jr W, 2001. Sea-surface conditions in northernmost Baffin Bay during the Holocene:palynological evidence. Journal of Quaternary Science,16(4): 353—363
Lloyd J M, 2006a. Late Holocene environmental change in Disko Bugt, west Greenland: interaction between climate, ocean circulation and Jakobshavn Isbrae. Boreas, 35(1): 35—49
Lloyd J M, 2006b. Modern distribution of benthic foraminifera from disko bugt, west Greenland. The Journal of Foraminiferal Research, 36(4): 315—331
Lloyd J M, Kuijpers A, Long A et al, 2007. Foraminiferal reconstruction of mid- to late-Holocene ocean circulation and climate variability in Disko Bugt, West Greenland. The Holocene, 17(8): 1079—1091
Lloyd J M, Moros M, Perner K et al, 2011. A 100 yr record of ocean temperature control on the stability of Jakobshavn Isbrae, West Greenland. Geology, 39(9): 867—870
Lloyd J M, Park L A, Kuijpers A et al, 2005. Early Holocene palaeoceanography and deglacial chronology of Disko Bugt,West Greenland. Quaternary Science Reviews, 24(14—15):1741—1755
Long A J, Roberts D H, 2003. Late Weichselian deglacial history of Disko Bugt, West Greenland, and the dynamics of the Jakobshavns Isbrae ice stream. Boreas, 32(1): 208—226
Massé G, Rowland S J, Sicre M A et al, 2008. Abrupt climate changes for Iceland during the last millennium: evidence from high resolution sea ice reconstructions. Earth and Planetary Science Letters, 269(3—4): 565—569
Matthews J A, 2007. Neoglaciation in Europe. In: Elias S eds.Encyclopedia of Quaternary Science. Amsterdam: Elsevier,1122—1133
McNeely R, Dyke A S, Southon J R, 2006. Canadian marine reservoir ages, preliminary data assessment. Open File 5049.Canada: Geological Survey of Canada, Open File, 3
Medlin L K, Priddle J, 1990. Polar Marine Diatoms. Cambridge:British Antarctic Survey, 190
Møller H S, Jensen K G, Kuijpers A et al, 2006. Late-Holocene environment and climatic changes in Ameralik Fjord,southwest Greenland: evidence from the sedimentary record.The Holocene, 16(5): 685—695
Moros M, Jensen K G, Kuijpers A, 2006. Mid-to late-Holocene hydrological and climatic variability in Disko Bugt, central West Greenland. The Holocene, 16(3): 357—367
Ólafsdóttir S, Jennings A, Geirsdóttir Á et al, 2010. Holocene variability of the North Atlantic Irminger current on the south- and northwest shelf of Iceland. Marine Micropaleontology, 77(3—4): 101—118
Ouellet-Bernier M M, de Vernal A, Hillaire-Marcel C et al, 2014.Paleoceanographic changes in the Disko Bugt area, West Greenland, during the Holocene. The Holocene, 24(11):1573—1583
Overpeck J, Hughen K, Hardy D et al, 1997. Arctic environmental change of the last four centuries. Science,278(5341): 1251—1256
Polyakova Y I, 2001. Late Cenozoic evolution of northern Eurasian marginal seas based on the diatom record.Polarforschung, 69: 211—220
Ramsey C B, 2008. Deposition models for chronological records.Quaternary Science Reviews, 27(1—2): 42—60
Ran L H, Jiang H, Knudsen K L et al, 2011. Diatom-based reconstruction of palaeoceanographic changes on the North Icelandic shelf during the last millennium. Palaeogeography,Palaeoclimatology, Palaeoecology, 302(1—2): 109—119
Reimer P J, Baillie M G, Bard E et al, 2009. IntCal09 and Marine09 radiocarbon age calibration curves, 0-50 000 years cal BP. Radiocarbon, 51(4): 1111—1150
Ren J, Gersonde R, Esper O et al, 2014. Diatom distributions in northern North Pacific surface sediments and their relationship to modern environmental variables. Palaeogeography,Palaeoclimatology, Palaeoecology, 402: 81—103
