
黄山松研究综述(1960-2014)及其知识图谱分析
2015-12-24 07:37苏松锦刘金福兰思仁李文周
福建农林大学学报(自然科学版) 2015年5期
苏松锦,刘金福,兰思仁,洪 伟,李文周
(1.福建农林大学,福建 福州350002;2.三明林业学校,福建 三明365001;3.福建农林大学海峡自然保护区研究中心,福建福州350002;4.戴云山国家级自然保护区管理局福建泉州362503)
黄山松(Pinus taiwanensis Hsia)为中国特有树种,是我国东部亚热带高海拔山地绿化、造林和生态恢复的重要树种.在中山气候恶劣环境下,黄山松具有马尾松无法企及的生态演替功能;在高山植被恢复、固碳、水源涵养、针叶药效开发以及应对全球气候变化等方面显示出巨大的生态、社会与经济价值.戴云山拥有6400 hm2原生性黄山松群落,是世界最大的黄山松种质基因基地,也是亚热带中山地区植被演替研究的理想地.为让国内外学者对黄山松最新研究动态有所了解,笔者对其分类与资源分布、遗传学、生态学、生理生化、森林培育和其它方面进行梳理.另一方面,在全球气候变暖、生物多样性监测以及森林生态系统定位监测研究日益深入背景下,对黄山松进行知识图谱分析可快速挖掘或预测其研究前沿,并追踪其发展动向,可为后续研究提供合作契机和参考依据.
1 黄山松研究进展
1.1 分类与资源分布
黄山松首个模式标本出自黄山西海门[1].不同学者对黄山松与台湾松分类争议较大,郑万钧等认为二者过渡性变异尚未达到质的飞跃,提议将其合并为黄山松,拉丁学名为Pinus taiwanensis Hsia,得到众多学者的支持[2];但从二者的外部形态、木材解剖超微结构、树脂道结构、细胞和遗传学特征等看,不少学者认为黄山松是独立种系[3,4].在马尾松和黄山松基因渐渗区存在杉松、武陵松等形态特征介于二者之间的变种.不同学者对黄山松与其它松树的分类关系可用图1表示.
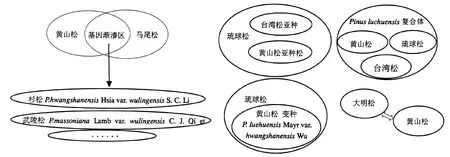
图1 黄山松与其它松树分类关系Fig.1 Relationship between Pinus-taiwanensis and other pine tree species
黄山松在豫、皖、浙、闽、台、赣、湘、鄂、晋、桂、黔、滇等省均有分布[2].从水平分布看,可将其分为台湾、南部、西南部、东南沿海、中西部、东北和北部大别山7个气候生态区[5]和闽南区、闽赣区、湘西南区、皖浙区、鄂湘赣和豫皖大别山区6个地理类型[6];从垂直分布看,黄山松在长江流域以南和以北海拔分布的上下限分别约为 800-1800 m 和 500-1700 m[5].
1.2 遗传学
中国大陆天然黄山松不同居群间遗传一致度变化范围为0.5-0.8,遗传距离与地理距离呈显著“S”型关系[6].从核型看,黄山松有4-5对具二次缢痕染色体,而台湾松仅第2对染色体具二次缢痕[7];从遗传多样性看,黄山松核苷酸多样性与种内遗传分化显著高于马尾松;二者在C3H座位上存在共享单倍型,种间差异GI大于C3H;GI基因座进化速度相对较快[8].在确定黄山松等松属植物DNA多态性方面,采用整合微流控芯片优于凝胶电泳方法[9].从谱系生物地理学看,黄山松、琉球松和台湾松种间、种内均存在大量花粉流;mtDNA主要遗传变异发生在3种松树居群间,而cpDNA遗传变异则主要集中在群体内部;黄山松群体历史扩张事件可能发生时间早于最近一次第四纪盛冰川期;3种松树在第四纪冰期可能存在多个避难所[4].黄山松与马尾松在海拔垂直分布上存在自然杂交和基因渐渗区,在基因渐渗过程中,马尾松基因向杂交群体渐渗频率高于黄山松[10].为揭示黄山松与马尾松的生殖隔离遗传机制,李淑娴[11]系统地开展了AFLP实验程序优化及引物组合筛选;遗传图谱构建及标记偏分离和偏分离遗传位点定位及相关基因组区域确定研究.
