
不同品种头花蓼叶片的比较解剖学研究
2015-12-22 02:39:10高凯悦廖海民刘世会赵德刚贵州大学生命科学学院农业生物工程研究院山地植物资源保护与种质创新省部共建教育部重点实验室贵州贵阳550025贵州省农业生物工程重点实验室贵州贵阳550025贵州大学药学院贵州贵阳550025
安徽农业科学 2015年16期
高凯悦,廖海民,刘世会,赵德刚,2* (.贵州大学生命科学学院/农业生物工程研究院,山地植物资源保护与种质创新省部共建教育部重点实验室,贵州贵阳 550025;2.贵州省农业生物工程重点实验室,贵州贵阳 550025;3.贵州大学药学院,贵州贵阳 550025)
头花蓼(Polygonum capitatum Ham ex D.Don)为蓼科(Polygonaceae)多年生匍匐草本植物,又称四季红、石莽草、太阳草等,主要分布于贵州、西藏、四川、广西、云南等省(区)。头花蓼全草入药,具有清热解毒、利尿通淋与活血止痛等功效[1],是贵州传统苗药。随着头花蓼在医药产业的不断发展,其市场需求也在急速上升。近年来,一些学者对头花蓼组织培养、人工栽培技术进行了大量研究[2-3],为头花蓼产业可持续发展奠定良好基础。但是,有关头花蓼解剖结构的研究尚未见报道,仅赛明兰等[4]对头花蓼药材进行过显微鉴定。笔者对野生头花蓼、GAP基地种植头花蓼、贵州省农业生物工程重点实验室选育的四倍体及其二倍体头花蓼品种叶片进行了解剖研究,以期为头花蓼优良品种的选育提供解剖学参考依据。
1 材料与方法
1.1 材料 供试材料4个,来源见表1。表1中的I、II、III品种由贵州大学熊源新教授鉴定,IV品种由贵州大学农业生物工程重点实验室诱导培育经流式细胞仪鉴定。

表1 供试头花蓼材料
1.2 方法
1.2.1 表皮撕片法。取相同部位叶片中部,采用指甲油印迹法撕取上下表皮[5]。表皮毛长度、表皮细胞和气孔大小均取50个的平均值;表皮细胞、气孔和表皮毛的密度取10个视野的平均值。
1.2.2 石蜡切片法。将新鲜叶片经浓度70%FAA固定,乙醇脱水,二甲苯透明,石蜡包埋。切片厚度10 μm,番红—固绿染色,加拿大树胶封片[6]。叶片、主脉、表皮细胞、栅栏组织及海绵组织的厚度均为30个的平均值,并且计算组织紧密度(CTR)、疏松度(SR)以及栅/海比。计算公式如下:
CTR=栅栏组织厚度/叶片厚度×100%
SR=海绵组织厚度/叶片厚度×100%
栅/海比=栅栏组织厚度/海绵组织厚度×100%
1.3 数据处理 用EXCEL和SPSS19.0软件对试验数据进行处理、统计分析。
2 结果与分析
2.1 叶表皮特征 头花蓼叶片表皮由表皮细胞、气孔器和表皮毛组成(图1)。
2.1.1 表皮细胞。头花蓼叶表皮细胞均排列紧密,其中上表皮细胞垂周壁近平直,细胞近方形,细胞较大,密度小;下表皮细胞垂周壁呈波状弯曲长条形,细胞较小,密度较大。由表2可知,在头花蓼4个材料中,相对于其他3个品种,多倍体细胞较大,密度更小,且差异在0.05水平显著。

表2 叶表皮细胞特征