Ren J, Jiang H, Seidenkrantz M S et al, 2009. A diatom-based reconstruction of Early Holocene hydrographic and climatic change in a southwest Greenland fjord. Marine Micropaleontology, 70(3—4): 166—176
Ribeiro S, Moros M, Ellegaard M et al, 2012. Climate variability in West Greenland during the past 1500 years: evidence from a high-resolution marine palynological record from Disko Bay. Boreas, 41(1): 68—83
Roncaglia L, 2004. New acritarch species from Holocene sediments in central West Greenland. Grana, 43(2): 81—88
Roncaglia L, Kuijpers A, 2004. Palynofacies analysis and organic-walled dinoflagellate cysts in late-Holocene sediments from Igaliku Fjord, South Greenland. The Holocene, 14(2): 172—184
Seidenkrantz M S, Aagaard-Sørensen S, Sulsbrück H et al, 2007.Hydrography and climate of the last 4400 years in a SW Greenland fjord: implications for Labrador Sea palaeoceanography. The Holocene, 17(3): 387—401
Seidenkrantz M S, Roncaglia L, Fischel A et al, 2008. Variable North Atlantic climate seesaw patterns documented by a late Holocene marine record from Disko Bugt, West Greenland.Marine Micropaleontology, 68(1—2): 66—83
Sha L B, Jiang H, Knudsen K L, 2012. Diatom evidence of climatic change in Holsteinsborg Dyb, west of Greenland,during the last 1200 years. The Holocene, 22(3): 347—358
Sha L B, Jiang H, Seidenkrantz M S et al, 2014. A diatom-based sea-ice reconstruction for the Vaigat Strait (Disko Bugt,West Greenland) over the last 5000 yr. Palaeogeography,Palaeoclimatology, Palaeoecology, 403: 66—79
Solomon S, Qin D, Manning M et al, 2007. Climate Change 2007:The Physical Science Basis: Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, United Kingdom,New York, NY, USA: Cambridge University Press, 996
Syvertsen E E, Hasle G R, 1984. Thalassiosira bulbosa Syvertsen,sp. nov., an arctic marine diatom. Polar Biology, 3(3):167—172
Tang C C L, Ross C K, Yao T et al, 2004. The circulation, water masses and sea-ice of Baffin Bay. Progress in Oceanography,63(4): 183—228
Vare L L, Massé G, Gregory T R et al, 2009. Sea ice variations in the central Canadian Arctic Archipelago during the Holocene.Quaternary Science Reviews, 28(13—14): 1354—1366
Vinther B M, Buchardt S L, Clausen H B et al, 2009. Holocene thinning of the Greenland ice sheet. Nature, 461(7262):385—388
von Quillfeldt C H, 1996. Ice algae and phytoplankton in north norwegian and arctic waters: species composition,succession and distribution. Norway: Doctoral Dissertation of University of Tromsø, 251
von Quillfeldt C H, 2000. Common diatom species in Arctic spring blooms: their distribution and abundance. Botanica Marina, 43(6): 499—516
Wagner B, Melles M, Hahne J et al, 2000. Holocene climate history of Geographical Society Ø, East Greenland-evidence from lake sediments. Palaeogeography, Palaeoclimatology,Palaeoecology, 160(1—2): 45—68
Williams K M, 1990. Late Quaternary paleoceanography of the western Baffin Bay region: evidence from fossil diatoms.Canadian Journal of Earth Sciences, 27(11): 1487—1494
Williams L, Bradley R, 1985. Paleoclimatology of the Baffin bay region. In: Andrew J T ed. Quaternary Environments:Eastern Canadian Arctic, Baffin Bay and Western Greenland.Boston: Allen and Unwin, 741—772
猜你喜欢
法医学杂志(2022年1期)2022-06-21
法医学杂志(2022年1期)2022-06-21
大气科学学报(2021年3期)2021-08-04
公民与法治(2020年2期)2020-05-30
热带海洋学报(2020年3期)2020-05-25
中国水运(2020年2期)2020-05-18
黑龙江水利科技(2020年8期)2020-01-12
环球时报(2020-01-03)2020-01-03
环球时报(2019-08-22)2019-08-22
环球时报(2016-06-28)2016-06-28