1.3 生态学
1.3.1 空间格局 植物空间格局研究有助于解决营造林中植株配置和采伐问题,是物种共存、森林更新与演替等研究的基础[12].植物空间格局与尺度、计量参数和研究方法等有关.同一物种在不同地区空间分布差异较大,如武夷山黄山松种群在高海拔呈聚集分布,而在低海拔呈随机分布[13];福建寿宁县黄山松种群呈聚集分布,聚集强度大树>幼树>中径树,并随取样尺度增加而减少[14];黄山地区黄山松种群幼年期呈聚集分布,随着年龄增长逐渐向随机分布过渡[15];而福建屏南县黄山松种群则呈随机分布[14].对于空间格局潜在生态学过程的解释,多数学者认为黄山松总体上呈聚集分布,并随年龄或海拔增加趋于随机分布,与其自身特性、竞争及所处的土壤、地形和气候等环境条件有关[15].负二项参数、扩散型指标Iδ、分形关联维数和计盒维数等传统计量参数与研究方法未考虑样点位置信息,且只能分析单一尺度空间格局;而O-ring统计弥补了传统指标的不足,且克服了Ripley's K函数随尺度变化出现累积效应的问题[16],是当前植被空间格局研究的主流方法.空间格局不仅因物种而异,而且同一物种在不同发育阶段、不同生境、不同林层也有明显区别[12].随着生物多样性监测网络不断拓展与研究的日益深入,采用O-ring统计在武夷山、戴云山、黄山和天柱山等地开展1-50 hm2大样地尺度黄山松种群空间格局是未来值得关注的方向之一.
1.3.2 生态位 生态位是生态学研究核心内容之一.戴云山黄山松8个群丛中91.78%种对生态位重叠值为0-0.5;与黄山松更新生态位重叠值较大的树种有窄基红褐柃(Eurya rubiginosa)和短尾越桔(Vaccinium carlesii)[17].武夷山黄山松林多数种群在坡度、坡向和海拔上生态适应能力相似性,生态位普遍重叠,资源共享趋势明显[18].古田山黄山松、木荷(Schima superba)、野漆树(Rhus sylvestris)和甜槠(Castanopsis eyrei)等生态位宽度和重叠度均较大[19].种群生态位宽的物种,其生态位重叠不一定高,如闽西北黄山松生态位宽度较大(0.5- 0.6),但其与杉木(Cunninghamia lanceolata)生态位重叠值仅 0.3424[20].
1.3.3 种群结构与动态 不同林型和同一林型不同密度下断面积最大增长速度径级范围差异显著[21].逻辑斯谛或广义逻辑斯谛模型在拟合黄山松种群数量动态规律时,效果良好[22].从结构与数量特征看,不同地区黄山松种群差异明显:黄山黄山松种群死亡高峰期为幼苗1级向2级的过渡期,其幼苗在海拔较高的山顶、山脊和陡坡等生境下自然更新较好[15].天柱山黄山松种群随海拔升高,林分密度增大,径级分布范围减小;种群存活曲线由Deevey-Ⅱ型向Deevey-Ⅰ型过渡,群落由稳定型向增长型过渡,与物种竞争引起的自疏和它疏作用有关[23].而福建龙栖山和屏南县的黄山松种群存活曲线整体上在DeeveyⅡ-Ⅲ型之间[22].
1.3.4 生态系统定位监测 森林生态系统定位监测主要包括水文、土壤、气象、生物等内容.林盛松[24]观测了闽东北鹫峰山天然黄山松林的降雨分配格局;董金相[25]估测了戴云山黄山松林植被层、0-60 cm土壤层和枯枝落叶层的碳储量,并分析其地形分异规律.韩勇等[26]研究了武夷山黄山松凋落叶在不同植物群落中的分解动态;吴泽民等[27]分析了黄山景区黄山松年轮S及Pb、Mn、Zn和Cu等元素的动态特征.林益明和林鹏等[28,29]则系统地报道了武夷山黄山松各器官灰分含量、干重热值、去灰分热值以及N、P、K、Na累积与循环规律.朱德煌[17]采用除趋势典范对应分析法分析了环境因子对黄山松群落格局的影响.郑仕平[30]发现黄山松林密度为3200株·hm-2的生产力和养分利用率最大.黄山松作为中国东部亚热带中山地区典型代表性群系,在全球变暖背景下,开展黄山松集水区与嵌套流域、蒸散量、根际微生态、小气候、碳通量、痕量气体与气溶胶、粗木质残体、稳定同位素和氮沉降等方面定位监测研究,有助于揭示其生态系统结构和功能的时空变化规律和对环境的响应与适应机制[31].