2.1.2 气孔器。头花蓼气孔器均呈为无规则型,由2个肾形保卫细胞和气孔组成,上表皮气孔数明显少于下表皮。由表3可知,多倍体上表皮气孔明显大于其他3个品种,上、下表皮的气孔密度均较低。

表3 叶表皮气孔特征
2.1.3 表皮毛。头花蓼的表皮毛仅分布于叶脉上,为单细胞表皮毛类型。由表4可知,上、下表皮的表皮毛长度均为多倍体较长,且与其他3个品种差异在0.05水平显著;上、下表皮的表皮毛密度无显著性差异。

表4 叶表皮毛特征
2.2 叶片解剖结构 由图2可知,头花蓼叶片为典型的异 面叶结构,即横切面由表皮、叶肉和叶脉3个部分组成,叶肉包括栅栏组织和海绵组织2个部分。
头花蓼上、下表皮均为一层细胞,细胞呈长方形,上表皮细胞较大且有明显的角质层。由表5可知,多倍体的叶片厚度较大,与其他3个品种差异在0.05水平显著;上、下表皮的厚度均为多倍体较大,但差异不明显。
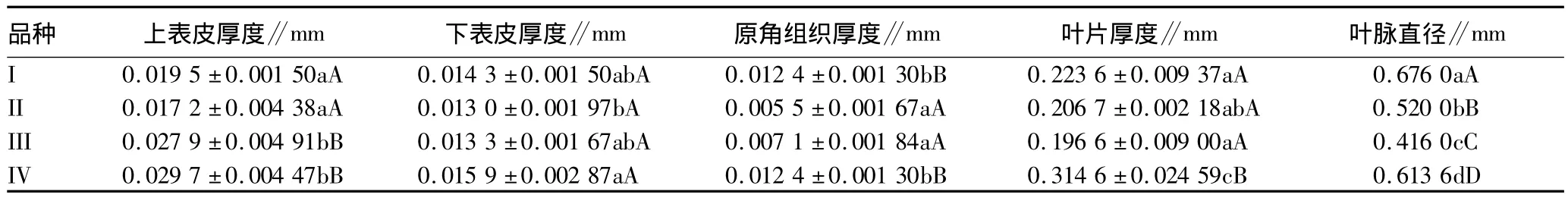
表5 叶片主脉和表皮结构特征
叶肉二型,分化为栅栏组织和海绵组织,在一些叶肉细胞中能看到草酸钙簇晶。靠近上表皮的部位为栅栏组织,组织两列,细胞呈长柱形,其组成细胞排列规则,细胞长轴垂直于叶表面;近下表皮部位的海绵组织,细胞近圆形,排列疏松,间隙较多。由表6可知,多倍体的栅栏组织和海绵组织厚度较大,与其他3个品种差异在0.05水平显著;组织紧密度、栅/海比较高。
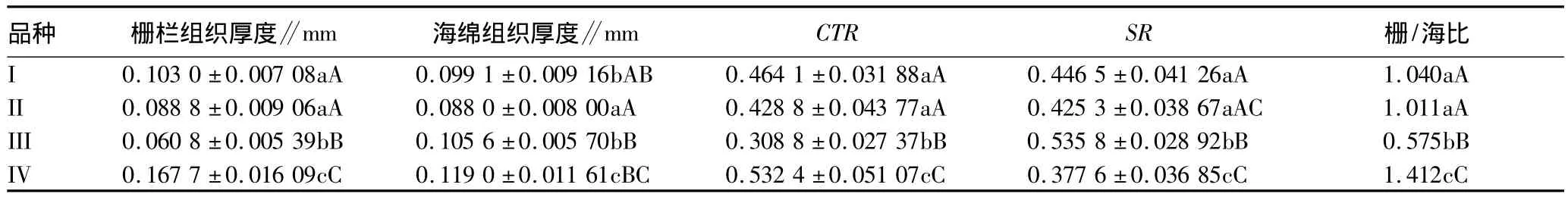
表6 叶肉的解剖结构特征
在头花蓼叶片中分布有大量的维管组织,构成各级叶脉,属典型网状脉序。叶脉维管束为外韧维管束类型,由木质部、形成层和韧皮部组成。木质部靠近叶的近轴面,韧皮部靠近叶的远轴面。头花蓼主脉靠近上下表皮处且有较多厚角组织分布,其细胞排列较紧密。主脉具2个维管束,上下排列。近轴面维管束较小,其韧皮部朝下,木质部朝上;远轴面维管束相对较大,木质部和韧皮部的排列与近轴面维管束相反,即其木质部朝下,而韧皮部朝上。
3 结论与讨论
3.1 头花蓼叶片的解剖结构 头花蓼叶片为典型的异面叶结构。横切面由表皮、叶肉和叶脉3个部分组成。叶肉包括栅栏组织和海绵组织2个部分。表皮由表皮细胞、气孔器和表皮毛组成。表皮细胞均排列紧密,上表皮细胞垂周壁近平直,细胞近方形,下表皮细胞垂周壁呈波状弯曲长条形。气孔器为无规则型,由呈肾形的2个保卫细胞和气孔组成,上表皮气孔数明显少于下表皮。表皮毛仅分布于叶脉上,下表皮相对较多。叶肉包括栅栏组织和海绵组织2个部分,靠近上表皮部位的为栅栏组织,其组成细胞排列规则,细胞呈长柱形,细胞长轴垂直于叶表面;近下表皮部位的为海绵组织,细胞近圆形,排列疏松,间隙较多。叶脉为外韧维管束类型,由木质部、形成层和韧皮部组成,木质部靠近叶的近轴面,韧皮部靠近叶的远轴面。
头花蓼主脉较特殊,具两个维管束,上下排列,近轴面维管束较小,其韧皮部朝下,木质部朝上;远轴面维管束相对较大,木质部和韧皮部的排列与近轴面维管束相反,即其木质部朝下,而韧皮部朝上。主脉的这种维管束排列较少见,在同属植物何首乌(Polygonum multiflorum Thunb.)有报道[7]。