1.3.5 群落特征与演替规律 不同地区黄山松群落特征差异大.天柱山黄山松群落物种组成共有52科105属,具有北亚热带常绿阔叶林和暖温带落叶阔叶林特征[32];古田山黄山松群落生活型以高位芽植物为主,群落叶以小型叶、单叶、草质和非全缘为主[33];湖南平江幕阜山黄山松群落种子植物约40科66属70种[34].从物种多样性看,天柱山黄山松群落物种Shannon-Wiener指数按灌木层、草本层和乔木层降序排列[32];古田山黄山松群落物种多样性灌木层>乔木层>草本层[33].黄山松群落多样性与海拔、中度旅游干扰等有关[36,37].黄山松耐逆向人为干扰能力弱,且正向人为干扰下恢复进程缓慢[37].从演替规律看,黄山生境优越地区,黄山松纯林朝针、阔混交林演变,并最终被落叶阔叶林群落取代;而在裸岩、陡坡、峰岭或梁岗等生境中,黄山松可形成稳定地形顶级群落[15,38].在平江幕阜山,黄 山松群落演替规律为裸地→黄山松林→黄山松+ 落 叶阔叶树混交林→落 叶阔叶林→常绿阔叶林[34].从干扰角度看,按5.3%/10 a林冠干扰强度算,西天目山黄山松阔叶林在2051-2071年可能完全被阔叶纯林取代[39].从更新角度看,吴泽民等[40]发现黄山松林隙形成与更新发生时间相近,40 m2林隙或110 m2扩展林隙的光照能保证黄山松幼苗的正常生长.
1.3.6 竞争 安徽天柱山黄山松种内竞争强度随对象木胸径增大而降低,采用逐步扩大范围法可有效地确定其邻体竞争范围[41].为精确预测林分生长,在构建林分竞争生长模型时需综合考虑林木胸径、冠幅和林龄等.黄山松种群自疏效应明显,利用枝条着枝深度和林木变量建立的树冠形状模型及密度效应模型,可测算其生产弹性值和边际产量[13].竞争格局与种群自身、环境等有关,对不同阶段黄山松幼树进行适当透光伐,可提高其与阔叶树的竞争能力[42].黄山松种内竞争在寿宁县小于屏南县;在闽西北,天然黄山松个体空间关联较强、竞争激烈和空间占据程度高的海拔范围为1300-1310 m和1400 m[43].林分密度控制对林分经营具有科学指导意义,考虑竞争因素,福建寿宁县黄山松合理栽植密度应控制在4500株·hm-2[44].
1.4 生理生化
黄山松适应高山严酷环境能力强,其生理生态稳定性主要通过改变表型来实现.不同海拔黄山松针叶长、单根叶重和比叶面积差异显著[45].黄山松中生型、边生型、内生型和鞘内型针叶树脂道与海拔关系密切,针叶树脂道数目主要受≥10℃积温、日照时数和降水量制约,年轮结构和轴向树脂道纬向变异明显[6].从光合特性看,黄山松不同年龄段1年生针叶时蒸腾强度、水分利用率日变化均呈双峰分布;黄山松幼龄受气孔限制明显,中幼龄抗旱能力最强[46].黄山松光合速率夏季>秋季>春季>冬季,春、夏、秋3季有光合午休现象;其光合速率主要受细胞间隙CO2浓度和气孔导度等影响[47].由于光补偿点和光饱和点均较高,黄山松幼苗在林下竞争中处于劣势,更新较困难.从生化角度看,低温处理下黄山松幼苗浸出液相对电导率、超氧化物歧化酶活性及丙二醛质量摩尔浓度均随海拔升高而降低[48].针叶叶绿素和蛋白质含量可反映黄山松的衰老状况[49].
1.5 森林培育
1.5.1 生长过程分析 生长过程是植物定向培育研究的理论基础.Logistic模型可较好地模拟黄山松生长过程.黄山松天然林胸径生长量比人工林低0.47 cm·a-1;树高连年生长量最高峰(第20年)和材积快速生长点(第40年)比人工林分别迟5年和20年[50].黄山松林分形高随林分树高增大而增大,而随胸高断面积或疏密度增大而减少[51].从生物量看,密度为1530株·hm-2的10年生黄山松人工林生产力约1.675 t/(hm2·a)[52].疏伐可显著提高球果生物量,当黄山松种子园郁闭度为0.5时,球果产量最高[53].采用经验公式W=a+bD+cD2预测黄山松生物量可达到较高精度,但速生阶段宜用W=a+bD[54].以立地质量为哑变量、间隔期为自变量的黄山松林分收获模型,可有效解决天然异龄林生长动态预测中林龄不易确定问题,并为择伐收益法在资产评估中的推广应用提供技术支撑[55].
1.5.2 种子园建设 黄山松优树选择可采用数量与形质指标,个体树与对比树相结合方法[56];高海拔黄山松采种最佳时间段为霜降前后,采种时间越早,种子品质越差[57].黄山松较低的种子发芽率(21.6%)与综合逆境活力(15.5%)一定程度上影响了其繁殖能力[15];在阴天对高约12 cm黄山松苗进行切根,可促进其径粗生长与根系发达,并显著提高其菌根感染率与造林成活率[58].黄山松无性系分株主要依靠嫁接来完成,嫁接时应优先考虑嫁接部位和接穗粗度.浙江天台县华顶林场黄山松种子园无性系物侯期和生长节律、土壤改良、树体管理、定砧与嫁接、花粉管理及病虫害防治等方面营建技术先进,效果良好,值得示范推广[59].
1.5.3 抚育与造林成效 抚育与造林成效分析是黄山松林集约化经营的重要环节.黄山松幼林生长主要受坡位、种源和造林密度影响,而整地方式、施肥和幼林抚育对其影响较小[60].黄山松与光皮桦2:1混交,林分生长最好,并可有效解决杉木采伐迹地更新困难问题[61].编制黄山松数量化地位指数表可扩展地位质量评定适用范围[62].高海拔山地杉木、黄山松人工混交林对幼龄杉木个体树高和中龄杉木个体胸径有促进作用,其立木蓄积、胸高断面积和生物量明显高于杉木纯林[63].
1.5.4 病虫害防治 对黄山松有威胁的病虫害主要有:松材线虫(Bursaphelenchus xylophilus)、松毛虫(Dendrulimus marmortus)、小卷蛾(Petrova cristata)、松果梢斑螟(Dioryctria pryeri)、松针褐斑病(Lecanosticta acicola)、细纹新须螨(Cenopalpus lineola)、小环绵盘菌(Cyclaneusmam inus)、黄山松黑松叶蜂(Nesodiprion.SP)、松斑螟(Salebria laruata)、松栎锈病、赤枯病、紫色病(缺镁)等[38,64].其中学者最为关注:黄山松是否为松材线虫感病树种[65-67]?黄山松感染松材线虫可能与供试苗、接种源以及接种气候条件等有关,自然状态下受松材线虫侵染的黄山松罹病木呈现出针叶部分变红、木材有松香味、有松褐天牛危害痕迹等特点;黄山松感病木仅分布在其海拔下限区和人工栽植的林分中,而较高海拔天然黄山松林中未见有感染松材线虫报道[65].筛选病虫害抗性候补木是种子园建设的重要工作.陈凤毛等[66]发现拟松材线虫CFS4,SLD3,ZJ1和KOR1四个株系对黄山松苗有致病作用,其中接种ZJ1或SLD3,黄山松苗死亡率均为16.7%.徐六一等[67]于2005-2007年间筛选了57个家系、216株黄山松抗性候补木,随后对筛选出的抗性候选个体开展MuPS标记识别分析.
1.6 其它
树轮气候学:树木年轮分析对了解影响植物生长关键因子和重建历史气候资料意义重大.不同地区黄山松直径生长及其影响因子差异较大,黄山黄山松径向生长主要受当年>10℃积温的太阳辐射和积温影响[68];大别山黄山松径向生长主要受当年2-7月平均气温限制[69];九龙山自然保护区和安徽省南部牯牛降的高海拔黄山松树轮径向生长主要受夏季(上年6-7月和当年6月)水热条件影响[70];而西大别山黄山松树轮宽度生长主要受9、10月的水热组合影响[71].
针叶的开发与利用:黄山松松针功能性成分研究目前仅见于马文秀[72]鉴定出8,15-异海松二烯酸、8,15-海松二烯酸和湿地松酸;刘晓庚等[73]报道了黄山松的粗蛋白质、多聚戊糖、纤维素、灰分、叶绿素和胡萝卜素等理化成分含量.黄山松松针在保健食品和药业生产上的开发与应用有待深入.
灾害分析:黄山松受冰雪灾害,易发生树干弯曲、断干、掘根、断冠和冻死等现象,树高径粗易断大枝和断冠;树矮径细、冠幅偏大易断干或被掘根;幼树易被冻死;尖削度小、树冠较高且不均匀的黄山松抵御冰雪灾害能力较弱[74].
2 黄山松文献知识图谱分析
以“黄山松”为主题词,Web of Science和中国知网CNKI为数据库,对1960-2014年与林学相关学科领域文献进行检索,共得到461条记录;去重、辨别(删除书评及会议等)和整理后,获得文献220篇.采用CitespaceⅢ软件[75]对文献进行格式转换,并对其关键词、作者、机构和期刊进行分析.
2.1 时间分布
黄山松受到关注和重视始于20世纪90年代,1995年以后,其发文数量呈波动上升.图2中发文数量较多的年份为2000年(14篇)、2006年(12篇)和2012年(13篇).220篇文献中仅有6篇被SCI收录,表明黄山松研究国际社会影响力较弱.