3.2 不同品种头花蓼叶片的结构差异 在头花蓼4个材料中,多倍体与所研究的其他3个材料间差异较大。其表皮细胞较大,细胞密度小;上表皮气孔较大,上、下表皮的气孔密度均较低;上、下表皮的表皮毛长度较长;叶片、上表皮和下表皮的厚度均较大;栅栏组织和海绵组织的厚度较大;其组织紧密度、栅/海比较高。这些形态解剖结构特征表明,多倍体品种具有一定的优越性,对环境的适应能力更强。一般认为,气孔密度越小,角质层越厚,叶肉的栅栏组织越发达,植物抗寒能力就越强[8-10]。多倍体品种气孔密度最小,叶片最厚,CTR最大,SR最小。在抗寒性方面,与普通二倍体相比,多倍体具有明显的优势。
[1]贵州植物志编委会.贵州植物志(第一卷)[M].贵阳:贵州人民出版社,1982:197.
[2]闫吉美,刘世会,龙凤荣,等.头花蓼组培快繁体系的建立[J].山地农业生物学报,2012,31(6):477 -480.
[3]孙长生,梁斌,李孟林,等.头花蓼规范化种植技术[J].中国现代中药,2007,9(1):36 -39.
[4]赛明兰,张赛群.四季红形态与组织鉴定[J].贵阳医学院学报,1995,20(3):201 -203.
[5]晏升禄,崔丽,徐爽,等.商陆与垂序商陆营养器官的解剖学研究[J].山地农业生物学报,2012,31(2):124 -128.
[6]李祖任,胡楠,杨吉刚,等.繁缕和鹅肠菜的花维管束系统比较解剖学研究及其系统学意义[J].植物科学学报,2013,31(6):525 -532.
[7]王舒颖,徐爽,廖海民.何首乌幼苗的解剖学及过渡区的初生维管系统[J].山地农业生物学报,2011,30(1):20 -26.
[8]胡正海.植物解剖学[M].北京:高等教育出版社,2010.
[9]李芳兰,包维楷.植物叶片形态解剖结构对环境变化的响应与适应[J].植物学通报,2005,22(S1):118 -127.
[10]谢庭味,欧阳美珊.13种木兰科树种叶片解剖与其抗寒性[J].武汉植物学研究,1989,7(3):234 -238.
猜你喜欢
放射学实践(2023年9期)2023-09-20 08:24:16
放射学实践(2021年9期)2021-09-22 05:42:30
中成药(2018年3期)2018-05-07 13:34:42
首都食品与医药(2017年5期)2017-11-03 07:40:52
放射学实践(2017年5期)2017-06-01 12:19:24
科学启蒙(2016年8期)2016-09-27 11:42:41
天津农业科学(2015年11期)2015-12-03 00:03:49
江苏农业科学(2015年4期)2015-06-15 21:11:35
快乐作文·高年级(2014年3期)2014-06-06 13:02:54
热带作物学报(2014年3期)2014-04-29 18:17:49