图2 1960-2014年黄山松林学相关文献年篇数Fig.2 Papers of forestry literature on Pinus taiwanensis per year from 1960 to 2014

图3 1960-2014年黄山松研究关键词的知识图谱Fig.3 Mapping knowledge domain of keywords in P.taiwanensis research from 1960 to 2014
2.2 关键词分析
将CitespaceⅢ软件中时区设为3年,以关键词为网络节点,基于各时区中高被引前10个关键词,生成关键词共被引聚类知识图谱(图3).关键词知识图谱网络中共有118个节点,107个连接,网络密度为0.0155.网络中节点大小表示文献数量,连线表示节点间合作关系,连线粗细表示合作的频次,中心性表示节点在网络中的重要性.从图3可知,黄山松知识图谱网络中重要性大于0.08的关键词为黄山松、马尾松、树脂道、天然更新和遗传多样性;频次大于6的关键词为黄山松、马尾松、松属、林分、生物量、树脂道、天然更新和种群(表1).
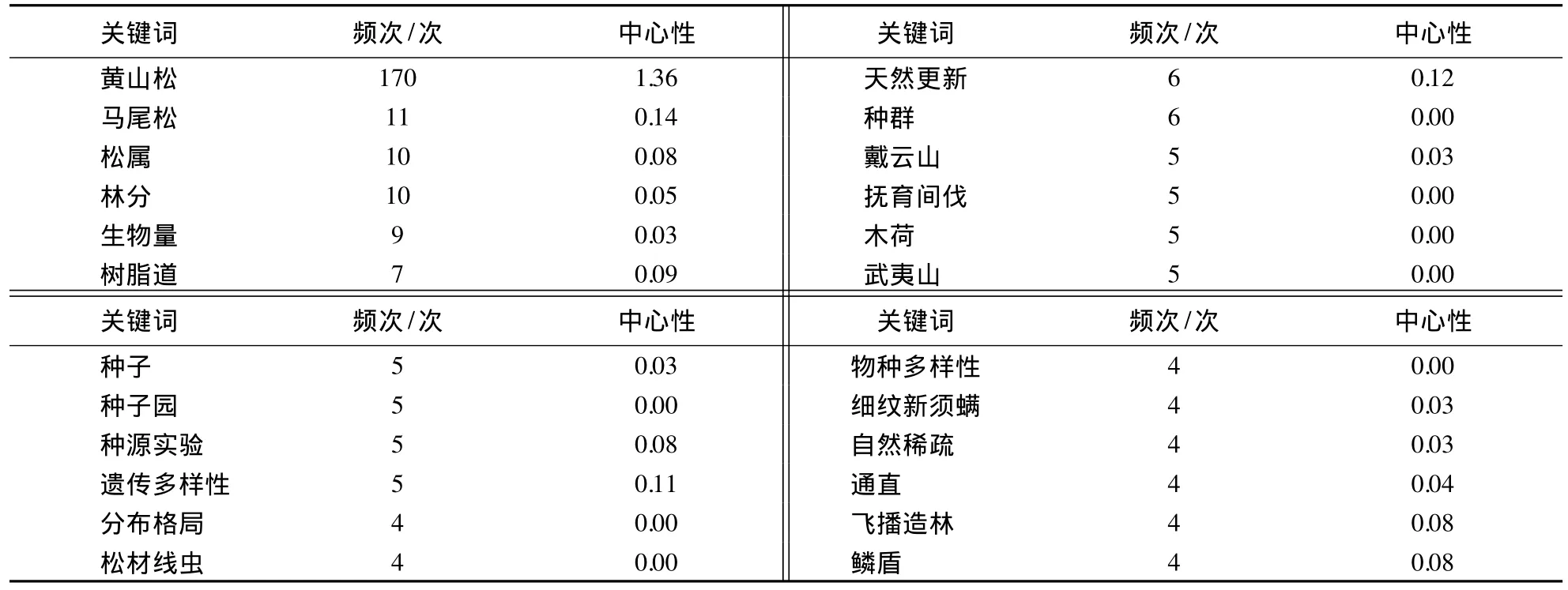
表1 1960-2014年黄山松研究前24个高频关键词Table 1 Top 24 keywords of P.taiwanensis research appearing frequently from 1960 to 2014
2.3 发文作者分析
作者共被引聚类知识图谱网络中共有327个节点,251个连接,网络密度为0.0047.节点越大,表明作者发文频数越高.洪伟在图谱中出现频次最高,主要从事黄山松种群生态学研究;其次为范义荣、吴承祯、吴泽民和罗世家等(图4),分别从事林木遗传与良种选育、种群生态学、森林生态学和植物学研究.发文较多作者之间合作关系密切,如吴承祯与洪伟,其它作者以独自研究状态为主.老师及其指导的学生是研究团体的核心部分.如福建农林大学洪伟与吴承祯、封磊、郑世群、刘金福和宋萍等;浙江农林大学范义荣与其学生,安徽农业大学吴泽民与其学生.发文数量在4篇以上作者共28人(表2).

图4 1960-2014年黄山松研究作者的知识图谱Fig.4 Mapping knowledge domain of authors in P.taiwanensis research(1960-2014)
2.4 发文期刊分析
参照《北京大学中文核心期刊目录(2011年版)》,黄山松研究前19位期刊118篇文献有78篇为农业科学类核心期刊.220篇文献中SCI期刊、核心期刊、博硕士论文分别有6、121和14篇,占总数的64.09%,表明发文质量相对较高(表3).

表2 1960-2014年黄山松研究前28个作者Table 2 Top 28 authors in P.taiwanensis research from 1960 to 2014
黄山松发文机构共被引聚类知识图谱网络中共有145个节点,48个连接,网络密度为0.0046.发文频次最高3家机构分别为福建农林大学、安徽农业大学和浙江农林大学(图5).福建农林大学与戴云山国家级自然保护区;黄山学院与黄山风景区园林局;南京林业大学和安徽省林业勘察设计院、湖北民族学院等之间合作关系较紧密,其它机构研究较独立,如安徽农业大学、浙江农林大学、福建省寿宁县林业局等.

表3 1960-2014年黄山松研究前19位期刊Table 3 Top 19 journals in Pinus taiwanensis research from 1960 to 2014

图5 1960-2014年黄山松研究发文机构的知识图谱Fig.5 Mapping knowledge domain of institutes in P.taiwanensis research from 1960 to 2014

表4 1960-2014年黄山松研究前17个机构Table 4 Top 17 institutes of P.taiwanensis research from 1960 to 2014
3 结论与展望
黄山松遗传学和生理生化研究结果至今仍未能较好地解决黄山松分类问题,但多数学者倾向于将黄山松作为独立种系,最常用学名为P.taiwanensis Hayata.目前生物多样性、生态系统定位观测、植被维持机制及其与环境因子耦合关系等研究,多基于1-50 hm2大样地定位观测数据.戴云山国家级自然保护区6400 hm2黄山松群落是中国东南沿海针叶林生态系统的典型代表,其科研重点应放在针叶林大样地建设、空间格局和生态系统定位监测上.黄山松人工林培育过程中应重视良种选育以及病虫害防治.黄山松径向生长及其对气候变化响应结果区域差异大.因此,戴云山黄山松树轮宽度气候学分析是一个极有意义的研究点.黄山松在全国分布范围较广,但其景观生态学研究几乎空白,在景观、区域或更大尺度上开展其碳储量(生物量)、叶面积指数LAI、植被净生产力NPP的反演研究,可丰富陆地生态系统和生物多样性与全球变化的相互作用机制研究.从关键词、作者和发文机构知识图谱看,黄山松林业领域研究以独立研究状态为主,合作研究较少,建议今后应加强南北方黄山松林景观或区域等大尺度合作研究.
[1]夏纬英.黄山松[J].中国植物学杂志,1936(1):17.
[2]中国科学院中国植物志编辑委员会.中国植物志(第七卷)[M].北京:科学出版社,1978.
[3]邹惠渝,黎志,罗世家,等.黄山松的分类学研究[J].植物研究,2003,23(3):278-279.
[4]余雷.黄山松的谱系生物地理学研究[D].合肥:安徽农业大学,2012.
[5]童再康,范义荣.黄山松分布区气候生态区划[J].中南林学院学报,1993,13(1):81-87.
[6]石雷.中国大陆地区黄山松地理变异的形态和RAPD分析[D].合肥:安徽农业大学,2012.
[7]邢有华,方永鑫,欧阳家安.黄山松的核型研究[J].安徽农学院学报,1985(2):72-76.
[8]张利锐,彭艳玲,任广朋,等.马尾松和黄山松两个核基因位点的群体遗传多样性和种间分化[J].植物生态学报,2011,35(5):531-538
[9]唐娟娟,吴江林,刘金华,等.用整合微流控芯片系统分析黄山松RAPD片段多态性[J].遗传,2003,25(4):419-424.
[10]罗世家,邹惠渝,梁师文.黄山松与马尾松基因渐渗的研究[J].林业科学,2001,37(6):118-122.
[11]李淑娴.马尾松和黄山松物种分化遗传机制研究[D].南京:南京林业大学,2012.
[12]HOU JH,Mi XC,LIU CR,et al.Spatial patterns and associations in a Quercus-Betula forest in northern China[J].Journal of Vegetation Science,2004,15:407-414.
[13]蔡小英.武夷山黄山松种群结构与动态研究[D].福州:福建农林大学,2008.
[14]宋萍,洪伟,吴承祯,等.天然黄山松种群空间格局的分形特征——关联维数[J].武汉植物学研究,2005,23(1):32-36.
[15]罗世家.黄山松的研究[D].南京:南京林业大学,1998.
[16]Wiegend T,Moloney K A.Rings,circles and null models for point pattern analysis in ecology[J].Oikos,2004,104:209-229.
[17]朱德煌.戴云山黄山松种群特征及其环境因子研究[D].福州:福建农林大学,2013.
[18]封磊,洪伟,吴承祯,等.武夷山黄山松林主要种群生态位特征[J].山地学报,2008,26(3):308-316.
[19]胡正华,于明坚,彭传正,等.古田山国家自然保护区黄山松林主要种群生态位研究[J].生态环境,2004,13(4):619-621,629.
[20]林思祖,黄世国,洪伟,等.杉阔混交林主要种群多维生态位特征[J].生态学报,2002,22(6):962-968.
[21]吴承桢,洪伟,林成来.黄山松种群数量动态研究[J].浙江林学院学报,1998,15(3):274-279.
[22]毕晓丽,洪伟,吴承祯.黄山松种群统计分析[J].林业科学,2002,38(1):61-67.
[23]段仁燕,黄敏毅,吴甘霖.黄山松种群结构与动态研究[J].生物学杂志,2009,26(1):35-37,34.
[24]林盛松.闽东北鹫峰山天然黄山松林内降雨分配的研究[J].福建林业勘察设计,2007(2):31-34.
[25]董金相.戴云山黄山松林碳储量及其影响因子研究[D].福州:福建农林大学,2012.
[26]韩勇,徐宪根,阮宏华,等.武夷山黄山松凋落叶在不同植物群落中的分解动态[J].南京林业大学学报:自然科学版,2010,34(3):141-145.
[27]吴泽民,高健,黄成林,等.黄山松年轮硫及重金属元素含量动态特征[J].应用生态学报,2005,16(5):820-824.
[28]林益明,林鹏.福建武夷山2个典型植物群落建群种的热值研究[J].武夷科学,1999,15:118-123.
[29]林益明,李振基,杨志伟,等.福建武夷山黄山松群落的氮、磷累积和循环[J].热带亚热带植物学报,1997,5(2):26-32.
[30]郑仕平.黄山松人工林养分循环与密度关系的研究[J].防护林科技,2014(8):13-16.
[31]丁访军.森林生态系统定位研究标准体系构建[D].中国林业科学研究院,2011.
[32]周葆华,余世金.天柱山黄山松群落特征及其环境功能评价[J].地理研究,2008,27(2):257-265.
[33]胡正华,于明坚,丁炳扬,等.古田山国家级自然保护区黄山松群落特征及物种多样性研究[J].生态环境,2003,12(4):436-439.
[34]李家湘,赵丽娟,黄展鹏.平江幕阜山黄山松群落特征及其演替规律的探讨[J].湖南林业科技,2004,31(5):16-18.
[35]任国学,刘金福,徐道炜,等.戴云山国家级自然保护区黄山松群落类型与物种多样性分析[J].植物资源与环境学报,2011,20(3):82-88.
[36]吴甘霖,黄敏毅,段仁燕,等.不同强度旅游干扰对黄山松群落物种多样性的影响[J].生态学报,2006,26(12):3924-3930.
[37]姚剑飞,方乐金,吴俊,等.干扰对黄山高海拔木本植被生长的影响[J].东北林业大学学报,2012,40(3):28-31.
[38]季春峰,邹惠渝,向其柏.黄山松研究进展(综述)[J].安徽农业大学学报,2004,31(1):111-114.
[39]夏冰,兰涛,贺善安,等.西天目山黄山松阔叶林的冠层干扰与动态推测[J].植物资源与环境,1995,4(3):15-20.
[40]吴泽民,黄成林,韦朝领.黄山松群落林隙光能效应与黄山松的更新[J].应用生态学报,2000,11(1):13-18.
[41]段仁燕,黄敏毅,吴甘霖,等.黄山松种群邻体范围与邻体竞争强度的研究[J].广西植物,2009,29(1):111-115.
[42]龚道怀.次生林内黄山松幼树的生长及其量化表达[J].林业勘察设计(福建),2009(1):102-105.
[43]毕晓丽,洪伟,吴承祯,等.黄山松林不同树种树冠分形特征研究[J].福建林学院学报,2001,21(4):347-350.
[44]张进德.黄山松造林密度与生物量的关系[J].林业科技通讯,2000(1):7-10.
[45]马元灿,王中生,余华,等.不同海拔梯度下黄山松与马尾松针叶形态·光合生理特性的研究[J].安徽农业科学,2012,40(29):14155-14158,14173.
[46]武金翠,吴泽民,洪淑媛,等.不同年龄黄山松蒸腾速率及水分利用效率的比较研究[J].安徽农业大学学报,2007,34(2):239-243.
[47]洪淑媛,吴泽民,刘西军.黄山松生理生态特性研究[J].安徽农业大学学报,2006,33(4):462-467.
[48]李淑娴,谭艳,陈颖,等.黄山松不同生理生化指标随海拔高度变化趋势[J].东北林业大学学报,2010,38(6):9-10,34.
[49]李东林,严景华,曹恒生,等.黄山松不同龄阶针叶衰老指标的比较研究[J].林业科学研究,1998,11(2):218-221.
[50]卢琦,赵体顺,罗天祥,等.黄山松天然林与人工林物种多样性和林分生长规律的比较研究[J].林业科学研究,1996,9(3):273-277.
[51]刘钦.黄山松人工林形高与断面积和疏密度的关系[J].林业科技,2003,17(增刊):19-20.
[52]胡道连,李志辉,谢旭东.黄山松人工林生物产量及生产力的研究[J].中南林学院学报,1998,18(1):60-64.
[53]陈见阳.黄山松种子园球果产量与郁闭度的研究[J].浙江林业科技,2014,34(1):58-60.
[54]郑德华.黄山松人工林生物量测定方法的比较[J].福建林业科技,1999,26(增刊):36-39.
[55]华伟平,池上评,李晓伟,等.黄山松天然林收获模型及在择伐收益法中的应用[J].福建林学院学报,2014,34(3):220-224.
[56]刘通高,张进德,范志平.黄山松天然林优树选择标准和方法的研究[J].福建林业科技,1999,26(增刊):54-57.
[57]王国英,占晓云,范义荣.不同采收期对黄山松种子品质的影响[J].浙江林业科技,1998,18(1):43-45.
[58]林盛松.黄山松大田苗切根培育技术研究[J].福建林业勘察设计,2006(2):122-125.
[59]范义荣,毛迎春,方陆明,等.黄山松种子园营建技术及初步效果[J].浙江林学院学报,1997,14(2):111-119.
[60]陈兰仙.造林技术措施对黄山松幼林生长的效应[J].福建林学院学报,2000,20(4):357-360.
[61]黄绍平,陶义俊,王兴平,等.庆元林场黄山松与光皮桦人工林混交营造比例研究[J].现代农业科技,2011(11):217,223.
[62]章礼拐.皖西黄山松人工林地位质量数量化评定的研究[J].安徽林业科技,1995(2):20-22.
[63]徐小牛,李宏开.杉木、黄山松人工混交林的生长特性[J].林业科学研究,1996,9(3):278-283.
[64]方乐金,李丰伯,万志兵,等.黄山松细纹新须螨发生与气象因素关系的分析[J].西南林业大学学报,2014,34(2):104-107.
[65]蒋丽雅,江顺利,汪振宇,等.黄山松自然状态下感染松材线虫病的初步调查[J].安徽农业大学学报,2006,33(1):5-8.
[66]陈凤毛,史延梅,王姝颖,等.不同株系拟松材线虫对黄山松和黑松苗木的致病性[J].林业科学,2011,46(12):86-90.
[67]徐六一,高景斌,蔡卫兵,等.黄山松松材线虫病抗性候补木的选拔及评价研究[J].安徽农业科学,2008,36(32):14093-14094,14115.
[68]吴泽民,黄成林,马青山.黄山松年轮生长和气候的关系[J].应用生态学报,1999,10(2):147-150.
[69]郑永宏,张永,邵雪梅,等.大别山地区黄山松和油松树轮宽度的气候意义[J].地理科学进展,2012,31(1):72-77.
[70]李玲玲,史江峰,侯鑫源,等.中国东南高海拔黄山松生长对气候的响应——以浙江省九龙山和安徽省枯牛降为例[J].应用生态学报,2014,25(7):1849-1856.
[71]彭剑峰,李国栋,李玲玲.西大别山小林海黄山松树轮宽度的气候意义[J].应用生态学报,2014,25(7):1857-1862.
[72]马文秀.黄山松松脂树脂酸少量成份研究[J].南京林业大学学报,1989,13(4):69-73.
[73]刘晓庚,陈梅梅.我国松针的开发利用研究及进展[J].林产化工通讯,2003,37(4):29-33.
[74]张志祥,刘鹏,邱志军,等.浙江九龙山自然保护区黄山松种群冰雪灾害干扰及其受灾影响因子分析[J].植物生态学学报,2010,34(2):223-232.
[75]CHEN C.CiteSpace II:Detecting and visualizing emerging trends and transient patterns in scientific literature[J].Journal of the American Society for Information Science and Technology,2006,57(3):359-377.?
猜你喜欢
今日农业(2022年15期)2022-09-20
体育科技文献通报(2022年3期)2022-05-23
辽金历史与考古(2021年0期)2021-07-29
科技传播(2019年22期)2020-01-14
民用飞机设计与研究(2019年4期)2019-05-21
红土地(2018年7期)2018-09-26
黄山学院学报(2017年5期)2017-11-03
山西教育·幼教(2017年11期)2017-04-13
武夷学院学报(2015年3期)2015-07-18
当代畜禽养殖业(2014年10期)2014-02-27
